三種厚樸葉綠體基因組的比較研究
2022-09-07 09:35:34尹彥棚周羅靜時小東侯飛俠高繼海
廣西植物 2022年8期
張 敏, 尹彥棚, 周羅靜, 任 波, 王 麗, 時小東, 侯飛俠, 彭 成, 高繼海*
( 1. 成都中醫藥大學 藥學院 西南特色中藥資源國家重點實驗室, 成都 611137; 2. 四川省林業科學研究院, 成都 610065; 3. 成都大學, 成都 610106 )
厚樸()、凹葉厚樸(subsp.)及日本厚樸()均為木蘭科藥材兩用經濟林闊葉樹種,廣泛分布于韓國、日本和中國等地區,具有水土保持、綠化、美化環境和藥用等多種應用價值(彭夢婕,2020)。目前三種厚樸的相關研究主要集中在化學成分、臨床應用及資源調查等方面(胥愛麗等,2021)。然而,日本厚樸非我國原產資源,生長于相對低溫環境中,具備許多優于另兩者的生物學性狀,如其耐寒能力相比于其他木蘭屬植物更強(Kwon & Oh, 2015),生長速度和成熟率都比同類植物更快,尤其是在生長幼期,以每年60~90 cm的速度成長(Oguchi et al., 2017),是解析和培育厚樸類物種優良性狀的理想材料。然而,日本厚樸的分子遺傳學研究資料匱乏,且三種厚樸的生長速度和抗寒能力差異的形成原因缺乏分子生物學機制研究,進而影響了厚樸類優良品種的選育。
葉綠體是植物細胞中必不可少的細胞器,在光合作用、固碳以及氨基酸的合成中起到重要作用,其全基因組包含大量遺傳信息,結構高度保守,被廣泛應用于植物分子進化及系統發育研究,并在藥用植物的遺傳轉化、基因工程和分子選育等方面發揮著重要作用(趙祺等,2021)。目前,厚樸和凹葉厚樸已有葉綠體基因組報道,而具有更多生物特性的日本厚樸卻缺乏葉綠體基因組的研究。鑒于此,本研究利用Illumina HiSeq高通量測序技術,對日本厚樸進行了葉綠體基因組測序,與既有的厚樸、凹葉厚樸葉綠體基因組比對,篩選出了諸多差異的核苷酸序列,破譯三種厚樸之間的親緣關系,找出三種厚樸具有更強生長發育能力和耐寒性等功能的相關基因,在一定程度上為以更強的生長速度和抗寒能力為優良性狀的目標物種提供了依據。
1 材料與方法
1.1 材料及預處理
日本厚樸新鮮、幼嫩、健康的葉片于2019年10月10日采自四川省成都市溫江區成都中醫藥大學藥用植物園(30°42′ E,103°49′ N),經國家中藥種質資源庫專家高繼海副教授鑒定所有樣品均來源于木蘭科木蘭亞屬日本厚樸()。三種厚樸的命名參考于中國植物物種信息數據庫(http://db.kib.ac.cn/)。葉片使用無菌水擦拭干凈,迅速凍存備用。憑證標本(館藏序列號為ZYC190910)保存于成都中醫藥大學中醫藥傳統文化博物館。
葉綠體提取步驟如下:新鮮葉片于液氮中研磨后懸于一定量A液(50 mmol·LTris,25 mmol·LEDTA,1.25 mol·LNaCl,0.25 mmol·LVc,1.5% PVP,pH 3.6)中,4層紗布過濾,4 ℃下200離心20 min,取上清,加入常溫緩沖液B(50 mmol·LTris,25 mmol·LEDTA,1.25 mol·LNaCl,0.25 mmol·LVc,1 mmol·LDTT,0.1%牛血清蛋白BSA,pH 8.0),常溫靜置,4 ℃下2 000離心10 min,棄上清,葉綠體沉淀于4 ℃保存備用。
1.2 葉綠體基因組提取和測序
