脊尾白蝦水通道蛋白基因4和11在堿度脅迫過程中的作用*
2022-09-05 02:14:28李明棟王佳佳葛倩倩李吉濤
漁業(yè)科學(xué)進展 2022年4期
李明棟 王佳佳 葛倩倩 秦 楨 劉 萍 李 健 李吉濤
脊尾白蝦水通道蛋白基因4和11在堿度脅迫過程中的作用*
李明棟1,2王佳佳1,2葛倩倩1,2秦 楨1,2劉 萍1,2李 健1,2李吉濤1,2①
(1. 中國水產(chǎn)科學(xué)研究院黃海水產(chǎn)研究所 農(nóng)業(yè)農(nóng)村部海洋漁業(yè)可持續(xù)發(fā)展重點實驗室 山東 青島 266071;2. 青島海洋科學(xué)與技術(shù)試點國家實驗室海洋漁業(yè)科學(xué)與食物產(chǎn)出過程功能實驗室 山東 青島 266071)
水通道蛋白(aquaporin)是一種細胞膜上特異性轉(zhuǎn)運水分子及其他中性代謝分子的膜蛋白家族,對生物的細胞內(nèi)外滲透壓穩(wěn)定具有重要的調(diào)節(jié)作用。為了了解水通道蛋白在脊尾白蝦()應(yīng)對堿度脅迫中的作用,本研究利用RACE技術(shù)成功克隆了脊尾白蝦水通道蛋白4 (aquaporin 4,)與水通道蛋白11 (aquaporin 11,)基因cDNA全長,基因的開放閱讀框為621 bp,編碼206個氨基酸,預(yù)測分子量為21.673 kDa,理論等電點為8.30,為疏水性蛋白,具有5個跨膜結(jié)構(gòu)域;基因的開放閱讀框長度為783 bp,編碼260個氨基酸,預(yù)測蛋白分子量為28.490 kDa,理論等電點為5.40,為疏水性蛋白,具有4個跨膜結(jié)構(gòu)域。序列比對結(jié)果顯示,基因與羅氏沼蝦()同源性最高,為94.63%;與斑節(jié)對蝦()同源性最高,為81.47%。為驗證水通道蛋白的功能,利用RNA干擾技術(shù)特異性沉默和基因,結(jié)果顯示,碳酸鹽堿度脅迫后,注射干擾后的脊尾白蝦死亡率顯著升高,干擾組72 h死亡率達到45%,干擾組72 h死亡率達到55%,與對照組差異顯著(<0.05)。同時發(fā)現(xiàn),干擾組在碳酸鹽堿度脅迫24、48與72 h時的血液滲透壓變化幅度顯著高于對照組(<0.05),72 h時滲透壓顯著升高(<0.05);干擾組血液滲透壓在3個時間點均顯著升高(<0.05)。以上結(jié)果表明,水通道蛋白在脊尾白蝦應(yīng)對堿度脅迫過程中起到了調(diào)節(jié)滲透壓、維持體內(nèi)外離子平衡的作用。
脊尾白蝦;水通道蛋白;基因克隆;滲透壓
脊尾白蝦()又稱小白蝦、迎春蝦,主要分布于黃渤海淺海低鹽水域,是我國重要的中小型經(jīng)濟蝦類。脊尾白蝦為廣溫、廣鹽、廣布種,具有生長速度快、繁殖周期短、適應(yīng)能力強等優(yōu)點,現(xiàn)已成為我國沿海地區(qū)的特色養(yǎng)殖品種。我國有低洼鹽堿水域4600萬hm2,具有養(yǎng)殖潛力的鹽堿水面積可達667萬hm2,鹽堿水由于具有高pH、高碳酸鹽堿度及離子組成復(fù)雜等特點,阻礙了鹽堿水資源的充分開發(fā)利用。脊尾白蝦作為一種對鹽堿環(huán)境適應(yīng)能力強的經(jīng)濟蝦類,是開展甲殼類鹽堿適應(yīng)分子機制研究的理想材料。
水通道蛋白(aquaporin, AQP)是一種細胞膜上特異性轉(zhuǎn)運水分子及其他中性代謝分子的膜蛋白家族(馮學(xué)超等, 2005; 王渝等, 2014)。目前,共有13種哺乳動物水通道蛋白被鑒定,根據(jù)其功能可分為三類:AQP0、AQP1、AQP2、AQP4和AQP5僅對水分子有通透性,為水選擇性水通道蛋白;AQP3、AQP7、AQP9和AQP10對水、甘油、尿素等均有通透性,為水甘油通道蛋白;AQP11與AQP12的功能還未確定且基因同源性低,為超級水通道蛋白。水通道蛋白對生物的細胞內(nèi)外滲透壓穩(wěn)定具有重要的調(diào)節(jié)作用,其對水產(chǎn)動物的生存意義重大。現(xiàn)階段對水通道蛋白的研究主要集中于植物與哺乳動物,研究表明,植物水通道蛋白可以通過調(diào)節(jié)根部的導(dǎo)水率來增加水分的吸收以響應(yīng)鹽脅迫與干旱脅迫;在哺乳動物中,水通道蛋白在腎臟的表達量最高,在水運輸?shù)闹饕M織中發(fā)揮重要作用(Li, 2019; 段夢莎, 2020; 郝建峰, 2020; 李娟等, 2021; 李青云, 2020; 張亞楠等, 2019)。目前,關(guān)于水產(chǎn)動物水通道蛋白的研究尚少,主要集中于三疣梭子蟹() (王渝等, 2014)、中華絨螯蟹() (楊志剛等, 2021)、薩羅羅非魚() (甘遠迪, 2014)等水產(chǎn)動物中,研究表明,水通道蛋白基因在水產(chǎn)動物的腸道、胃與鰓組織中相對表達量較高,且鹽度脅迫可顯著改變其表達模式,推測水通道蛋白在水產(chǎn)動物滲透壓調(diào)節(jié)過程中起重要作用。本研究根據(jù)前期堿度脅迫下脊尾白蝦的轉(zhuǎn)錄組測序結(jié)果,克隆堿度脅迫下的差異表達基因和,并分析其序列特征;利用RNA干擾技術(shù)特異性地沉默和基因的表達,然后對脊尾白蝦進行堿度脅迫,驗證和基因在堿度脅迫過程中的作用。結(jié)果可為解析脊尾白蝦響應(yīng)堿度脅迫機制提供基礎(chǔ)信息。
