pcDNA3.1-HisB-MEG3 體外表達(dá)載體的構(gòu)建及分析驗(yàn)證
2022-09-03 11:36:04宋昕宇羅錦堂郭若楠
天津農(nóng)業(yè)科學(xué) 2022年8期
關(guān)鍵詞:研究
宋昕宇,羅錦堂,郭若楠,李 新
(天津農(nóng)學(xué)院 動(dòng)物科學(xué)與動(dòng)物醫(yī)學(xué)學(xué)院/天津市農(nóng)業(yè)動(dòng)物繁育與健康養(yǎng)殖重點(diǎn)實(shí)驗(yàn)室,天津 300384)
在生命過程中起著至關(guān)重要作用的生物大分子及其結(jié)構(gòu)、功能和組成,一直以來都是科學(xué)家們熱忱探索的課題。生命的基本過程就是從DNA 轉(zhuǎn)錄成RNA,再翻譯成蛋白質(zhì),從而發(fā)揮生物學(xué)功能的過程,其中RNA 的編碼尤為重要。科學(xué)家們把RNA 分成編碼RNA 和非編碼RNA(Non-coding RNA),其中長鏈非編碼RNA(Long noncoding RNA,lncRNA)占將近98%。近十年來,隨著對(duì)lncRNA 研究的深入,越來越多的研究成果證明了lncRNA 不是基因組上的“ 噪音”,其在諸多生命活動(dòng)進(jìn)程和新陳代謝過程中具有極其重要的作用。
作為一個(gè)已有大量研究的lncRNA,MEG3(Maternally Expressed Gene 3),又 稱GTL2 (gene trap locus 2)基因,于2000 年首次被Miyoshi 等人報(bào)道,是一個(gè)lncRNA,并且是一個(gè)母系印記基因。MEG3一般位于染色體末端Dlk1-Dio3 印跡區(qū)內(nèi),人的14q32 染色體上。在小鼠上位于12 號(hào)染色體,在綿羊上位于18 號(hào)染色體,在牛上位于21 號(hào)染色體,并且小鼠MEG3 基因與人類具有同源性。
在人類醫(yī)學(xué)方面,己有研究表明lncRNA MEG3在2 型糖尿病患者胰島中表達(dá)異常。同時(shí),MEG3是第一個(gè)被發(fā)現(xiàn)具有腫瘤抑制功能的lncRNA,它在人類的正常組織中廣泛表達(dá)。MEG3 還可以通過多種途徑抑制細(xì)胞增殖,是一個(gè)重要的腫瘤抑制因子,與多種惡性腫瘤的發(fā)病機(jī)制有關(guān)。
在維持動(dòng)物有機(jī)體骨骼肌發(fā)育發(fā)面,MEG3 同樣扮演著重要的調(diào)控角色,它可以顯著促進(jìn)衛(wèi)星細(xì)胞的分化,調(diào)控骨骼肌的發(fā)育過程。MEG3 在小鼠骨骼肌發(fā)育過程中呈現(xiàn)顯著變化,它對(duì)C2C12 細(xì)胞的分化具有促進(jìn)作用,而過表達(dá)MEG3 對(duì)C2C12 細(xì)胞的增殖具有抑制作用。MEG3 可以促進(jìn)小鼠、豬、牛、羊等動(dòng)物肌肉細(xì)胞的分化,抑制細(xì)胞增殖,調(diào)節(jié)肌肉分化抑制因子。因此,MEG3 可作為畜牧業(yè)生產(chǎn)應(yīng)用的分子標(biāo)記。