針對葉綠體材料,采用改良的CTAB法(Matthes et al., 2020)分離提取DNA。DNA經檢測合格后,先用超聲波機械打斷,再進行片段純化、末端修復、3′端加A、連接測序接頭,進行PCR擴增形成測序文庫,最后使用高通量測序平臺Illumina HiSeq PE150 進行測序。測序得到的原始測序序列(Raw Reads),里面含有帶接頭的、低質量的Reads,為了保證信息分析質量,對Raw Reads進行過濾、質控,得到Clean Reads,用于后續信息分析。數據過濾的主要步驟如下:(1)去除帶接頭的Reads;(2)過濾N含量超過10%的Reads;(3)去除質量值低于10的堿基超過50%的Reads。對過濾后的高質量數據隨機抽取2 000條 Reads數據,通過BLAST軟件比對NT庫檢測樣品是否受到污染。
1.3 葉綠體基因組的組裝和注釋
原始序列上傳于國家生物信息中心數據庫(序列號PRJCA004348)。先利用Galaxy在線平臺(https://usegalaxy.org)對日本厚樸葉綠體基因組測序結果進行組裝(Yan et al., 2015),且下載厚樸(., NC_020316)和凹葉厚樸(subsp, JN867581)的葉綠體基因組FASTA文件,合并后作為日本厚樸的參考基因文件。再通過CPGAVAS2在線平臺(http://www.herbalgenomics.org/cpgavas2)完成厚樸、凹葉厚樸、日本厚樸葉綠體基因組的注釋。
1.4 聚類分析
除厚樸、凹葉厚樸外,又于NCBI數據庫中下載荷花玉蘭(, JN867584)、星花玉蘭(, NC_039941)、望春玉蘭(, KY085894)、武當玉蘭(, JX280401)、玉燈玉蘭(‘Lamp’, JN227740)、寶華玉蘭(, MH607378)、紫玉蘭(, JX280397)、天女木蘭(, NC_041435)、云南擬單性木蘭(, KF753638)、鵝掌楸(, NC_030504)、北美鵝掌楸(, NC_008326)共11種木蘭科植物的葉綠體基因組數據,其中包括2種鵝掌楸屬植物和其他9種木蘭科植物。利用MEGA X軟件,基于鄰接法(NJ法)構建日本厚樸在內的14種木蘭科植物的系統發育樹,觀察并分析它們之間的親緣關系(Yan et al., 2015)。
2 結果與分析
2.1 葉綠體基因組信息
通過Illumina HiSeq高通量測序平臺測序共獲得日本厚樸葉綠體19 816 708 條原始數據,移除接頭和低質量的Reads,共獲得Clean Reads 19 791 019條,Q30為91.33%。通過在線組裝發現日本厚樸全長160 051 bp,符合目前已知的木蘭類植物葉綠體基因組大小范圍(159 429~160 183 bp),厚樸葉綠體擁有目前已知木蘭類植物的最大基因組(160 183 bp),而凹葉厚樸的葉綠體基因組為160 099 bp。
三種厚樸葉綠體基因均具有典型的四分區域結構,其中LSC分別為88 210 bp(厚樸)、88 145 bp(凹葉厚樸)和 88 156 bp(日本厚樸),SSC分別為18 843、18 832和18 771 bp,2段反向互補重復的IR區(IRA和IRB)分別為26 565、26 566和26 562 bp。三種厚樸IR、LSC和SSC區域的GC值存在一定的差異,其中GC含量最高的區域都為IR區,分別為43.2%、43.1%和43.2%,LSC區次之,均為37.9%,而SSC區域的GC值最小,僅分別為34.2%、34.3%和34.3%。與其他物種葉綠體基因組相似的是,三種厚樸的1也跨越了SSC和IRA區,其中位于IRA區域長度均為1 279 bp,厚樸、凹葉厚樸位于SSC區域長度為4 311 bp,比日本厚樸長51 bp。除此之外,日本厚樸特有的1基因還跨越了SSC和IRB區,其序列長度分別為29、1 279 bp。