1 材料與方法
1.1 實驗材料及處理
實驗用脊尾白蝦平均體長為(2.1±0.4) cm,均取自山東日照海辰水產(chǎn)有限公司。實驗前將脊尾白蝦在100 L的桶中暫養(yǎng)7 d,按時投喂飼料,連續(xù)充氧。
取6尾體長、體重相近且活力旺盛的脊尾白蝦,取其鰓、肝胰腺、肌肉、腸、血細胞和胃組織于液氮中儲存,用于合成RACE模板。
1.2 AQP4與AQP11基因的cDNA全長克隆
使用Up Plus RNA試劑盒(TRAN, 中國)提取脊尾白蝦RNA。配制1.0%的瓊脂糖凝膠進行電泳,檢測RNA的條帶完整性;使用Thermo NanoDrop 2000分光光度計檢測RNA的純度及濃度。RACE模板第一鏈?zhǔn)褂肧MARTer?RACE 5′/3′試劑盒合成。根據(jù)脊尾白蝦轉(zhuǎn)錄組中與的測序結(jié)果,利用Primer Premier 5.0軟件設(shè)計3′與5′引物,引物序列見表1。使用Advantage 2 Polymerase Mix高保真聚合酶依據(jù)說明書進行目的基因RACE 5′和3′末端巢式PCR擴增,將獲得的PCR產(chǎn)物進行切膠回收、連接轉(zhuǎn)化,然后挑取陽性單克隆,經(jīng)M13通用引物進行菌落PCR鑒定,并將所篩選的菌液進行測序。
表1 水通道蛋白基因克隆所用引物序列

Tab.1 The sequence of primers used for AQP gene cloning
1.3 生物信息學(xué)分析
使用Contig Express軟件進行拼接,得到基因與基因cDNA全長,將cDNA序列用ORFFinder在線軟件(https://www.ncbi.nlm.nih.gov/ orffinder/)預(yù)測開放閱讀框,得到基因的氨基酸序列后,使用在線軟件SignalP-5.0 Server (http://www.cbs. dtu.dk/services/SignalP/)、SMART(http://smart.embl- heidelberg.de/)、TMHMM Server v.2.0 (http://www.cbs. dtu.dk/services/TMHMM/)和ProtParam tool(http://web. expasy.org/protparam/)預(yù)測其蛋白質(zhì)信號肽、蛋白質(zhì)結(jié)構(gòu)域、蛋白跨膜情況、蛋白質(zhì)特性和親水性。
利用在線軟件NCBI BLAST (https://blast.ncbi. nlm.nih.gov/Blast.cgi)分析基因與其他物種的同源性和一致性;使用DNAMAN軟件對多個物種氨基酸序列進行比對;使用MEGA 7.0軟件構(gòu)建Neighbor- joining進化樹。
1.4 RNA干擾實驗
1.4.1 dsRNA的合成 使用T7 RNAi Transcription Kit-BOX1 (Vazyme, 中國)合成dsRNA。合成干擾的引物需添加T7啟動子,使用在線設(shè)計軟件https:// www.flyrnai.org/cgi-bin/RNAi_find_primers.pl設(shè)計引物,引物由生工生物工程(上海)有限公司合成。
本次干擾實驗以綠色熒光蛋白(green fluorescent protein (GFP)為對照,具體的合成步驟:將GFP質(zhì)粒溶解,取2 μL質(zhì)粒溶液加入100 μL大腸桿菌() DH5α感受態(tài)細胞,冰浴30 min,42℃熱激90 s,再冰浴2 min,加入900 μL未加卡那霉素(KAN)的LB液體培養(yǎng)基,200 r/min震蕩培養(yǎng)45 min。將菌液6000 r/min離心5 min,取沉淀涂至已加KAN的LB固體培養(yǎng)基上,過夜培養(yǎng)12 h以上。挑取單克隆菌落至已加KAN的LB液體培養(yǎng)基中,震蕩培養(yǎng)12 h。以培養(yǎng)后的菌液為模板,使用T7 RNAi Transcription Kit-BOX1按照dsRNA合成步驟合成GFP,其引物GFP-T7F:GATCACTAATACGACTCACT ATAGGGATGGTGAGCAAGGGGGAGGA;GFP-T7R:GATCACTAATACGACTCACTATAGGGTTACTTGTACAGCTCGTCCA。