有研究發(fā)現(xiàn),lncRNA 在氧化應(yīng)激反應(yīng)中起到一定的作用,然而,關(guān)于lncRNA 在氧化損傷小鼠肌肉中表達(dá)情況尚不清楚。羅海靜等挑選了5 種與氧化應(yīng)激有關(guān)的LncRNA,試驗(yàn)結(jié)果發(fā)現(xiàn)lncRNA MEG3 在氧化損傷小鼠肌肉中的表達(dá)最高,這一研究充分證明了lncRNA MEG3 對(duì)損傷肌肉的修復(fù)起著很大作用。楊睿分別構(gòu)建了2 種MEG3 的體外表達(dá)載體pcDNA3.1-MEG3-TTCC和pcDNA3.1-MEG3-CCCA,并將這2 種體外表達(dá)載體瞬時(shí)轉(zhuǎn)染至豬骨骼肌衛(wèi)星細(xì)胞中,再對(duì)lncRNA MEG3 的相對(duì)表達(dá)量進(jìn)行檢測(cè),結(jié)果顯示這2 種體外表達(dá)載體均能在體外進(jìn)行表達(dá),且pcDNA3.1-MEG3-TTCC 組的表達(dá)量顯著高于pcDNA3.1-MEG3-CCCA 組。該研究說明,這2 種MEG3 的體外表達(dá)載體都能在骨骼肌衛(wèi)星細(xì)胞增殖和分化過程中起著重要的調(diào)控作用。這一試驗(yàn)為lncRNA MEG3 對(duì)豬骨骼肌生長發(fā)育的調(diào)控作用的研究提供了充足的理論依據(jù)。
因此,無論是在癌癥或腫瘤等疾病方面,還是在骨骼肌的分化和發(fā)育方面,lncRNA MEG3 都扮演著重要的角色。在前期的研究中,筆者對(duì)牛骨骼肌衛(wèi)星細(xì)胞進(jìn)行了轉(zhuǎn)錄組學(xué)、翻譯組學(xué)及微肽組學(xué)3 個(gè)組學(xué)的測(cè)序分析,經(jīng)過多組學(xué)聯(lián)合分析發(fā)現(xiàn)MEG3 具有小的開放閱讀框(sORF)結(jié)構(gòu),并在微肽組學(xué)中發(fā)現(xiàn)與之預(yù)測(cè)氨基酸序列對(duì)應(yīng)的微肽產(chǎn)物,因此,初步判斷MEG3 可能具有編碼能力,可以編碼產(chǎn)生功能性微肽。鑒于此,我們?cè)O(shè)計(jì)擴(kuò)增了牛MEG3 基因ORF 區(qū)的編碼序列,構(gòu)建了融合蛋白載體pcDNA3.1-HisB-MEG3,把重組質(zhì)粒轉(zhuǎn)染至293T 細(xì)胞,檢測(cè)其外源表達(dá)情況,為后續(xù)開展更深入的MEG3 功能探究提供材料和夯實(shí)基礎(chǔ),同時(shí)也為深入篩選驗(yàn)證具有編碼產(chǎn)物的lncRNA 提供技術(shù)支持和佐證材料。
1 材料和方法
1.1 細(xì)胞來源
293T 細(xì)胞、牛骨骼肌衛(wèi)星細(xì)胞(BSMSCs)均為天津市農(nóng)業(yè)動(dòng)物繁育與健康養(yǎng)殖重點(diǎn)實(shí)驗(yàn)室凍存。
1.2 質(zhì)粒
質(zhì)粒載體為帶有His 表達(dá)標(biāo)簽的pcDNA3.1-HisB(圖1),購自武漢淼靈生物科技有限公司。

圖1 空載質(zhì)粒pcDN3.1-HisB 基本結(jié)構(gòu)圖
1.3 試劑與耗材
試驗(yàn)用主要試劑和耗材見表1。

表1 試驗(yàn)所需主要試劑和耗材
1.4 儀器設(shè)備
