房山紫堇葉綠體基因組特征及其系統(tǒng)進化關系
2022-08-31 03:22:38李澳旋杜曉蓉喬永剛
草地學報 2022年8期
關鍵詞:植物
孫 哲, 李澳旋, 杜曉蓉, 宋 蕓, 喬永剛*
(1. 山西農(nóng)業(yè)大學生命科學學院, 山西 太谷 030801; 2. 中獸醫(yī)藥現(xiàn)代化山西省重點實驗室, 山西 太谷 030801)
紫堇屬是罌粟科最大的一個屬,在全球約有440種,為草本植物,無乳汁[1]。房山紫堇(CorydalisfangshanensisW. T. Wang ex S. Y. He)又稱石黃連、土黃連,是罌粟科紫堇屬的一種多生長于海拔500~1 600 m左右石灰?guī)r山坡的植物[2],開白色小花,生命力頑強,可防風固沙、保持水土[3];其根可入藥,具有清熱解毒的功效[4]。因獨特的形態(tài),特殊的生長環(huán)境和稀少的數(shù)量,房山紫堇被列為北京市重點保護野生植物。開展房山紫堇的保護和科學研究,可以使其在園林綠化、水土保持等方面發(fā)揮重要作用,也有利于豐富草地生態(tài)系統(tǒng)。
葉綠體是植物光合作用的場所[5],隨著一些生物技術的開發(fā)和應用,葉綠體的基因組大小得到確定,被子植物的葉綠體基因組大小為120~170 kb不等,通常編碼110~130個基因[6]。高等植物的葉綠體基因組結構高度保守,可以確保在進化群體之間基因的直接同源性,包括反向重復區(qū)(Inverted repeats,IRs)和大單拷貝區(qū)(Large single copy,LSC)、小單拷貝區(qū)(Small single copy,SSC),植物葉綠體基因組的大小差異由IRa和IRb決定[7]。葉綠體基因組所包含的遺傳信息沒有核基因組多,但其結構穩(wěn)定并且保守[8],可以為高等植物群體遺傳學、系統(tǒng)發(fā)育和基因組進化提供依據(jù)[9-10]。
紫堇屬植物在形態(tài)上很難區(qū)分,被認為是罌粟科中最難分類的一個屬[11],葉綠體基因組可以為紫堇屬植物的系統(tǒng)發(fā)育提供參考,國內(nèi)外已經(jīng)有紫堇(CorydalisedulisMaxim.)、陜西紫堇(CorydalisshensianaLiden)[12]、毛黃堇(CorydalistomentellaFranch.)、石生黃堇(CorydalissaxicolaBunting)[13]、斑花黃堇(CorydalisconspersaMaxim.)[14]等紫堇屬植物葉綠體基因組的相關報道。本研究對房山紫堇葉綠體基因組進行測序,對其GC含量、SSR位點等進行分析,并與紫堇屬其他物種的基因組進行多重比較,以期為房山紫堇的系統(tǒng)發(fā)育和紫堇屬植物的親緣關系提供數(shù)據(jù)支撐,同時豐富紫堇屬植物的葉綠體基因組數(shù)據(jù)庫。
1 材料與方法
1.1 試驗材料
房山紫堇種子采集自山西省陵川縣丈河村(北緯35°33′~35°38′,東經(jīng)113°08′~113°17′),在實驗室育苗后,移栽到藥用植物資源圃。于2020年10月采集健康無病害的房山紫堇植株的新鮮葉片,用酒精棉擦拭后,快速放入液氮,保存于—80℃冰箱備用。