通過基因組結構比較,發現三種厚樸葉綠體的rRNA和tRNA數量相同(表1)。在基因數量方面,凹葉厚樸相比厚樸多2個單拷貝基因,而兩者又較日本厚樸分別少了4個和6個。在編碼區數量方面,凹葉厚樸與厚樸相比多2個編碼區,而日本厚樸數量最多,比前兩者分別多8個和10個。這揭示了厚樸與凹葉厚樸的葉綠體基因組可能存在較小的結構和功能差異,而日本厚樸的葉綠體基因組結構和功能可能較其他兩者更為豐富。
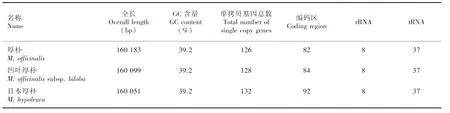
表 1 三種厚樸葉綠體基因基本信息Table 1 Basic chloroplast genes information of three Magnolia species
2.2 葉綠體內含子、外顯子比較
對三種厚樸的葉綠體基因組注釋文件進行分析,發現在近百個木蘭類葉綠體基因組編碼基因中,19個存在內含子、外顯子的差異而3個基因的內含子、外顯子數量和長度完全一致(表2)。在這19個差異片段中,厚樸與凹葉厚樸的差異主要表現在11個,其中9個片段的差異區在I類內含子(In I,核苷酸數量差異在20以內),2個在Ⅱ類內含子(InⅡ,核苷酸數量差異在2以內),還有1個差異片段存在Ⅱ類外顯子(2的EPⅡ,40 bp,此區域也為厚樸、凹葉厚樸葉綠體基因組差異最大的區域)。相較于前兩個厚樸物種,日本厚樸存在更多差異,尤其是-基因的I類內含子比前兩者多920個核苷酸,甚至還多了三種全表達的功能基因(22、1-和15,)(表2),這些結果再次顯示厚樸與凹葉厚樸的葉綠體基因組之間的差異較小,而日本厚樸的葉綠體基因組在結構和功能方面較其他兩者更為豐富。
在三種厚樸葉綠體的2個12基因中,一個正常含有3個外顯子,而另一個缺失了I類外顯子,無法正常表達,可能為假基因。植物葉綠體基因普遍具有保守性與突變性(轉移或損失等)并存的現象,其轉錄涉及到復雜的反向剪接等過程,本研究發現三種厚樸中12的幾個外顯子為反向排列,且為非順序式。核糖體小亞基基因的假基因轉化在物種進化過程中起到重要作用(Liu et al., 2020),因此12核苷酸序列的差異可為厚樸類親緣關系研究提供重要參考。三種厚樸葉綠體差異最大的3基因,其差異區域數達4個(I類、Ⅱ類內含子和I類、Ⅲ類外顯子),日本厚樸的I類外顯子增加了102個核苷酸。此外,與木蘭類植物的葉綠體基因組條形碼-、等相似,本研究針對三種厚樸的比較分析發現-、16、1、、、2等基因的變異位點率、信息位點率較高,也具備開發為少數木蘭植物鑒定條形碼的潛力。

表 2 三種厚樸葉綠體基因組內含子、外顯子比較Table 2 Comparison of introns and exons of three Magnolia species chloroplast genomes
2.3 葉綠體功能基因比較


表 3 三種厚樸葉綠體的功能基因比較Table 3 Comparison of chloroplast functional genes of three Magnolia species
綜合本文內含子與外顯子、功能基因的結構分析結果, 三種厚樸葉綠體基因組的差異主要分布于LSC區和IR區,涉及核糖體大亞基、核糖體小亞基和未知功能基因類群,尤其是諸多未知功能的基因,而木蘭屬中這類疑似非功能基因的結構特征、變異原因,以及與適生環境差異的關聯性,有待進一步深入研究。
2.4 部分木蘭科植物的親緣關系
由圖1可知,在14種近緣物種葉綠體基因組構建的NJ發育樹(節點支持率均在75%以上)中,鵝掌楸屬和其他屬是木蘭科中的兩個獨立的譜系,這與之前報道的結果一致(陳凱,2019)。在其他屬的2個次級類群中,三種厚樸、天女木蘭、荷花玉蘭聚類在一起,其中日本厚樸與凹葉厚樸親緣關系最為密切,其次是厚樸。其他屬中另外一個次級類群包括云南擬單性木蘭、玉燈玉蘭、紫玉蘭、武當玉蘭、望春玉蘭、星花玉蘭、寶華玉蘭,它們與三種厚樸親緣關系由近及遠。