CO培養(yǎng)箱(Thermo-scientific);Light Cycle 96熒光定量PCR 儀(Roche);超敏電泳凝膠成像分析系統(tǒng)(Bio-Rad);制冰機(jī)(SANYO);電泳儀(Bio-Rad);恒溫循環(huán)水浴鍋(德國勞達(dá)貿(mào)易有限公司);高壓滅菌鍋(TOMY);渦旋振蕩器(德國艾卡公司);電子天平(美國奧豪斯公司);超純水制造系統(tǒng)(優(yōu)普);恒溫?fù)u床(IKA)。
1.5 核苷酸序列
依據(jù)轉(zhuǎn)錄組學(xué)、翻譯組學(xué)及微肽組學(xué)多組學(xué)分析結(jié)果獲得MEG3 的ORF 核酸序列信息,體外合成末端帶有XhoI 和XbaI 酶切位點(diǎn)的目標(biāo)片段序列。目的片段由金開瑞生物科技有限公司合成。
1.6 MEG3 外源表達(dá)載體構(gòu)建
將MEG3 ORF 區(qū)的體外合成片段與pcDNA3.1-HisB 空載質(zhì)粒分別用限制性核酸內(nèi)切酶XhoI 和XbaI 進(jìn)行雙酶切處理(酶切體系見表2),37 ℃水浴20 min。酶切產(chǎn)物利用DNA 產(chǎn)物純化試劑盒進(jìn)行純化回收。調(diào)整純化后的目的片段與空載質(zhì)粒pcDNA3.1-HisB 的比例為質(zhì)粒∶目的片段=1∶3,利用T4 連接酶進(jìn)行連接,反應(yīng)條件為22 ℃2 h,4 ℃過夜。隨后將連接產(chǎn)物按照1∶10 的比例加入預(yù)先解凍的感受態(tài)細(xì)胞中進(jìn)行轉(zhuǎn)化。將轉(zhuǎn)化后的細(xì)菌在37 ℃搖床中復(fù)活1 h。隨后鋪30 uL 于Amp 平板上,37 ℃恒溫培養(yǎng)10~12 h。挑取單克隆菌落接種于5 mL含有氨芐青霉素(1∶1 000)的液體培養(yǎng)基中振蕩培養(yǎng),篩選陽性轉(zhuǎn)化子,將驗(yàn)證正確的陽性轉(zhuǎn)化子送至上海生工生物工程有限公司測(cè)序,測(cè)序引物為F:CGCAAATGGGCGGTAGGCGTG;R:TAGAAGGCAC AGTCGAGG。利用DNA MAN 軟件進(jìn)行序列比對(duì)分析,測(cè)序結(jié)果正確的菌液進(jìn)行質(zhì)粒抽提,獲得pcDNA3.1-HisB-MEG3 的體外表達(dá)載體。

表2 DNA/pcDNA3.1-HisB 酶切體系
1.7 目標(biāo)微肽體外表達(dá)
1.7.1 細(xì)胞培養(yǎng) 本研究使用293T 細(xì)胞系進(jìn)行MEG3 的體外表達(dá),使用BSMSCs 進(jìn)行RNA 水平的定量檢測(cè)。
細(xì)胞復(fù)蘇:取出實(shí)驗(yàn)室凍存的細(xì)胞,37 ℃水浴1~2 min 迅速解凍復(fù)蘇,加入1 mL 完全培養(yǎng)基(20%FBS+DMEM),離心后棄上清,加入3 mL 完全培養(yǎng)基重懸細(xì)胞,轉(zhuǎn)入60 mm 培養(yǎng)皿中,在37 ℃、5%CO培養(yǎng)箱中培養(yǎng),觀察細(xì)胞數(shù)量及生長情況。
細(xì)胞傳代:當(dāng)細(xì)胞生長至匯合率達(dá)到80%~90%時(shí)對(duì)細(xì)胞進(jìn)行傳代。棄培養(yǎng)基,加入1 mL 胰酶,37 ℃靜置消化2 min,加入1 mL 完全培養(yǎng)基中和胰酶,輕吹重懸細(xì)胞,吸取細(xì)胞懸液離心,棄上清,加入完全培養(yǎng)基重懸細(xì)胞,按照適當(dāng)比例接種于細(xì)胞培養(yǎng)板中。持續(xù)觀察細(xì)胞狀態(tài)。