1.4.2 dsRNA的注射 準(zhǔn)備9個10 L整理箱,每個箱子加入8 L海水,調(diào)節(jié)其碳酸鹽堿度至8.26 mmol/L,穩(wěn)定24 h,每天調(diào)節(jié)2次海水堿度。設(shè)置GFP組、組和組,每組3個平行。挑選體長、體重相近的脊尾白蝦進行脅迫實驗,每個箱子放入10尾蝦,依據(jù)組別在脊尾白蝦的第4腹足基部注射干擾,注射量為1 μg/g,觀察記錄脊尾白蝦在24、48和72 h的死亡率。
另設(shè)置空白組、GFP組、組和組,調(diào)節(jié)其碳酸鹽堿度至8.26 mmol/L,每個箱子放入30尾脊尾白蝦,依據(jù)組別在脊尾白蝦的第4腹足基部注射干擾,注射量為1 μg/g,空白組不進行脅迫與干擾,在24、48和72 h 3個時間點,取其血液測量滲透壓,并取鰓組織檢測其中基因表達量。
1.4.3 干擾后血淋巴滲透壓的變化 每個時間點取6尾蝦的血淋巴混合測量其滲透壓。將混合后的血淋巴樣品放入4℃冰箱中24 h后,3000 r/min離心15 min,取60 μL上清液,使用滲透壓測定儀測量滲透壓,每個樣品測量3次。
1.4.4 干擾后AQP基因表達量的檢測 在干擾后24、48和72 h 3個時間點取GFP組、組與組的鰓組織,液氮研磨,使用Up Plus RNA kit (TRAN,中國)提取RNA。
通過熒光定量PCR (qRT-PCR)分析與在不同組織中的表達,以18S基因作為內(nèi)參基因,針對、與18S設(shè)計特異性引物(表2),按照ChamQTM SYBR?Color qPCR master mix說明書,利用熒光定量PCR儀器進行定量分析,通過2–ΔΔCt法計算分析注射干擾后與基因mRNA的相對表達量,利用SPSS 17.0軟件進行單因素方差分析(one-way ANOVA),運用多重比較進行顯著性檢驗。
表2 qRT-PCR所用引物

Tab.2 Primers used for qRT-PCR
2 結(jié)果與分析
2.1 EcAQP4與EcAQP11基因cDNA的克隆與序列分析
本研究利用RACE技術(shù)成功克隆了脊尾白蝦與的cDNA全長,序列分析結(jié)果顯示,基因開放閱讀框(ORF)長度為621 bp,共編碼206個氨基酸,預(yù)測蛋白分子量為21.673 kDa,理論等電點為8.30,為疏水性蛋白(圖1);跨膜分析結(jié)果顯示,該基因跨膜5次;SMART分析顯示,該基因具有5個跨膜結(jié)構(gòu)域(圖2)。基因ORF長度為783 bp,共編碼261個氨基酸,預(yù)測其蛋白分子量為28.490 kDa,理論等電點為5.40,為疏水性蛋白(圖3);跨膜分析結(jié)果顯示,該基因跨膜4次;SMART分析顯示,該基因具有4個跨膜結(jié)構(gòu)域(圖4)。
2.2 AQP4與AQP11氨基酸序列及同源性分析
將EcAQP4與ECAQP11的氨基酸序列在NCBI上比對發(fā)現(xiàn),與EcAQP4氨基酸序列同源性較高的物種有羅氏沼蝦()斑節(jié)對蝦()、凡納濱對蝦()、三疣梭子蟹(圖5);與EcAQP11氨基酸序列同源性較高的物種有斑節(jié)對蝦、灰眼雪蟹()、凡納濱對蝦(圖6)。同源性分析顯示,EcAQP4具有多個保守區(qū)域,其中與羅氏沼蝦同源性最高,為94.63%,與三疣梭子蟹同源性最低,為68.14%,所比對的物種均含有HINPAVT和PLAIGL這2個保守域;EcAQP11比EcAQP4保守性較差,與斑節(jié)對蝦同源性最高,達到81.47%,與灰眼雪蟹同源性最低,為67.18%。
2.3 AQP4與AQP11基因系統(tǒng)進化樹分析
利用MEGA 7.0軟件對脊尾白蝦與基因的氨基酸序列進行系統(tǒng)進化分析(圖7,圖8),結(jié)果顯示,基因的進化樹分為兩支,第一支為節(jié)肢動物門,第二支為脊索動物門,脊尾白蝦與羅氏沼蝦聚為一支,親緣關(guān)系最為相近,與其他甲殼綱動物親緣關(guān)系也較近;脊索動物門中的硬骨魚類為一支,與脊尾白蝦親緣關(guān)系較遠。基因的進化樹分為兩支,第一支為昆蟲綱,第二支為甲殼綱,脊尾白蝦與斑節(jié)對蝦與凡納濱對蝦聚為一支,親緣關(guān)系較近。

圖1 EcAQP4基因cDNA全長及推導(dǎo)的氨基酸序列
ATG:起始密碼子;TAA:終止密碼子。圖3同。
ATG: Initiation codon; TAA: Termination codon. The same as in Fig.3.