1.2 試驗方法
1.2.1DNA提取和測序 用CTAB法提取保存的房山紫堇葉片DNA[15],DNA質(zhì)量和濃度檢測合格后,將DNA片段化(超聲波打斷),經(jīng)過片段純化、末端修復等途徑獲得測序文庫,質(zhì)檢合格后在陜西博瑞德生物科技有限公司用Illumina HiSeq進行測序。
1.2.2基因的組裝和注釋 用NOVOPlasty軟件[16]對測序所得的房山紫堇葉綠體基因組進行組裝,參考序列為Coreanomeconhylomeconoides(NC_031446.1)[17],獲得的葉綠體基因組用CPGAVAS軟件[18]進行基因注釋,結果提交至GenBank,序列號為MZ772033,用OGDRAW在線軟件[19]將結果可視化,GC含量用MEGA7[20]軟件進行分析。
1.2.3簡單重復序列(Simple sequence repeats,SSR)位點分析 利用MISA軟件[21]對房山紫堇的SSR位點進行分析,設置重復次數(shù):單核苷酸>10,雙核苷酸>6,三核苷酸>5,四核苷酸>5,五核苷酸>5,六核苷酸>5;2個SSR之間的最小距離為100 bp。
1.2.4密碼子組成及偏好性 運用軟件CodonW1.4.2[22]分析其葉綠體基因組的有效密碼子數(shù)(Effective number of codon,Nc)、相對同義密碼子使用情況(Relative synonymous codon usage,RSCU)、密碼子適應指數(shù)(Codon adaption index,CAI)、密碼子偏好性指數(shù)(Codon bias index,CBI)、最優(yōu)密碼子使用頻率(Frequency of optimal codons,FOP)等密碼子組成參數(shù),RSCU>1的密碼子具有偏好性[22]。
1.2.5多重比較和IR收縮與擴張分析 利用mVISTA軟件[23]將房山紫堇與石生黃堇、紫堇、毛黃堇葉綠體基因組進行多重比較,使用IRscope軟件[24]分析房山紫堇與石生黃堇、紫堇、毛黃堇的IR收縮與擴張情況。
1.2.6系統(tǒng)發(fā)育分析 以房山紫堇基因組序列和已經(jīng)公布的9個紫堇屬植物基因組序列為研究對象,選取苦參(SophoraflavescensAlt.)和苦豆子(SophoraalopecuroidesL.)為外群[25],利用MAFFT軟件[26]對12個物種的葉綠體全基因組進行比對,并使用MEGA7軟件構建ML系統(tǒng)發(fā)育樹,自展值Bootstrap value設為1 000,使用Kimura 2-parameter模型。
2 結果與分析
2.1 葉綠體基因組特征
基于邊合成邊測序技術,使用Illumina HiSeq高通量測序平臺對房山紫堇葉綠體基因組進行測序,共得到10.45 GB原始數(shù)據(jù),各樣品Q30堿基百分比均不小于90%。房山紫堇的葉綠體基因組為191 128 bp,GC含量為40.2%;呈現(xiàn)典型的四分體結構(圖1),大單拷貝區(qū)(LSC)為97 302 bp、小單拷貝區(qū)(SSC)為9 836 bp,反向重復區(qū)(IRa,IRb)為83 990 bp,GC含量分別為39.1%,35.2%,42.1%。