1.7.2 細(xì)胞轉(zhuǎn)染 待293T 細(xì)胞融合度達(dá)到80%,依據(jù)Lipofectamine 3000 轉(zhuǎn)染試劑的說明書要求轉(zhuǎn)染pcDNA3.1-HisB-MEG3 質(zhì)粒和對(duì)照組pcDNA3.1-HisB 空質(zhì)粒(NC)。每組試劑設(shè)3 個(gè)生物學(xué)重復(fù)。
使用opti-MEM 培養(yǎng)基稀釋lipofectamine 3000、P3000 和載體質(zhì)粒。293T 細(xì)胞由無抗培養(yǎng)基培養(yǎng)。12 孔板1 孔配置方法A:62.5 μL opti-MEM+3.75 μL lip 3000;B:62.5 μL opti-MEM +2.5 μL P3000+cDNA3.1-HisB-MEG3/pcDNA3.1-HisB 空質(zhì)粒(NC),加入1 250 ng 質(zhì)粒。將上述試劑室溫孵育5 min,隨后A、B 兩管中試劑混合均勻再次孵育10 min。加入到293T 細(xì)胞培養(yǎng)皿中輕晃搖勻。轉(zhuǎn)染6 h 后更換為有抗培養(yǎng)基,轉(zhuǎn)染后24 h 分別收集細(xì)胞mRNA 和蛋白樣品,進(jìn)行后續(xù)分析。
1.8 目標(biāo)微肽驗(yàn)證與鑒定
1.8.1 細(xì)胞總RNA 提取及外源微肽mRNA 表達(dá)鑒定 采用RNA 快速提取試劑盒分別抽提上述轉(zhuǎn)染了pcDNA3.1-HisB-MEG3 質(zhì)粒和對(duì)照組pcDNA3.1-HisB 空質(zhì)粒的293T 細(xì)胞(24 孔板中培養(yǎng))的總RNA,按照HiFiScript cDNA Synthesis Kit 說明書進(jìn)行cDNA 的合成,反轉(zhuǎn)錄體系見表3。反應(yīng)程序?yàn)椋?2 ℃15 min,85 ℃5 min。

表3 反轉(zhuǎn)錄體系
以GAPDH 為內(nèi)參,利用實(shí)時(shí)熒光定量PCR(qRT-PCR)檢測(cè)MEG3 融合蛋白的mRNA 表達(dá)情況。引物序列F:ATGTTTTTCCCAAACAGGTTGGTC;R:TTATTGGAGCTCGGAGACGG。每組試驗(yàn)設(shè)置3個(gè)生物學(xué)重復(fù)。
1.8.2 細(xì)胞總蛋白提取及Western blot 分析 利用RIPA 蛋白裂解液裂解并收集分別轉(zhuǎn)染pcDNA3.1-HisB-MEG3 質(zhì)粒及pcDNA3.1-HisB 空質(zhì)粒(NC)的293T 細(xì)胞的總蛋白樣品。將含有外源表達(dá)蛋白的細(xì)胞總蛋白通過12%SDS-PAGE 凝膠進(jìn)行電泳分離,隨后300 mA 轉(zhuǎn)膜2 h。用5%脫脂奶粉封閉液室溫封閉PVDF 膜2 h,根據(jù)目標(biāo)蛋白分子量大小剪取PVDF 膜,并用適當(dāng)一抗(anti-His 抗體/DAPDH抗體)4 ℃孵育過夜,TBST 洗膜后用熒光標(biāo)記二抗(Secondary Antibodies-Goat Anti-Rabbit Mouse IgGHRP)室溫孵育1 h。按照1∶1 的比例配置ECL 超敏發(fā)光液,并在超敏電泳凝膠成像分析系統(tǒng)下拍照保存。分析目的蛋白表達(dá)情況。
