二肽基肽酶-4通過CXCR4/mTOR信號通路介導小鼠肺泡巨噬細胞MH-S自噬的機制研究
2022-08-31 07:11:44楊帆鄧璐陳睦虎
中國現代醫學雜志 2022年16期
關鍵詞:小鼠
楊帆,鄧璐,陳睦虎
(西南醫科大學附屬醫院1.急診科,2.甲狀腺外科,四川 瀘州 646000)
肺巨噬細胞是免疫細胞的一種,能夠吞噬外來病原體及人體衰老的細胞,同時肺巨噬細胞參與特異性免疫,刺激細胞因子的分泌[1-2]。多種肺疾病的病理特征均由炎癥細胞及細胞因子導致,在其發病過程中,肺巨噬細胞發揮著重要作用。目前認為肺巨噬細胞在一些細菌或有害物質的刺激下,分泌大量炎癥因子,部分因子又可趨化中性粒細胞等向肺部遷移,最終引起氣道炎癥或損傷,加重疾病[3]。因此,有效抑制炎癥因子的釋放對肺疾病的治療具有重要意義。自噬是調節肺巨噬細胞吞噬、殺傷,以及調節細胞因子的重要機制,能夠調控機體免疫系統[4]。本課題組前期研究發現,脂多糖(Lipopolysaccharide,LPS)能誘導肺巨噬細胞自噬。目前有多項研究表明,肺損傷及炎癥反應與肺巨噬細胞炎性小體有關,因此推測肺巨噬細胞的自噬過程能夠調控炎癥反應[5-6]。二肽基肽酶-4(dipeptidyl peptidase-4,DPP-4)在肺巨噬細胞中大量表達,參與細胞免疫、自噬等多種生物學過程,目前DPP-4 對趨化因子及細胞因子的調控作用已得到證實[7-8]。本研究探究DPP-4 通過CXCR4/mTOR 通路對小鼠肺巨噬細胞自噬的機制,現報道如下。
1 材料與方法
1.1 細胞來源及培養
小鼠肺泡巨噬細胞MH-S 購自上海酶研生物科技有限公司。取出液氮凍存的MH-S 細胞,恢復至室溫后轉移至25 mL 培養瓶,同時加入15 mL 含10%胎牛血清的達爾伯克改良伊格爾培養基(Dulbecco's modified Eagle's medium,DMEM)培養液,在37℃、5%二氧化碳培養箱中培養24 h。
1.2 主要試劑及儀器
1.2.1 主要試劑DPP-4 表達病毒、si-DPP-4 病毒載體(上海鈺博生物科技有限公司),白細胞介素1β(Interleukin-1β,IL-1β)、白細胞介素6(Interleukin-6,IL-6)及腫瘤壞死因子-α(tumor necrosis factor-ɑ,TNF-ɑ)酶聯免疫試劑盒(上海廣銳生物科技有限公司),轉染試劑盒(上海研卉生物科技有限公司),TRIzol、逆轉錄試劑盒(上海雅吉生物科技有限公司),實時熒光定量聚合酶鏈反應(quantitative realtime polymerase chain reaction,qRT-PCR)試劑盒(上海一研生物科技有限公司),DPP-4、mTOR 兔抗大鼠一抗(1 mg/mL)及山羊抗兔二抗(1 mg/mL)(上海圻明生物科技有限公司),CXCR4兔抗大鼠一抗(1 mg/mL)及山羊抗兔二抗(1 mg/mL)(上海康朗生物科技有限公司)。
1.2.2 主要儀器倒置熒光顯微鏡(型號:Echo Revolve,北京普邁精醫科技有限公司),酶標儀(型號:LONZA ELx808,北京澤平科技有限責任公司),全自動生化分析儀(型號:PUZS-300,上海帝博思生物科技有限公司)。
1.3 細胞分組及干預