圖1 房山紫堇葉綠體基因組示意圖Fig.1 Chloroplast genome map of Corydalis fangshanensis
房山紫堇葉綠體基因組共注釋到127個基因(表1),包括84個蛋白質(zhì)編碼基因、35個tRNA基因以及8個rRNA基因,占比分別為66.1%,27.6%,6.3%。根據(jù)不同的功能可將這些基因分為4類,包括atpA,atpB,atpE,atpF,atpH等44個與光合作用有關的基因;rpl14,rpl16等73個與自我復制相關的基因;還有infA等其他基因以及一些功能未知的基因。其中17個基因具有內(nèi)含子,rps12,trnA-UGC,trnE-UUC,ndhB基因的內(nèi)含子多于1。

表1 房山紫堇葉綠體基因功能分類Table 1 Chloroplast gene functional classification of Corydalis fangshanensis
2.2 SSR位點分析
對房山紫堇單基因序列進行分析,結果顯示(表2),共檢測到40個SSR位點,其中單核苷酸有31個,雙核苷酸有8個,三核苷酸有1個。單核苷酸最多,包括7個A(占比17.5%),1個C(占比2.5%),23個T(占比57.5%);雙核苷酸總共8個,包括4個AT(占比10%)和4個TA(占比10%);三核苷酸只有1個CAA(占比2.5%)。
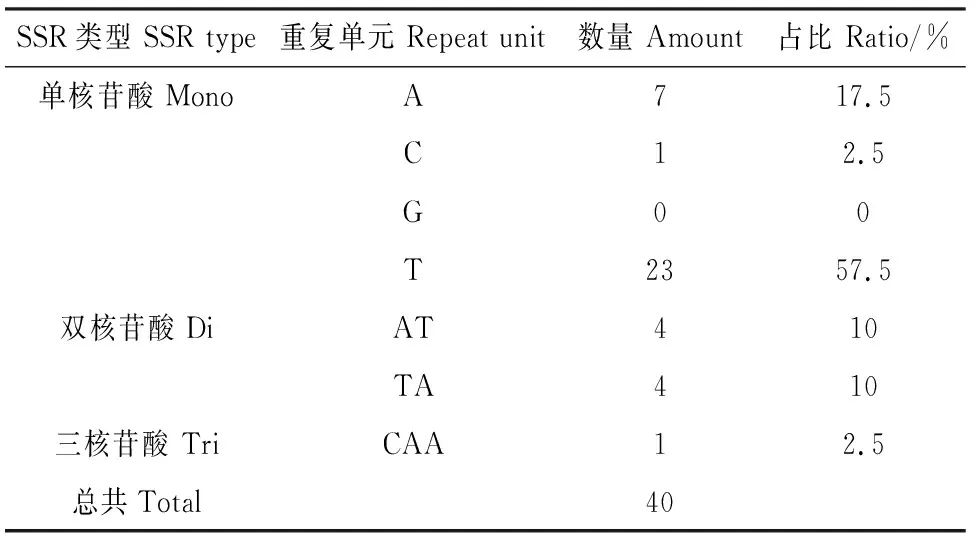
表2 房山紫堇SSR位點分析Table 2 SSR loci analysis of Corydalis fangshanensis
2.3 密碼子偏好性分析
對房山紫堇葉綠體基因組的密碼子組成分析結果顯示(表3):房山紫堇基因組的有效密碼子數(shù)為56.57,說明其密碼子偏好性較弱;GC含量為41.2%,GC3s含量為38.5%,均小于50%,說明密碼子偏向使用堿基A和堿基T。

表3 房山紫堇葉綠體基因組密碼子使用的總體特征Table 3 Overall characterization of codon usage in the chloroplast genome of Corydalis fangshanensis
對房山紫堇葉綠體基因組的相對同義密碼子進行分析(圖2),RSCU>1的密碼子有35個,其中以A/T堿基結尾的有28個,占80%,表明這些密碼子偏向以A/T結尾。

圖2 房山紫堇葉綠體基因組的相對同義密碼子使用情況Fig.2 Relative synonymous codon usage in chloroplast genomes of Corydalis fangshanensis
2.4 比較基因組學和IR收縮與擴張分析
以測序所得的房山紫堇基因組序列為參照,將其與石生黃堇、紫堇、毛黃堇進行相似性比較分析,結果如圖3所示,4種紫堇屬植物的編碼區(qū)較為保守,在ycf2,trnN-GUU,ndhF等基因所在的位置存在明顯的變異。邊界分析結果如圖4所示,房山紫堇rpl2基因位于LSC區(qū);ndhA基因位于IRb與SSC的交界處,在IRb區(qū)267 bp,SSC區(qū)域1 902 bp;ycf1位于SSC區(qū)域,距離SSC與IRa的交界407 bp;rpl23位于IR區(qū)域,距離IRa與LSC的交界35 bp,石生黃堇與房山紫堇的邊界收縮情況最為相似。

圖3 房山紫堇與石生黃堇、紫堇、毛黃堇的多重比較結果Fig.3 The results of multiple comparisons between Corydalis fangshanensis and Corydalis saxicola,Corydalis edulis and Corydalis tomentella

圖4 房山紫堇與石生黃堇、紫堇、毛黃堇的邊界比較Fig.4 Boundary comparison of Corydalis fangshanensis and Corydalis saxicola,Corydalis edulis and Corydalis tomentella
2.5 系統(tǒng)發(fā)育分析
為了鑒定房山紫堇在紫堇屬植物中的進化位置,使用MEGA7構建ML系統(tǒng)進化樹,結果顯示(圖5),紫堇屬植物可以分為兩大類,房山紫堇與同屬植物紫堇、毛黃堇、石生黃堇聚為一大類,與紫堇[10]的親緣關系最近;灰綠黃堇(CorydalisaduncaMaxim.)[27]與其余5種紫堇屬植物聚為另一大類;外源植物苦參與苦豆子聚為一類,與前人的研究一致[16]。