1.9 統(tǒng)計(jì)分析
對(duì)qRT-PCR 結(jié)果進(jìn)行t 檢驗(yàn)分析,按2Ct 計(jì)算,GAPDH 基因作為內(nèi)參對(duì)檢測(cè)的目的基因進(jìn)行歸一化處理。“ *”表示差異顯著(P<0.05),“ **”表示差異極顯著(P<0.01)。
2 結(jié)果與分析
2.1 pcDNA3.1-HisB-MEG3 表達(dá)載體構(gòu)建及鑒定結(jié)果
在前期研究中,筆者團(tuán)隊(duì)發(fā)現(xiàn)lncRNA MEG3 可能具有編碼潛能,能編碼產(chǎn)生微肽,為驗(yàn)證這一結(jié)果,筆者利用前期的微肽組學(xué)結(jié)果和生物信息學(xué)手段,分析獲得了lncRNA MEG3 可能的ORF 區(qū),該ORF 區(qū)長度為555 bp,可能編碼產(chǎn)生長度為185 aa的微肽(圖2)。因此,筆者人工合成了末端帶有XhoI、XbaI 酶切位點(diǎn)的lncRNA MEG3 的ORF 區(qū)序列(圖3),為了方便后續(xù)試驗(yàn),筆者在起始密碼子后同時(shí)添加了6×His 以及3×Flag 小分子標(biāo)簽,再將其連入帶有His 蛋白表達(dá)標(biāo)簽的pcDNA3.1-HisB 質(zhì)粒中,以增加檢測(cè)的敏感性。由圖4 可知,本研究選擇pcDNA3.1-HisB 空載質(zhì)粒作為載體,經(jīng)過XhoI、XbaI 雙酶切,可將目的片段連接在空載質(zhì)粒pcDNA3.1-HisB 上,從而在細(xì)胞中表達(dá)MEG3 蛋白。質(zhì)粒中含有T7 啟動(dòng)子,可大量轉(zhuǎn)錄表達(dá)目標(biāo)蛋白,6×His 標(biāo)簽位于目的基因MEG3 的上游,經(jīng)過PCR擴(kuò)增獲得了全長6 167 bp 的pcDNA3.1-HisB-MEG3融合載體。

圖2 目的片段核苷酸序列

圖3 MEG3 的ORF 區(qū)序列

圖4 pcDNA3.1-HisB-MEG3 重組質(zhì)粒圖譜
為驗(yàn)證pcDNA3.1-HisB-MEG3 融合載體序列的正確性,利用XhoI、XbaI 限制性內(nèi)切酶進(jìn)行質(zhì)粒雙酶切,對(duì)酶切產(chǎn)物及重組質(zhì)粒進(jìn)行凝膠電泳,如圖5 所示,泳道1 是經(jīng)過雙酶切的重組質(zhì)粒,在該泳道中形成一個(gè)長度為644 bp 的帶有目的片段的條帶以及一個(gè)長度為5 523 bp 的不含目的基因的條帶,2 個(gè)條帶的長度均符合要求。由此可以初步判斷,pcDNA3.1-HisB-MEG3 表達(dá)載體構(gòu)建成功。同時(shí),設(shè)計(jì)lncRNA MEG3 ORF 區(qū)序列的特異性引物,對(duì)pcDNA3.1-HisB-MEG3 的融合載體進(jìn)行PCR 擴(kuò)增及測(cè)序分析,測(cè)序結(jié)果如圖6 所示,序列完全匹配,說明pcDNA3.1-HisB-MEG3 表達(dá)載體構(gòu)建成功。
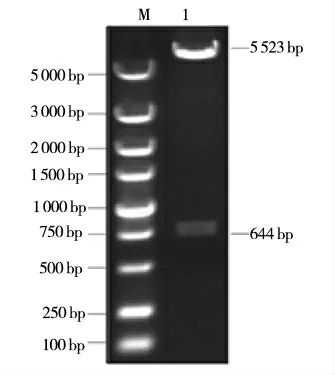
圖5 pcDNA3.1-HisB-MEG 重組質(zhì)粒雙酶切結(jié)果

圖6 pcDNA3.1-HisB-MEG3 測(cè)序結(jié)果