取對數生長期的MH-S 細胞,接種于6 孔板,每孔取1×105個細胞繼續貼壁生長,當細胞融合75%~85%時進行轉染。分別設置PBS 組、LPS 組、DPP-4 組,si-DPP-4 組、DPP-4 + BafA1 組及si-DPP-4+BafA1 組。
PBS 組僅為PBS 培養的MH-S 細胞;LPS 組在MH-S 細胞中加入100 ng/mL LPS,誘導培養24 h;DPP-4 組在LPS 組的基礎上,加入100 nmol/L DPP-4表達病毒轉染MH-S 細胞;si-DPP-4 組在LPS 組的基礎上,加入100 nmol/L si-DPP-4 病毒載體轉染MH-S 細胞;DPP-4 + BafA1 組 在LPS 組的基礎上,在DPP-4 表達病毒轉染MH-S 細胞中加入自噬抑制劑BafA1 干預;si-DPP-4 + BafA1 組在LPS 組的基礎上,在si-DPP-4 病毒載體轉染的MH-S 細胞中加入自噬抑制劑BafA1 干預。
1.4 細胞轉染
根據細胞分組進行操作,每孔加入1 mL 病毒稀釋液(病毒感染復數=100,DMEM 培養液1∶99),具體操作步驟參考轉染試劑盒說明書,轉染48 h 后,采用綠色熒光蛋白(green fluorescent protein,GFP)檢測轉染效率,當轉染效率>70%時,可用于后續實驗[9]。
1.5 酶聯免疫吸附試驗檢測MH-S 細胞上清液炎癥因子水平
取各組MH-S 細胞培養液,離心取上清液,采用酶聯免疫吸附試驗試劑盒檢測MH-S 細胞IL-1β、IL-6 及TNF-ɑ 水平,向稀釋后的標準品中加入MH-S 細胞上清液50 μL,混勻后于恒溫水浴鍋中溫育0.5 h,分別加入50 μL 酶標志物、50 μL 顯色劑A、50 μL 顯色劑B 及50 μL 終止液,用酶標儀檢測各孔450 nm 處吸光度,根據IL-1β、IL-6 及TNF-ɑ 的標準曲線計算其相應濃度[10]。每組有4 個復孔,實驗均重復3 次,取均值。
1.6 腺病毒檢測MH-S細胞自噬流變化
取各組穩定轉染的MH-S 細胞,接種于12 孔板上,孔內鋪放細胞爬片,加入攜帶mRFP-GFP-LC3的腺病毒,進行貼壁生長,當細胞融合度為70%時,移除上清液,PBS 沖洗3 次,加入4%多聚甲醛固定,再沖洗3 次,加入抗熒光猝滅封片液進行封片,于熒光顯微鏡下觀察自噬流的變化,即GFP、紅色熒光蛋白(red fluorescent protein,RFP)及其合并通道(黃色)[11]。GFP 減弱表明自噬小體與溶酶體融合,形成自噬溶酶體。
1.7 qRT-PCR 檢 測DPP-4、CXCR4、mTOR mRNA的表達
取各組穩定轉染的MH-S 細胞,采用TRIzol 試劑盒提取MH-S 細胞總RNA,檢測RNA 濃度及純度。采用逆轉錄試劑盒,以RNA 為模板,通過逆轉錄得到cRNA。設置20 μL 的PCR 反應體系:cDNA模板1 μL,SYBR 10 μL,正、反向引物各0.5 μL,純水8 μL;反應體系設置完成后進行擴增,反應條件:95℃預變性2 min,95℃變性30 s,60℃退火30 s,72℃延伸30 s,共40 個循環。根據2-ΔΔCt法計算DPP-4、CXCR4、mTOR 相對表達量[12],引物序列見表1。

表1 qRT-PCR引物序列