圖2 EcAQP4的跨膜結(jié)構(gòu)域

圖3 EcAQP11基因ORF及其推導(dǎo)的氨基酸序列

圖4 EcAQP11的跨膜結(jié)構(gòu)域
2.4 RNA干擾后EcAQP4與EcAQP11的表達變化
取干擾后脊尾白蝦的鰓組織,提取RNA,進行反轉(zhuǎn)錄,通過qRT-PCR檢測2個基因在各個時間點的表達量。結(jié)果顯示,注射dsRNA對脊尾白蝦與基因干擾效果明顯。與GFP對照組相比,基因在注射干擾24 h與48 h后的表達量下降80%左右,72 h干擾效果逐漸減弱;基因在注射干擾24 h時表達量下降87%,干擾效果顯著,48 h時基因表達量下降60%,72 h時基因表達量下降35%。
2.5 RNA干擾對脊尾白蝦死亡率的影響
統(tǒng)計每個時間點各組別的死亡率,如圖10所示。與GFP組相比,注射干擾后,與組死亡率顯著升高(<0.05),說明RNA干擾這2個基因表達后,脊尾白蝦耐堿度能力降低,最終導(dǎo)致死亡。同時,發(fā)現(xiàn)24 h與48 h 2個時間點AQP11組死亡率顯著高于AQP4組(<0.05),在72 h時2組死亡率接近,相比基因,基因能夠更加快速地影響脊尾白蝦滲透壓調(diào)節(jié)能力,在脊尾白蝦耐堿機制中起到更為重要的作用。綜上所述,與基因?qū)刮舶孜r耐堿能力有重要的作用。

圖5 EcAQP4基因氨基酸序列與其他已知物種同源序列比對
NPA結(jié)構(gòu)單元以方框表示;2個保守序列HINPAVT和PLAIGL以下劃線表示。
The NPA structural motifs are marked in the box. Two conserved sequences (HINPAVT and PLAIGL) are underlined.
2.6 RNA干擾對脊尾白蝦血液滲透壓的影響
使用滲透壓測量儀測量各組在不同時間點脊尾白蝦的血液滲透壓,結(jié)果如圖11所示。與空白組相比,堿度脅迫后,脊尾白蝦血液滲透壓顯著升高(<0.05),這是由于水體堿度升高,其中的離子濃度也隨之升高,脊尾白蝦體內(nèi)累積的離子升高所導(dǎo)致,但總體來看,GFP組脊尾白蝦血液滲透壓相對穩(wěn)定,呈現(xiàn)先上升后下降的趨勢;干擾組的滲透壓變化趨勢與GFP組相同,在48與72 h時2個時間點其滲透壓高于GFP組;干擾組滲透壓則是一直升高的,72 h時顯著升高(<0.05),升高的幅度較大。推測干擾與基因表達后,水分子的轉(zhuǎn)運受到影響,脊尾白蝦滲透壓調(diào)節(jié)能力降低,造成AQP4組上升與下降趨勢大于對照組;AQP11干擾組脊尾白蝦滲透壓上升明顯,說明EcAQP11在脊尾白蝦滲透壓調(diào)節(jié)過程中發(fā)揮重要作用,是脊尾白蝦耐堿機制中的重要基因。

圖6 EcAQP11基因氨基酸序列與其他已知物種同源序列比對
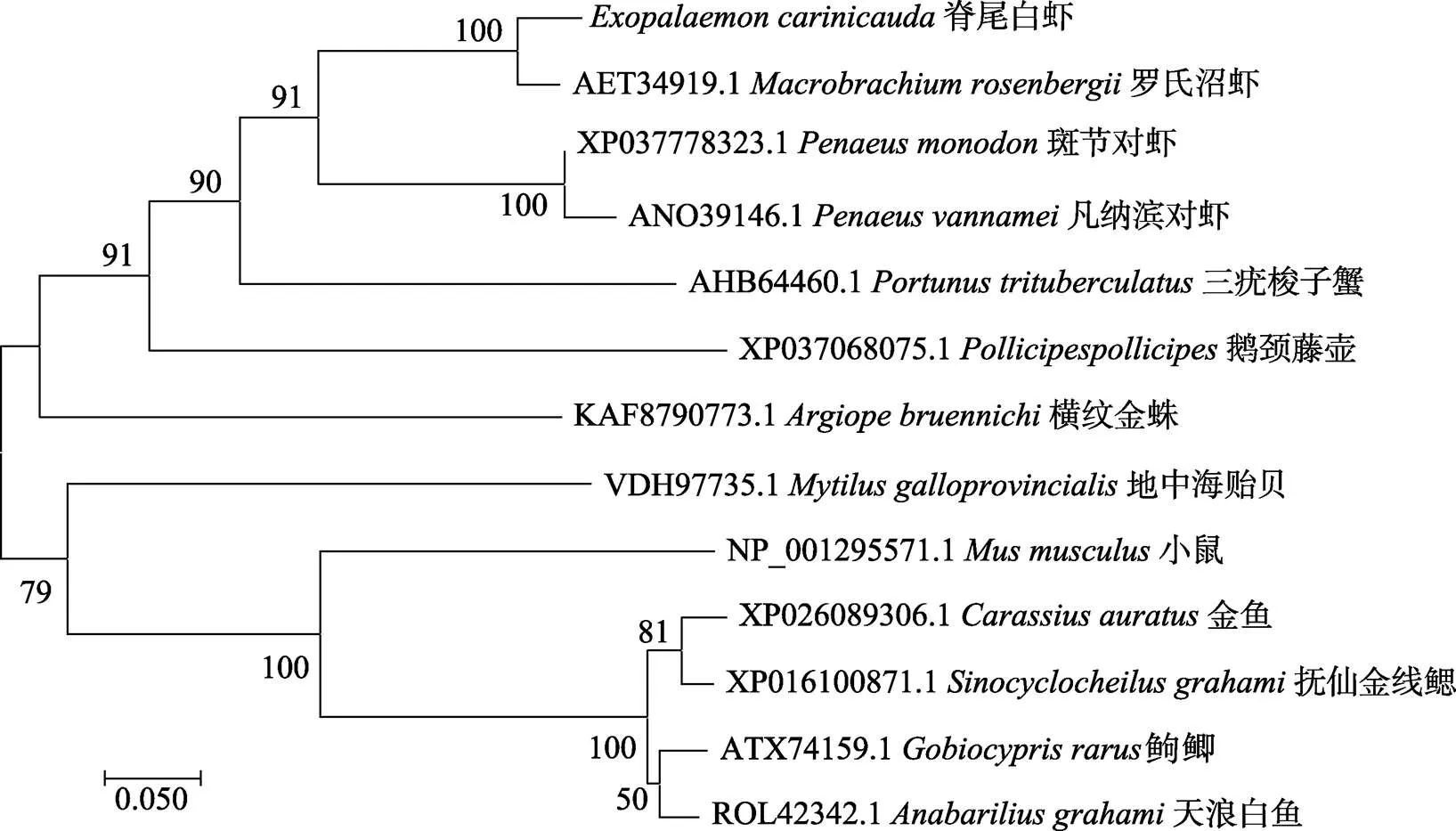
圖7 基于EcAQP4基因氨基酸序列構(gòu)建的Neighbor-joining進化樹

圖8 基于EcAQP11基因氨基酸序列構(gòu)建的Neighbor-joining進化樹

圖9 脊尾白蝦注射干擾后EcAQP4 (A)與EcAQP11 (B)基因的表達情況
GFP為對照組。不同字母表示差異顯著(<0.05),下同。
GFP was the control group. Different letters indicate significant difference (<0.05). The same as below.

圖10 RNA干擾后脊尾白蝦的死亡率
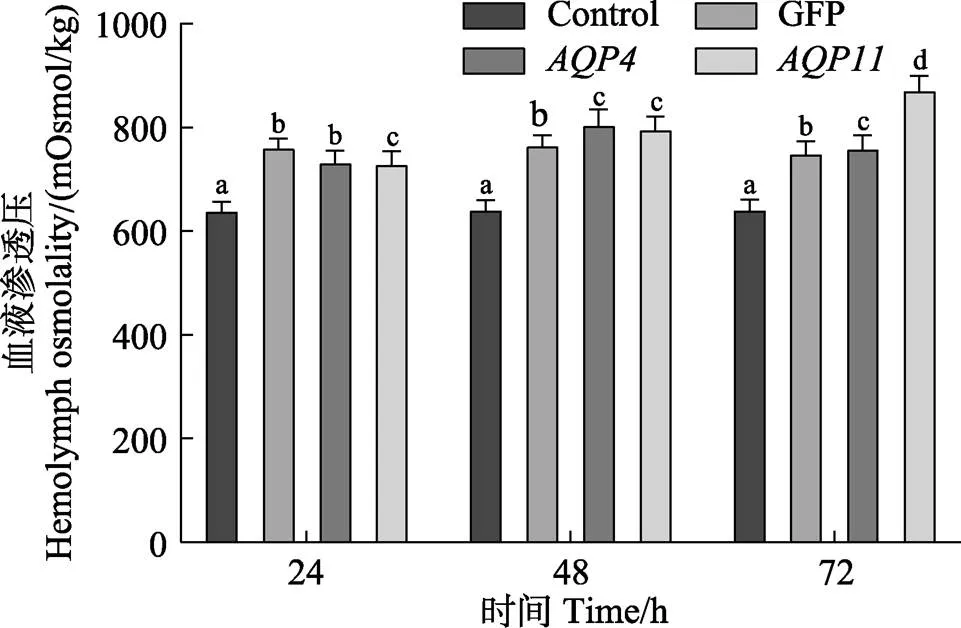
圖11 RNA干擾后脊尾白蝦的血液滲透壓變化
3 討論
本研究通過對轉(zhuǎn)錄組中差異表達基因的分析,發(fā)現(xiàn)在堿度脅迫后,與滲透壓調(diào)節(jié)相關(guān)的基因大量表達,為了進一步了解脊尾白蝦的耐堿機制,我們從差異表達基因中挑選了水通道蛋白這一滲透壓調(diào)節(jié)的關(guān)鍵基因(Deane, 2006; Holm, 2005; MacIver, 2009; Preston, 1992; Robinson, 1996; 隋海心等, 2004),通過RACE技術(shù)克隆cDNA全長并對其進行序列分析,利用RNA干擾技術(shù)驗證其在堿度脅迫過程中的響應(yīng)機制。
3.1 AQP4與AQP11基因的氨基酸序列及同源性分析
本研究利用RACE技術(shù)首次克隆獲得脊尾白蝦與基因的cDNA全長,并運用生物信息學(xué)軟件對其序列進行分析。通過比對發(fā)現(xiàn),脊尾白蝦基因保守性較好,具有HINPAVT和PLAIGL兩個保守域,與高沿等(2017)研究結(jié)果相同。
3.2 EcAQP4與EcAQP11基因的功能驗證