2.2 細(xì)胞中pcDNA3.1-HisB-MEG3 mRNA 的表達(dá)檢測(cè)
為了進(jìn)一步驗(yàn)證pcDNA3.1-HisB-MEG3 在BSMSCs 中的過表達(dá)情況,于BSMSCs 細(xì)胞分別轉(zhuǎn)染pcDNA3.1-HisB 空載體(NC)和pcDNA3.1-HisBMEG3 表達(dá)載體24 h 后,提 取BSMSCs 增殖期總RNA,檢測(cè)與對(duì)照組相比pcDNA3.1-HisB-MEG3 表達(dá)水平的變化。如圖7 所示,外源轉(zhuǎn)入的pcDNA3.1-HisB-MEG3 在BSMSCs 細(xì)胞中其mRNA 表達(dá)量比NC 對(duì)照組高約23 倍,差異極顯著。因此,可以初步判斷在牛骨骼肌衛(wèi)星細(xì)胞中 pcDNA3.1-HisB-MEG3 重組載體mRNA 可以正常表達(dá)。

圖7 MEG3 mRNA 表達(dá)檢測(cè)結(jié)果
2.3 考馬斯亮藍(lán)定性檢測(cè)蛋白的表達(dá)情況
本研究驗(yàn)證了pcDNA3.1-HisB-MEG3 在293T細(xì)胞中的mRNA 表達(dá)水平后,利用考馬斯亮藍(lán)染色對(duì)pcDNA3.1-HisB-MEG3 編碼的微肽在蛋白水平的表達(dá)進(jìn)行了初步檢測(cè)。前期預(yù)測(cè)出MEG3 ORF 序列編碼一個(gè)185 aa 的微肽,將其連入pcDNA3.1-HisB 載體后其編碼的微肽-His 標(biāo)簽融合蛋白的大小約為17 kDa。考染結(jié)果如圖8 所示,帶有目的基因MEG3 的重組質(zhì)粒轉(zhuǎn)染293T 細(xì)胞后,在25~35 kDa 處(圖中方框處)具有明顯的蛋白條帶,而NC組無明顯條帶,這說明pcDNA3.1-HisB-MEG3 在293T 細(xì)胞中表達(dá)了融合蛋白產(chǎn)物,證明其能在蛋白水平進(jìn)行表達(dá)。
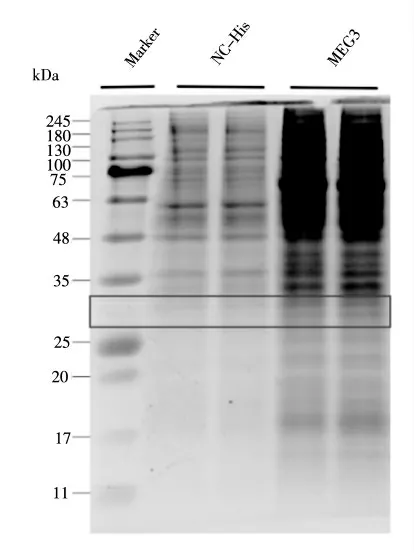
圖8 293T 細(xì)胞中考馬斯亮藍(lán)染色定性檢測(cè)MEG3 的蛋白表達(dá)水平
2.4 Western blot 試驗(yàn)結(jié)果
為進(jìn)一步驗(yàn)證MEG3 微肽融合蛋白的外源表達(dá),利用His 標(biāo)簽蛋白的特異性His 抗體進(jìn)行了Western blotting 分析。結(jié)果如圖9 所示,His 抗體孵育后,在pcDNA3.1-HisB-MEG3 轉(zhuǎn)染組檢測(cè)到明顯的特異性條帶,而NC 對(duì)照組沒有檢測(cè)到條帶,說明本研究構(gòu)建的pcDNA3.1-HisB-MEG3 在293T 細(xì)胞中融合蛋白成功表達(dá)。
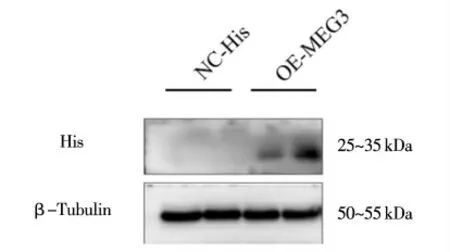
圖9 293T 細(xì)胞中Western blot 檢測(cè)MEG3 的蛋白表達(dá)水平變化
3 結(jié)論與討論