1.8 Western blottig 檢測CXCR4/mTOR 通路蛋白的表達
取各組穩定轉染的MH-S 細胞,加入適量裂解液裂解混勻,裂解MH-S 細胞,1 500 r/min 離心20 min,取上清液進行免疫凝膠電泳,分離CXCR4、mTOR 蛋白條帶,并采用濕膜法轉移到PVDF 膜上,5%脫脂奶粉封閉2 h,將PVDF 膜分別在含有一抗(1∶500 稀釋)及二抗(1∶1 000 稀釋)的孵育盒中孵育,孵育后PBS 沖洗3 次進行顯影、定影,分析相應灰度值,目的蛋白相對表達量=目的蛋白/β-actin 灰度值[13]。
1.9 統計學方法
數據分析采用SPSS 21.0 統計軟件。計量資料以均數±標準差(±s)表示,比較用方差分析,進一步兩兩比較用LSD-t檢驗。P<0.05 為差異有統計學意義
2 結果
2.1 MH-S細胞轉染情況
正常MH-S 細胞形態均勻,呈梭形,而轉染后細胞一般成簇存在,呈漩渦樣生長,經綠色熒光蛋白標記后,細胞在熒光顯微鏡下呈綠色熒光。見圖1。
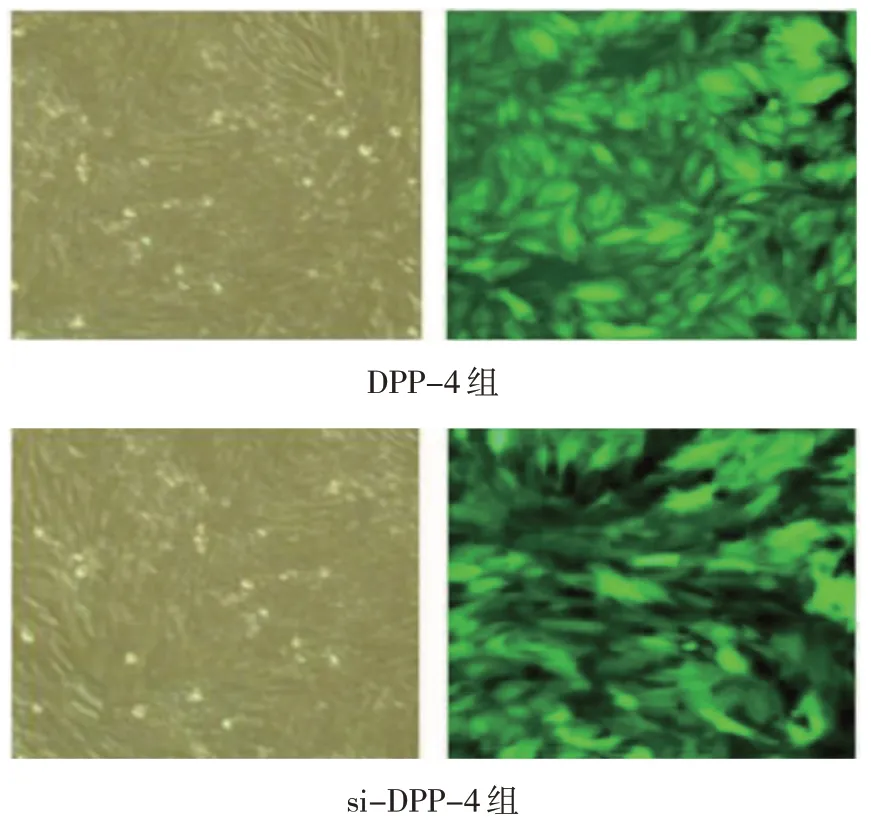
圖1 MH-S細胞轉染圖 (左圖:×100,右圖:×200)
2.2 各組MH-S細胞上清液炎癥因子水平比較
各組IL-1β、IL-6 及TNF-α 水平比較,經方差分析,差異有統計學意義(P<0.05)。進一步兩兩比較結果:LPS 組IL-1β、IL-6 及TNF-α 高于PBS 組(P<0.05);DPP-4 組與LPS 組IL-1β、IL-6 及TNF-α 水平比較,差異無統計學意義(P>0.05);si-DPP-4 組IL-1β、IL-6 及TNF-α 高于LPS 組(P<0.05);si-DPP-4組和DPP-4 + BafA1 組IL-1β、IL-6 及TNF-α 高于DPP-4 組(P<0.05);si-DPP-4 + BafA1 組IL-1β、IL-6高于si-DPP-4 組(P<0.05)。見表2。
表2 各組MH-S細胞上清液炎癥因子水平比較(ng/mL,±s)
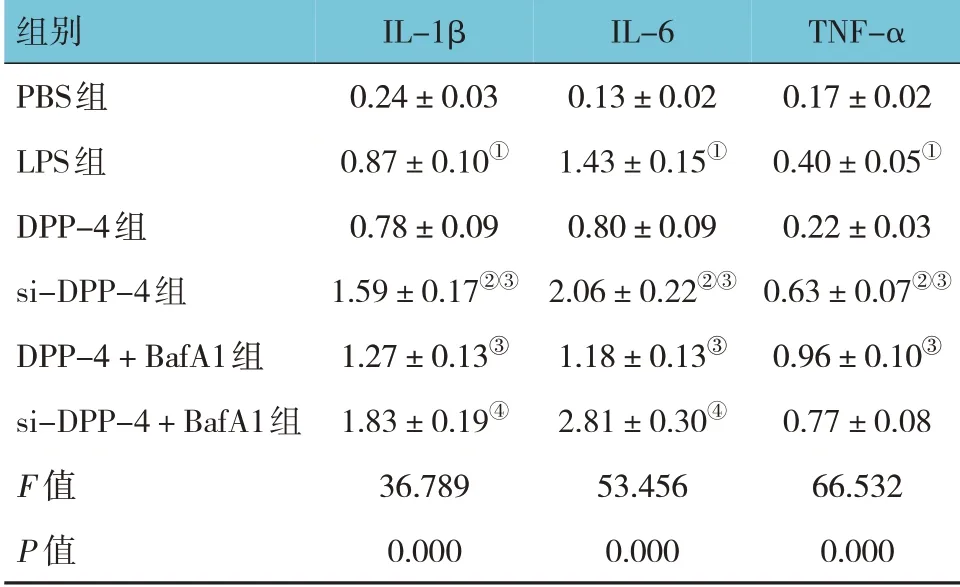
表2 各組MH-S細胞上清液炎癥因子水平比較(ng/mL,±s)
注:①與PBS組比較,P<0.05;②與LPS組比較,P<0.05;③與DPP-4組比較,P<0.05;④與si-DPP-4組比較,P<0.05。
TNF-α 0.17±0.02 0.40±0.05①0.22±0.03 0.63±0.07②③0.96±0.10③0.77±0.08 66.532 0.000組別PBS組LPS組DPP-4組si-DPP-4組DPP-4+BafA1組si-DPP-4+BafA1組F 值P 值IL-1β 0.24±0.03 0.87±0.10①0.78±0.09 1.59±0.17②③1.27±0.13③1.83±0.19④36.789 0.000 IL-6 0.13±0.02 1.43±0.15①0.80±0.09 2.06±0.22②③1.18±0.13③2.81±0.30④53.456 0.000
2.3 各組MH-S細胞自噬流的變化