水通道蛋白是生物細胞膜轉(zhuǎn)運水分子的重要結(jié)構(gòu),在維持細胞內(nèi)外滲透壓平衡中起重要作用,現(xiàn)階段關(guān)于水通道蛋白的研究有很多,最初人們在腦星形膠質(zhì)細胞、胃壁細胞和腎集合小管等部位發(fā)現(xiàn)AQP4蛋白,它是一種水特異性通道蛋白,僅對水分子具有滲透性,其在人體中主要作用為維持腦內(nèi)水平衡及腦水腫發(fā)生發(fā)展(張士保等, 2013)。現(xiàn)階段對AQP4的研究主要集中于其在腦水腫、視神經(jīng)脊髓炎(Yang, 2013)、癲癇(Hsu, 2011)等中樞神經(jīng)系統(tǒng)疾病中所起的作用,其調(diào)節(jié)機制主要是維持細胞間隙K+緩沖與滲透壓平衡,但其在水產(chǎn)動物中的研究還未見報道。基因在各個物種之間的氨基酸同源性低,功能尚不確定,但有研究表明,具有水轉(zhuǎn)運活性,作為水通道發(fā)揮作用(Yakata, 2007),對體內(nèi)水的穩(wěn)態(tài)平衡有重要作用(He, 2017),其發(fā)揮作用的主要方式為維持上皮細胞水和離子的吸收轉(zhuǎn)運,保持細胞滲透壓平衡。
為驗證與兩基因在脊尾白蝦適應(yīng)堿度脅迫中所起的作用,本研究設(shè)計合成了相應(yīng)的dsRNA,對脊尾白蝦進行注射干擾后,通過qRT-PCR技術(shù)檢測了堿度脅迫后上述2個基因的表達變化,結(jié)果顯示,與對照組相比,各個時間點基因的表達均發(fā)生了顯著下降,說明所設(shè)計的dsRNA具有良好的干擾效果。對脊尾白蝦注射干擾后進行堿度脅迫,發(fā)現(xiàn)其死亡率明顯升高,堿度脅迫72 h時,組死亡率達到45%,組達到55%,說明基因的沉默對脊尾白蝦的耐堿度能力產(chǎn)生明顯影響。滲透壓測定結(jié)果顯示,與空白組相比,GFP對照組血液滲透壓升高,各時間點總體變化不大,從側(cè)面反映了脊尾白蝦的耐堿能力較強。滲透壓的變化趨勢總體來說呈現(xiàn)先上升后下降的趨勢,與鹽堿脅迫對尼羅羅非魚()血清滲透壓變化趨勢相同(趙麗慧等, 2014)。在堿度脅迫條件下,脊尾白蝦體內(nèi)的離子逐漸累積增強,導(dǎo)致血液滲透壓增高,但脊尾白蝦通過體內(nèi)的滲透壓調(diào)節(jié)機制將滲透壓維持穩(wěn)定。與GFP對照組相比,干擾組脊尾白蝦的血液滲透壓上升下降的幅度變大,說明基因的沉默表達影響了脊尾白蝦的滲透壓調(diào)節(jié)能力,從而導(dǎo)致脊尾白蝦耐堿度能力減弱,死亡率升高。相比對照組和干擾組,干擾組的滲透壓升高更明顯,且呈現(xiàn)一直升高的趨勢,這可能是基因在脊尾白蝦耐堿機制中起到了更重要的作用,其對脊尾白蝦的滲透壓調(diào)節(jié)影響較大。前文提到,基因現(xiàn)階段的功能與調(diào)控機制尚未完全解析,但本研究結(jié)果表明,基因在脊尾白蝦滲透壓調(diào)節(jié)中發(fā)揮了重要作用。
本研究成功克隆了脊尾白蝦與基因,設(shè)計干擾dsRNA并對脊尾白蝦進行注射沉默上述2個基因的表達,結(jié)果發(fā)現(xiàn),與對照組相比,注射干擾后的脊尾白蝦在堿度脅迫下死亡率明顯升高;干擾組脊尾白蝦血液滲透壓變化幅度增大,干擾組血液滲透壓升高明顯,干擾與表達后的脊尾白蝦耐堿度脅迫能力減弱,表明上述2個基因在脊尾白蝦適應(yīng)堿度脅迫過程中發(fā)揮著重要作用。
DEANE E E, WOO N Y S. Tissue distribution, effects of salinity acclimation, and ontogeny of aquaporin 3 in the marine teleost, silver sea bream (). Marine Biotechnology, 2006, 8(6): 663–671
DUAN M S. Identification of aquaporin gene in tea tree and study offunctional. Master′s Thesis of Northwest A&F University, 2020 [段夢莎. 茶樹水通道蛋白基因的鑒定與的功能研究. 西北農(nóng)林科技大學(xué)碩士研究生學(xué)位論文, 2020]
FENG X C, MA T H. Physiological importance of aquaporin water channels accessed by phenotype studies of aquaporin knockout mice. Progress in Biochemistry and Biophysics, 2005, 32(4): 291–297 [馮學(xué)超, 麻彤輝. 水通道蛋白的生理功能——水通道基因敲除小鼠表型研究進展. 生物化學(xué)與生物物理進展, 2005, 32(4): 291–297]
GAN Y D. cDNA cloning of aquaporin 3 in,and tissue expression patterns under salinity stresses.Master′s Thesis of Shanghai Ocean University, 2014[甘遠迪. 薩羅羅非魚、尼羅羅非魚AQP3cDNA序列克隆及鹽度脅迫下組織表達特征. 上海海洋大學(xué)碩士研究生學(xué)位論文, 2014]
