玉米/大豆間作種植對農田大型土壤動物群落動態變化的影響
2022-09-12 06:36:36趙巴音那木拉德海山劉鵬飛常國軍
中國土壤與肥料 2022年7期
趙巴音那木拉,德海山,紅 梅*,劉鵬飛,興 安,葉 賀,常國軍,美 麗
(1.內蒙古農業大學草原與資源環境學院,內蒙古自治區土壤質量與養分資源重點實驗室,內蒙古 呼和浩特 010011;2.赤峰市巴林左旗委巡察辦,內蒙古 赤峰 025450;3.內蒙古沿河流域生態產業有限公司,內蒙古 鄂爾多斯 017000)
我國面臨農業資源、環境和糧食等問題的諸多挑戰,間作種植模式再次引起關注[1]。豆科-禾本科間作種植模式歷史悠久[2],我國每年種植面積超過2800萬hm2[3]。豆科-禾本科間作作為一種傳統的種植模式,能夠充分利用水、肥、光和熱等自然資源,實現了農田土地在時間和空間上的集約化利用[4],提高自然資源利用效率[5-6],有效減少病蟲草害的發生,減少農藥的施用[7],有利于保護農田環境質量[8];同時利用豆科作物的共生固氮作用,將固定的氮向禾本科作物轉移,促進禾本科作物對氮素的吸收[9]。在自然界中物質循環的原動力主要是靠土壤生物完成,它直接或間接參與有機物的分解和營養元素的礦化,已有研究證明原生動物、線蟲、螨類、跳蟲、蚯蚓和一些大型節肢動物參與氮循環[10],土壤動物是土壤生態系統的重要組成部分及物質循環的重要參與者[11],調節土壤養分礦化等物質循環的過程[12],并在維持和發揮農田生態系統正常功能上起著不可替代的作用[13]。
關于玉米/大豆間作種植模式對大型土壤動物群落結構變化的影響研究報道較少,為此,本研究通過玉米/大豆間作和玉米單作2種種植模式,探討玉米/大豆間作種植模式對大型土壤動物群落的響應特征,為間作種植模式下土壤動物群落動態變化提供科學評價指標和依據。
1 材料與方法
1.1 試驗地概況
本研究在內蒙古呼倫貝爾市阿榮旗樂昌五組(48°20′N,123°31′E)進行,位于大興安嶺向松嫩平原過渡的黑土帶,海拔平均204 m。半干旱、半濕潤溫帶大陸性氣候,雨熱同季,年均氣溫1.7℃,年有效積溫2394℃,年均降水量約450 mm,降水主要集中在6~8月。無霜期90~130 d。土壤類型為黑鈣土,0~20 cm土層土壤有機質38.24 g/kg,全氮1.94 g/kg,有效磷17.91 mg/kg、速效鉀148.27 mg/kg。
1.2 試驗設計
試驗開始的時間為2016年,采用統一常規耕作方法,隨機區組設計,3次重復。設置玉米單作(M)和玉米/大豆間作(M-S)2種種植方式,施肥統一為摻混肥料(N-P2O5-K2O=21-14-10),施用量為540 kg/hm2。灌水方式為井水噴灌,生育期灌水2次;玉米株距25 cm,行距60 cm,小區面積400 m2。每個小區四周有5壟作為保護行,小區之間間隔2 m。