圖5 基于葉綠體基因組的紫堇屬植物系統(tǒng)發(fā)育分析Fig.5 Phylogenetic analysis of Corydalis based on chloroplast genome
3 討論
紫堇屬植物物種豐富,種類繁多,僅根據(jù)形態(tài)很難對其分類。2015年山西首次報道發(fā)現(xiàn)了房山紫堇,房山紫堇全株可入藥,抗逆性強,對環(huán)境要求較低,可在干旱缺水的石灰?guī)r環(huán)境中生存,具有防風固沙、水土保持的生態(tài)價值[28]。房山紫堇被列入《世界自然保護聯(lián)盟紅色名錄》(IUCN)中,中國珍稀瀕危植物信息系統(tǒng)將其列為易危(VU)植物,禁止直接采挖[29]。
目前有部分紫堇屬植物的葉綠體基因組數(shù)據(jù)得到公布[30],大部分紫堇屬植物的葉綠體基因組有待揭示。葉綠體基因、線粒體基因和細胞核基因是系統(tǒng)發(fā)育和物種起源研究中常用的三種數(shù)據(jù)來源[31]。高度保守的葉綠體基因組[8]含有豐富的遺傳信息,研究房山紫堇的葉綠體基因組對了解房山紫堇的屬內(nèi)進化位置、遺傳變異、系統(tǒng)發(fā)育以及開發(fā)利用等具有重要的價值。
SSR位點分析對研究物種鑒定、遺傳變異、系統(tǒng)進化有重要意義,對房山紫堇進行SSR位點分析,發(fā)現(xiàn)其有40個SSR位點,包括單核苷酸、雙核苷酸和三核苷酸,房山紫堇葉綠體基因組SSR多含有AT堿基,與前人研究結果一致[32]。Nc,RSCU,GC,GC3s均可以反映密碼子的偏好性,Nc值通常在20~61之間,靠近61偏好性弱,靠近20偏好性強;RSCU>1時,該密碼子為使用頻率較高的密碼子,RSCU=1時,密碼子沒有偏好性,RSCU<1時,其為低使用頻率的密碼子;GC和GC3s含量大于50%時,說明該物種葉綠體基因組偏向使用GC堿基,反之,偏向使用AT堿基。對房山紫堇的密碼子分析結果顯示:Nc值靠近61,說明房山紫堇葉綠體基因組密碼子使用性偏弱;RSCU>1的密碼子超過半數(shù),其中以A/T堿基結尾的占80%,并且GC含量和GC3s含量均小于50%,房山紫堇偏向使用AT堿基,與段春燕[33]、宋蕓[34]等人的研究結果一致。植物葉綠體基因組的收縮和擴張是其變異的主要機制,本研究對房山紫堇基因組與石生黃堇、紫堇、毛黃堇進行比較分析,結果顯示石生黃堇與房山紫堇的邊界收縮情況高度相似。聚類結果顯示:紫堇屬植物序列被聚為一類,兩種豆科植物作為外源種被聚為一類[25],與預期結果一致,房山紫堇與紫堇的親緣關系最近,在對房山紫堇進行開發(fā)利用時,可將紫堇作為參考物種。本研究豐富了紫堇屬植物的葉綠體基因組數(shù)據(jù),為探索紫堇屬植物的起源及親緣關系提供了依據(jù),為房山紫堇的進一步科學研究奠定了基礎。
4 結論
本研究用Illumina HiSeq對房山紫堇進行測序,將測序結果進行整理得出以下結論:房山紫堇的葉綠體基因組大小為191 128 bp,GC含量為40.2%,呈現(xiàn)典型的四段式結構,注釋到127個基因;有40個SSR位點;葉綠體基因組的密碼子偏好性較弱,偏向使用A和T這兩種堿基;編碼區(qū)較為保守;房山紫堇與紫堇的親緣關系最近。目前對房山紫堇的報道很少,本研究為房山紫堇的物種保護、系統(tǒng)進化、資源的開發(fā)利用提供了相關試驗數(shù)據(jù)和理論依據(jù)。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