GAO Y, HU C Q, REN C H,. Molecular cloning of aquaporin-4 () gene in the Pacific white shrimp () and the effect of salinity stress on its expression in hepatopancreas. Marine Sciences, 2017, 41(2): 61–70[高沿, 胡超群, 任春華, 等. 凡納濱對蝦水孔蛋白-4的cDNA克隆及鹽度脅迫對其肝胰腺mRNA表達水平的影響. 海洋科學(xué), 2017, 41(2): 61–70]
HAO J F. Genome-wide identification of aquaporin gene family in cotton and functional characterization in response to osmotic stresses. Master′s Thesis of Zhengzhou University, 2020 [郝建峰. 棉花水通道蛋白家族基因鑒定及其滲透脅迫應(yīng)答功能研究. 鄭州大學(xué)碩士研究生學(xué)位論文, 2020]
HE L Q, HUANG N, LI H,. AMPK/α-ketoglutarate axis regulates intestinal water and ion homeostasis in young pigs. Journal of Agricultural and Food Chemistry, 2017, 65(11): 2287–2298
HOLM L M, JAHN T P, M?LLER A L B,.NH3and NH4+permeability in aquaporin-expressingoocytes. Pflügers Archiv-European Journal of Physiology, 2005, 450(6): 415–428
HSU M S, SELDIN M, LEE D J,Laminar-specific and developmental expression of aquaporin-4 in the mouse hippocampus. Neuroscience, 2011, 178: 21–32
LI J, WANG L, LUO X L,Cloning of AQP1 and AQP3 genes and their locations and expressions in different tissues of yak (). Acta Theriologica Sinica, 2021, 41(1): 51–58 [李娟, 王利, 羅曉林, 等. 牦牛水通道蛋白AQP1和AQP3基因克隆及在不同組織中表達和定位. 獸類學(xué)報, 2021, 41(1): 51–58]
LI Q Y. Genome-wide identification and transcriptome analysis of aquaporin gene family inMaster′s Thesis of Shandong Agricultural University, 2020 [李青云. 普通煙草中水通道蛋白基因家族的鑒定和轉(zhuǎn)錄組分析. 山東農(nóng)業(yè)大學(xué)碩士研究生學(xué)位論文, 2020]
LI W X, ZHANG D Y, ZHU G Z,. Combining genome-wide and transcriptome-wide analyses reveal the evolutionary conservation and functional diversity of aquaporins in cotton. BMC Genomics, 2019, 20(1): 538
MACIVER B, CUTLER C P, YIN J,Expression and functional characterization of four aquaporin water channels from the European eel (). Journal of Experimental Biology 2009, 212(17): 2856–2863
PRESTON G M, CARROLL T P, GUGGINO W B,Appearance of water channels inoocytes expressing red cell CHIP28 protein. Science, 1992, 256(5055): 385–387
ROBINSON D G, SIEBER H, KAMMERLOHER W,. PIP1 aquaporins are concentrated in plasmalemmasomes of arabidopsis thaliana mesophyll. Plant Physiology, 1996, 111(2): 645–649
SUI H X, REN G. Structure and mechanism of water channels. Progress in Chemistry, 2004, 16: 145 [隋海心, 任罡. 水分子通道蛋白的結(jié)構(gòu)與功能. 化學(xué)進展, 2004, 16(2): 145–152]