各組MH-S 細胞GFP、RPF 及Merge 數量比 較,經方差分析,差異有統計學意義(P<0.05)。進一步兩兩比較結果:LPS 組GFP、RPF 及Merge 數量高于PBS 組(P<0.05);DPP-4 組GFP、RPF 及Merge 數量高于LPS 組(P<0.05);si-DPP-4 組GFP、RPF 及Merge 數量低于LPS 組(P<0.05);si-DPP-4 組和DPP-4+BafA1 組GFP、RPF 及Merge 數量低于DPP-4組(P<0.05);si-DPP-4 + BafA1 組GFP、RPF 及Merge數量低于si-DPP-4 組(P<0.05)。見表3和圖2。

圖2 各組MH-S細胞自噬流的變化 (×400)
表3 各組MH-S細胞GFP、RPF及Merge數量比較 (個/細胞,±s)

表3 各組MH-S細胞GFP、RPF及Merge數量比較 (個/細胞,±s)
注:①與PBS組比較,P<0.05;②與LPS組比較,P<0.05;③與DPP-4組比較,P<0.05;④與si-DPP-4組比較,P<0.05。
組別PBS組LPS組DPP-4組si-DPP-4組DPP-4+BafA1組si-DPP-4+BafA1組F 值P 值Merge 104.83±10.68 171.66±17.83①199.67±20.56②97.68±9.96②③138.34±15.75③77.98±8.02④318.456 0.000 GFP 106.68±0.07 169.70±0.08①221.64±0.17②95.75±0.04②③133.58±0.17③79.45±8.17④364.897 0.000 RPF 156.97±16.86 201.55±22.64①274.75±30.54②138.73±14.47②③189.29±18.68③119.68±21.65④495.588 0.000
2.4 各組MH-S 細胞DPP-4、CXCR4、mTOR mRNA相對表達量比較
各組DPP-4、mTOR、CXCR4 mRNA 相對表達量比較,經方差分析,差異有統計學意義(P<0.05)。進一步兩兩比較結果:LPS 組CXCR4、mTOR mRNA相對表達量高于PBS 組(P<0.05);DPP-4 組DPP-4 mRNA 相對表達量高于LPS 組(P<0.05),CXCR4、mTOR mRNA 相對表達量低于LPS 組(P<0.05);si-DPP-4 組DPP-4 mRNA 相對表達量低于DPP-4 組(P<0.05),CXCR4、mTOR mRNA 相對表達量高于DPP-4 組(P<0.05);DPP-4 + BafA1 組CXCR4、mTOR mRNA 相對表達量高于DPP-4 組(P<0.05);si-DPP-4 + BafA1 組CXCR4、mTOR mRNA 相對表達量高于si-DPP-4 組(P<0.05)。見表4。
表4 各組M H-S細胞DPP-4、CXCR4、mTOR mRNA相對表達量比較 (±s)

表4 各組M H-S細胞DPP-4、CXCR4、mTOR mRNA相對表達量比較 (±s)
注:①與PBS組比較,P<0.05;②與LPS組比較,P<0.05;③與DPP-4組比較,P<0.05;④與si-DPP-4組比較,P<0.05。