IncRNA 是一類轉(zhuǎn)錄本長度超過200 個(gè)核苷酸的RNA 分子,是轉(zhuǎn)錄組的主要組成部分,它正在成為調(diào)控發(fā)育進(jìn)程中的核心參與者。同時(shí),生物體基因組可以轉(zhuǎn)錄產(chǎn)生數(shù)目巨大、形式各異和功能不同的lncRNA 分子。已有研究結(jié)果表明,轉(zhuǎn)錄lncRNA分子的基因來源于生物體基因組上的許多區(qū)域,并且多數(shù)lncRNA 分子序列上都含有小的、未被注釋過的開放閱讀框序列。越來越多的研究表明這些缺少蛋白編碼潛能的lncRNA 分子可以翻譯成為多肽分子并參與相關(guān)的生命活動(dòng)。
胡艷妹等研究發(fā)現(xiàn),MEG3 在胰島β 細(xì)胞高表達(dá),具有組織特異性。干擾MEG3 后,小鼠葡萄糖耐受能力受損,血清胰島素水平降低,胰島素合成能力下降。初步證實(shí),MEG3 可以通過參與胰島素的合成和分泌來維持成年小鼠胰島功能,進(jìn)一步豐富和完善了胰島發(fā)育和功能維持的基因調(diào)控網(wǎng)絡(luò)。程寧等研究發(fā)現(xiàn),MEG3 可促進(jìn)白細(xì)胞介素1β 誘導(dǎo)的大鼠椎間盤髓核細(xì)胞增殖并抑制細(xì)胞凋亡,揭示了MEG3 對(duì)大鼠椎間盤髓核細(xì)胞增殖及凋亡的分子機(jī)制起著決定性作用,進(jìn)一步奠定了MEG3 的研究基礎(chǔ)。
在骨分化和形成方面,有研究表明,MEG3 的表達(dá)上調(diào)能抑制骨的自噬,從而抑制成骨分化和骨形成代謝。同時(shí),還有不少研究表明,在牛骨骼肌衛(wèi)星細(xì)胞中l(wèi)ncRNA MEG3 能夠促進(jìn)牛原代骨骼肌衛(wèi)星細(xì)胞分化。在對(duì)動(dòng)物生長發(fā)育性狀的研究方面,通過剖析4 個(gè)綿羊品種的MEG3 基因的不同基因型和單倍型對(duì)生長發(fā)育的影響表明,MEG3 基因的不同基因型和單倍型在灘羊的體高、體長和胸圍性狀上存在顯著差異,說明MEG3 對(duì)于綿羊生長發(fā)育性狀有影響。
除了在動(dòng)物上的研究和試驗(yàn),在人類醫(yī)學(xué)以及各種疾病的研究中,大量研究結(jié)果表明,MEG3 在預(yù)測(cè)宮頸癌患者腫瘤大小和淋巴結(jié)轉(zhuǎn)移方面具有足夠的敏感性和特異性,MEG3 在調(diào)節(jié)細(xì)胞增殖和凋亡基礎(chǔ)上與宮頸癌的進(jìn)展有關(guān)。因此,MEG3 可能是宮頸癌的一個(gè)潛在診斷工具和預(yù)后標(biāo)志物。同時(shí),MEG3 甲基化不僅是宮頸癌的危險(xiǎn)因素,也是HR-HPV 感染和淋巴結(jié)轉(zhuǎn)移的危險(xiǎn)因素。更多研究表明,MEG3 在多種癌癥中丟失,包括膠質(zhì)瘤、腦膜瘤、胃癌和膀胱癌。因此,無論是在農(nóng)業(yè)科學(xué)還是人類科學(xué)方面,MEG3 都有著巨大的研究價(jià)值。
為了后續(xù)在lncRNA 中針對(duì)MEG3 基因展開相關(guān)研究,本試驗(yàn)成功構(gòu)建MEG3 的融合蛋白載體pcDNA3.1-HisB-MEG3,并能夠在外源的293T 細(xì)胞中正常表達(dá),為判斷MEG3 對(duì)腫瘤和癌癥及其他疾病中潛在作用的研究夯實(shí)理論基礎(chǔ),同時(shí)也說明lncRNA MEG3 在骨骼肌的生長發(fā)育中起著重要作用,有利于揭示部分非編碼RNA 也具有編碼潛能這一重要論題。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