WANG Y, Lü J J, LIU P,. Cloning and characterization of aquaporins 1 and its expression analysis under salinity stress inJournal of Fishery Sciences of China, 2014, 21(5): 893–901 [王渝, 呂建建, 劉萍, 等. 三疣梭子蟹水通道蛋白1 cDNA及其鹽度脅迫下的表達分析. 中國水產(chǎn)科學(xué), 2014, 21(5): 893–901]
YAKATA K, HIROAKI Y, ISHIBASHI K,. Aquaporin-11 containing a divergent NPA motif has normal water channel activity. Biochimica et Biophysica Acta, 2007, 1768
YANG Y, HUANG D H, WU W P,The role of aquaporin-4 antibodies in Chinese patients with neuromyelitis optica. Journal of Clinical Neuroscience, 2013, 20(1): 94–98
YANG Z G, ZHANG L, CHEN C Y,. Study on cloning, expression and RNA interference of aquaporin 1 gene of. Journal of Fudan University (Natural Science), 2021, 60(1): 93–101 [楊志剛, 張龍, 陳春宇, 等. 中華絨螯蟹水通道蛋白1基因的克隆、表達及RNA干擾研究. 復(fù)旦學(xué)報(自然科學(xué)版), 2021, 60(1): 93–101]
ZHAO L H, ZHAO L, JEERAWAT T,. Effects of salinity-alkalinity on serum osmolality, ion concentration and mRNA expression of ion transport enzymes of. Journal of Fisheries of China, 2014, 38(10): 1696–1704 [趙麗慧, 趙金良, Jeerawat T, 等. 鹽堿脅迫對尼羅羅非魚血清滲透壓、離子濃度及離子轉(zhuǎn)運酶基因表達的影響. 水產(chǎn)學(xué)報, 2014, 38(10): 1696–1704]
ZHANG S B, DAI D W, DAI Y M. Advances of aquaporin-4 research. Chinese Journal of Clinical Neurosciences, 2013, 21(4): 451–455 [張士保, 代大偉, 代亞美. 水通道蛋白-4的研究進展. 中國臨床神經(jīng)科學(xué), 2013, 21(4): 451–455]
ZHANG Y N, WANG Y, ZHU J J,. Analysis of cloning and expression of AQP7 gene in Tibetan goat. Southwest China Journal of Agricultural Sciences, 2019, 32(8): 1956–1960 [張亞楠, 王永, 朱江江, 等. 藏山羊AQP7基因克隆和不同組織器官差異表達分析. 西南農(nóng)業(yè)學(xué)報, 2019, 32(8): 1956–1960]
The Roles of Aquaporin Gene 4 and 11 ofunder Alkalinity Stress
LI Mingdong1,2, WANG Jiajia1,2, GE Qianqian1,2, QIN Zhen1,2, LIU Ping1,2, LI Jian1,2, LI Jitao1,2①
(1. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Qingdao, Shandong 266071, China;2. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao, Shandong 266071, China)