mTOR mRNA 0.83±0.09 1.26±0.13①1.02±0.10②1.58±0.16②③1.34±0.13③1.86±0.18④49.656 0.000組別PBS組LPS組DPP-4組si-DPP-4組DPP-4+BafA1組si-DPP-4+BafA1組F 值P 值DPP-4 mRNA 0.68±0.07 0.70±0.08 1.64±0.17②0.33±0.04②③1.58±0.17 0.35±0.04 132.456 0.000 CXCR4 mRNA 0.74±0.08 1.15±0.12①0.92±0.10②1.47±0.15②③1.23±0.13③1.73±0.18④73.456 0.000
2.5 各組MH-S 細胞CXCR4/mTOR 通路蛋白相對表達量比較
各組CXCR4、p-mTOR/mTOR 蛋白相對表達量比較,經方差分析,差異有統計學意義(P<0.05)。進一步兩兩比較結果:LPS 組CXCR4、p-mTOR/mTOR 蛋白相對表達量高于PBS 組(P<0.05);DPP-4組CXCR4、p-mTOR/mTOR 蛋白相對表達量低于LPS組(P<0.05);si-DPP-4 組 和DPP-4 + BafA1 組CXCR4、p-mTOR/mTOR 蛋白相對表達量高于DPP-4組(P<0.05);si-DPP-4 + BafA1 組CXCR4、p-mTOR/mTOR 蛋白相對表達量高于si-DPP-4 組(P<0.05)。見表5和圖3。

圖3 各組MH-S細胞CXCR4/mTOR通路蛋白的表達
表5 各組MH-S細胞CXCR4/mTOR通路蛋白相對表達量比較 (±s)

表5 各組MH-S細胞CXCR4/mTOR通路蛋白相對表達量比較 (±s)
注:①與PBS組比較,P<0.05;②與LPS組比較,P<0.05;③與DPP-4組比較,P<0.05;④與si-DPP-4組比較,P<0.05。
p-mTOR/mTOR蛋白0.28±0.03 0.48±0.05①0.31±0.04②0.87±0.09②③0.83±0.09③0.97±0.10④104.567 0.000組別PBS組LPS組DPP-4組si-DPP-4組DPP-4+BafA1組si-DPP-4+BafA1組F 值P 值CXCR4蛋白0.23±0.04 0.62±0.07①0.47±0.05②0.79±0.09②③0.81±0.09③0.93±0.10④89.457 0.000
3 討論
細胞自噬是機體重要的生物學過程,能夠調節細胞代謝平衡,促進增殖分化等,在非生理狀態下自噬作用被激活,能夠調控多種細胞因子,發揮保護細胞的作用[14]。肺巨噬細胞在免疫應答過程中發揮重要作用。多數研究認為,肺巨噬細胞的自噬作用在動脈粥樣硬化、塵肺及癌癥等多種疾病中均有積極作用[15]。DPP-4 在肺巨噬細胞中大量表達,在細胞自噬過程中具有一定作用。在鄭文彬[16]的研究中,DPP4 抑制劑(西格列汀)具有降糖作用,同時還可以通過抑制NF-κB 和JNK/AP1 通路減輕肝臟炎癥,上調AMPK/mTOR 信號通路增強細胞自噬作用,進而改善肝脂肪變性,表明DPP-4 參與細胞炎癥及自噬作用的調節過程。國外有研究顯示,DPP-4 對mTOR 通路有一定的調控作用[17],因此推測DPP-4可能通過mTOR 通路介導小鼠肺泡巨噬細胞MH-S自噬。CXCL12 是DPP-4 底物中的一種,而CXCR4是CXCL12 的特異性受體,在肺巨噬細胞中高表達,相關研究表明,CXCR4/mTOR 信號能夠對腫瘤相關巨噬細胞發揮調控作用[18]。