圖 1 部分木蘭科物種系統發育樹Fig. 1 Phylogenetic tree of partial Magnoliaceae plants
3 討論與結論
本研究使用全基因組測序來組裝和比較三種厚樸的葉綠體基因組,以此來推測三種厚樸之間多種生物學性狀差異的形成原因。低溫環境下,植物最先受到抑制的生理代謝過程是光合作用,低溫能顯著降低光合效率、CO同化作用和光系統Ⅱ活性,從而影響植物的正常生長發育(王璐等,2020)。通過三種厚樸葉綠體的功能基因比較,發現日本厚樸基因在數量上較另兩者增加了一個,這可能緣于基因的自我復制。和是光合系統Ⅱ的關鍵基因,例如小麥成熟葉綠體psbC蛋白的合成發生于含序列的轉錄本上,形成-復合物,它們的轉錄水平受到光誘導, 通過增強光合系統Ⅱ亞基的合成和維持能力,提高葉綠體的合成速度(Gamble et al., 1988; Gamble & Mullet, 1989),從而提高葉綠體的光合作用強度,進而加快植物的生長發育。此外,-基因協同轉錄的mRNAs可以翻譯產生D2和CP43蛋白,與反應中心D1、CP47、放氧復合體蛋白及捕光復合體Ⅱ等蛋白,共同參與葉綠體光合系統Ⅱ的光合電子傳遞,在強光條件下維持光合系統Ⅱ的功能(莊焜揚,2020)。-復合體直接參與光合系統Ⅱ亞基的形成,在生長旺盛和存在顏色差異的葉片中發揮著保護光合系統、減少強光傷害的作用(Adachi et al., 2012),這在花葉矢竹葉等光合系統Ⅱ中基因的研究中也得到證實(許冰清等,2015)。因此,日本厚樸葉綠體增加的基因表明其具備合成更多-復合體與D2蛋白的潛力,這可能是日本厚樸生長速度更快、適應較北緯度地區低溫和強光脅迫環境的原因之一。
在高變異位點率的功能基因中,22為日本厚樸中特有,其位于LSC區,屬于基因表達相關功能基因,可用于物種鑒定(Feng et al., 2019)。此外,本研究發現三種厚樸的差異基因主要分布于22所屬的基因簇中,它們組成23操縱子的大轉錄單元,翻譯為核糖體大蛋白亞基。本研究發現,此操縱子在三種厚樸中存在顯著的結構差異:23-2-19-22(日本厚樸)-3(凹葉厚樸、日本厚樸)-16-14-8--36-11-。通過三個物種的親緣關系可知,凹葉厚樸、日本厚樸與厚樸分化的過程中,該操縱子19-16之間原有的核苷酸空隙逐次被3、22基因填充,同時造成操縱子中多個功能基因的內含子、外顯子核苷酸發生了增減(減少為主),功能蛋白的數量和活性也發生了改變。植物葉綠體除了光合作用外,還部分參與氨基酸、核苷酸、脂類和淀粉等各類成分的生物合成,支撐起該植物的生物學性狀(Namgung et al., 2021),日本厚樸具備更豐富的核糖體基因簇,這無疑形成了其更豐富的生物學特性。
tRNA作為核酸信息水平和蛋白質功能水平的適配器,在蛋白質翻譯中起著核心作用,其結構修飾影響植物體的溫度適應能力(Lorenz et al., 2017)。本研究發現日本厚樸tRNA-ALA比其他兩種厚樸多3個,具有快速轉運和積累丙氨酸的潛力,而游離態的丙氨酸能抵抗多種外界環境刺激(Mustroph et al., 2014),如寒冷因子。除了數量因素外,三種厚樸葉綠體中tRNA對應的氨基酸種類也不同。日本厚樸為本屬植物中分布較為北端(如千島群島)的物種,其生境中弱光和低溫信號交叉影響著植物的生長發育,在這種光溫脅迫條件下,tRNA可能一方面積累更多的游離丙氨酸,另一方面降低正常蛋白質翻譯速率,將正常蛋白質合成轉向應激反應蛋白質的合成(馮德江等,2002),最終使得植株具備更強的耐寒性,這又為厚樸優良品種的分子篩選和培育提出了參考。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