Aquaporins are a family of cell membrane proteins, and their key role is to specifically transport water molecules and other neutral metabolic molecules. Moreover, aquaporins play an important role in regulating the balance between internal and external osmotic pressure in organisms. Here, aquaporin 4 and 11 genes fromwere successfully cloned using rapid amplification of cDNA ends cloning. The open reading frame ofis 621 bp, encoding 206 amino acids, with a predicted molecular weight of 21.673 kDa and a theoretical isoelectric point of 8.30; it is a hydrophobic protein with five transmembrane structural domains. The open reading frame ofis 783 bp, encoding 260 amino acids, with a predicted molecular weight of 28.490 kDa and a theoretical isoelectric point of 5.40; it is a hydrophobic protein with four transmembrane domains. In sequence alignment, AQP4 inshared the highest homology with that in(94.63%), while AQP11 inshared the highest homology with that in(81.47%).Furthermore, RNA interference was used for silencingandexpression to verify their function. In carbonate alkalinity stress, the mortality ofincreased significantly following RNA interference. At 72 h, mortality reached 45% and 55% in theandgroups, respectively, being significantly higher than that in the control group. The blood osmotic pressure ofin thegroup was significantly higher than that in the control group. Similarly, the blood osmotic pressure ofin thegroup was significantly increased. In summary, aquaporins play important roles in regulating osmotic pressure and maintaining ion balance in response to alkalinity stress in.
; Aquaporin;Gene cloning; Osmolality
LI Jitao, E-mail: lijt@ysfri.ac.cn
10.19663/j.issn2095-9869.20211231003
S917
A
2095-9869(2022)04-0051-10
*國家重點研發(fā)計劃課題(2018YFD0901302)、國家自然科學(xué)基金(32072974)、財政部和農(nóng)業(yè)農(nóng)村部: 國家現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系和中國水產(chǎn)科學(xué)研究院基本科研業(yè)務(wù)費項目(2020TD46)共同資助 [This study was supported by National Key Research and Development Program of China (2018YFD0901302), National Natural Science Foundation of China (32072974), China Agriculture Research System of MOF and MARA, and Central Public-Interest Scientific Institution Basal Research Fund, CAFS (2020TD46)]. 李明棟,E-mail: lmdanyany@163.com
李吉濤,研究員,E-mail: lijt@ysfri.ac.cn
2021-12-31,
2022-02-11
http://www.yykxjz.cn/
李明棟, 王佳佳, 葛倩倩, 秦楨, 劉萍, 李健, 李吉濤. 脊尾白蝦水通道蛋白基因4和11在堿度脅迫過程中的作用. 漁業(yè)科學(xué)進展, 2022, 43(4): 51–60
LI M D, WANG J J, GE Q Q, QIN Z, LIU P, LI J, LI J T. The roles of aquaporin gene 4 and 11 ofunder alkalinity stress. Progress in Fishery Sciences, 2022, 43(4): 51–60
(編輯 馮小花)