在馬曉娟等[19]研究中顯示,DPP-4 抑制劑能有效降低血糖并抑制血清肺巨噬細胞移動抑制因子水平,從而發揮抗炎作用,對糖尿病合并冠狀動脈粥樣硬化性心臟病患者起到一定的保護作用。黃玉芳等[20]研究顯示,DPP-4 抑制劑能夠下調小鼠肺巨噬細胞系RAW264.7 細胞的炎癥反應,其作用機制與PKA/NF-κB 通路有關。上述研究均顯示,DPP-4 對小鼠肺巨噬細胞具有一定的抗炎作用。本研究中同樣對各組MH-S 細胞炎癥因子水平進行檢測,其中LPS 組IL-1β、IL-6 及TNF-α 明顯高于PBS組,表明MH-S 細胞在LPS 的誘導下激活細胞炎癥反應,釋放大量炎癥因子,分析其原因為LPS 是內毒素的主要成分,能夠與細胞中多種受體結合,誘導細胞炎癥反應的發生。與LPS 組比較,DPP-4 組炎癥因子水平呈下降趨勢,但無統計學意義,而si-DPP-4 組炎癥因子水平升高,說明DPP-4 對細胞炎癥水平具有一定的抑制作用,但抑制作用較弱,導致各組炎癥因子變化不明顯,而抑制DPP-4 表達能夠促進炎癥反應的發生。與DPP-4 組比較,si-DPP-4 組和DPP-4 + BafA1 組IL-1β、IL-6 及TNF-α 升高,提示抑制DPP-4 表達能夠促進炎癥反應,DPP-4 在自噬抑制劑的作用下,同樣有促炎作用,反向說明DPP-4 高表達和細胞自噬均有抗炎作用。與si-DPP-4 組比較,si-DPP-4 + BafA1 組IL-1β、IL-6 降低,表明在DPP-4 低表達情況下,抑制細胞自噬作用相較于炎癥反應的促進作用更強,進一步說明DPP-4 高表達與細胞自噬均有抗炎作用。
CXCL12/CXCR4 通路在腫瘤轉移、炎癥、免疫等過程中發揮重要作用。國外有研究顯示,抑制DPP-4表達能夠通過CXCL12/CXCR4/mTOR 通路促進上皮間質轉化,加速乳腺癌細胞轉移,且能夠增加乳腺癌細胞的化療耐藥性[21-22]。本研究對MH-S 細胞自噬流、CXCR4/mTOR 通路mRNA 和蛋白進行檢測,結果顯示,經過LPS 誘導,細胞自噬流升高,mTOR mRNA、CXCR4 mRNA、p-mTOR 蛋白、CXCR4 蛋白表達上調,表明在炎癥反應的刺激下,CXCR4/mTOR 通路部分被激活,而在DPP-4 過表達的情況下,自噬流進一步升高,通路mRNA 和蛋白表達進一步上調,表明DPP-4 能夠有效激活CXCR4/mTOR 通路,促進細胞自噬;而在DPP-4 低表達的情況下,自噬作用受到抑制。另外本研究在DPP-4 過表達及低表達的情況下給予自噬抑制劑,結果顯示,兩種情況下細胞自噬流及通路蛋白表達均下調,進一步說明DPP-4 能夠通過CXCR4/mTOR 通路對MH-S 細胞自噬作用進行調控。
綜上所述,DPP-4 能夠通過調控小鼠肺巨噬細胞自噬作用影響細胞炎癥因子分泌,其作用機制與CXCR4/mTOR 通路有關,未來可對DPP4 抑制劑的抗炎作用進行深入研究,為炎癥性疾病的治療提供新的研究思路。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