樣品采集于2016年玉米和大豆生長季,即玉米出苗期、拔節期、抽穗期和成熟期,具體時間為6月14日、7月18日、8月15日和9月26日,分別于玉米單作種植和玉米/大豆間作種植小區的玉米行進行采樣。為了消除邊界效應,在每個小區典型玉米生長生境選擇3個取樣點,分別對0~10、10~20和20~30 cm共3個土層依次用鐵鍬沿土壤剖面取樣,每個樣點采樣面積為20 cm×20 cm=400 cm2,采用手揀法收集大型土壤動物(體長在2 mm以上的土壤動物)。將收集到的大型土壤動物裝在盛有75%酒精的塑料瓶中。分類鑒定參考《中國土壤動物檢索圖鑒》[14],并在顯微鏡(Olympus CKX41)下對標本進行鑒定,根據食性不同進一步歸類為植食性、捕食性、腐食性和雜食性大型土壤動物。
1.3 數據統計與分析
類群等級按個體數占總個體數比例進行劃分:10%以上為優勢類群,1%~10%為常見類群,1%以下的為稀有類群。大型土壤動物群落特征分析采用Shannon-Weiner多樣性指數(H′=-Σpilnpi,式中,Pi為第i個物種的個體數占總個體數的比率)、Pielou均勻度指數(E=H′/lnS,式中,H′為多樣性指數,S為類群數)、Margalef豐富度指數[D=(S-1)/lnN,式中,S為類群數,N為總個體數]和Simpson優勢度指數(C=Σpi2)進行。大型土壤動物功能群的分類參考文獻[15-16]進行。采用重復的方差分析方法,評價種植模式(M和M-S)、采樣時間(6、7、8和9月)及垂直分布(0~10、10~20和20~30 cm土層)對土壤大型動物群落多樣性指數和功能群的影響。采用LSD比較各種植模式、采樣時間和垂直分布之間差異顯著性分析,P<0.05為統計學中有顯著差異;數據分析處理與作圖采用Excel 2003、SAS 9.0進行。
2 結果與分析
2.1 大型土壤動物群落組成
研究區共捕獲大型土壤動物818只,分別隸屬于12目34個類群(表1)。其中優勢類群為象甲科幼蟲、隱翅甲科和線蚓科3類,占總個體數的68.95%;常見類群為葉甲科、象甲科、步甲科、跳甲科、蟻科、正蚓科和奧地蜈蚣科7類,占總個體數的22.49%;其余24類為稀有類群,占總個體數的8.56%;優勢類群構成了研究區大型土壤動物的主體。研究區玉米單作種植獲取大型土壤動物345只,占總研究區的42.18%;玉米間作種植獲取大型土壤動物473只,占總研究區的57.82%。
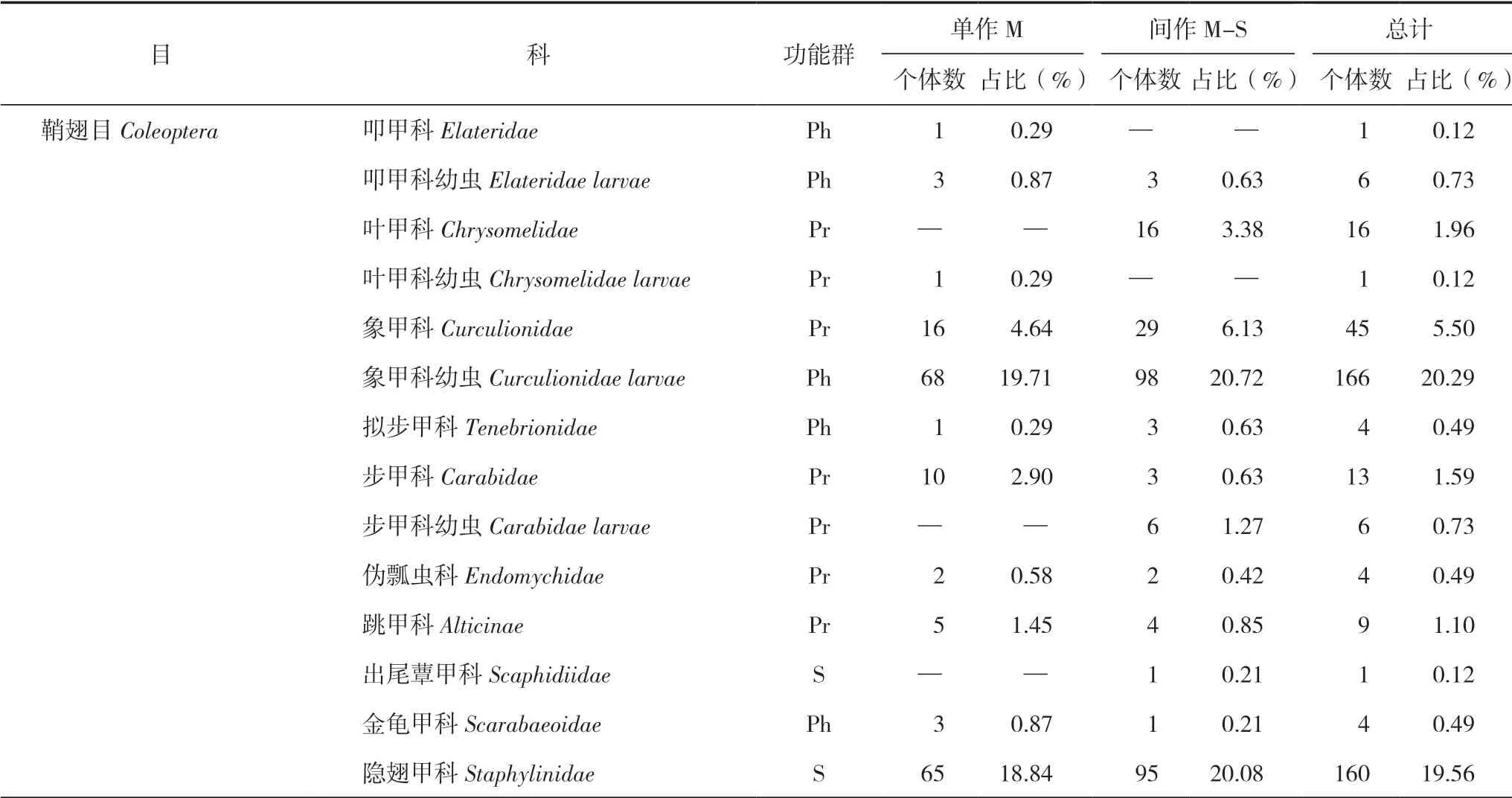
表1 大型土壤動物群落組成
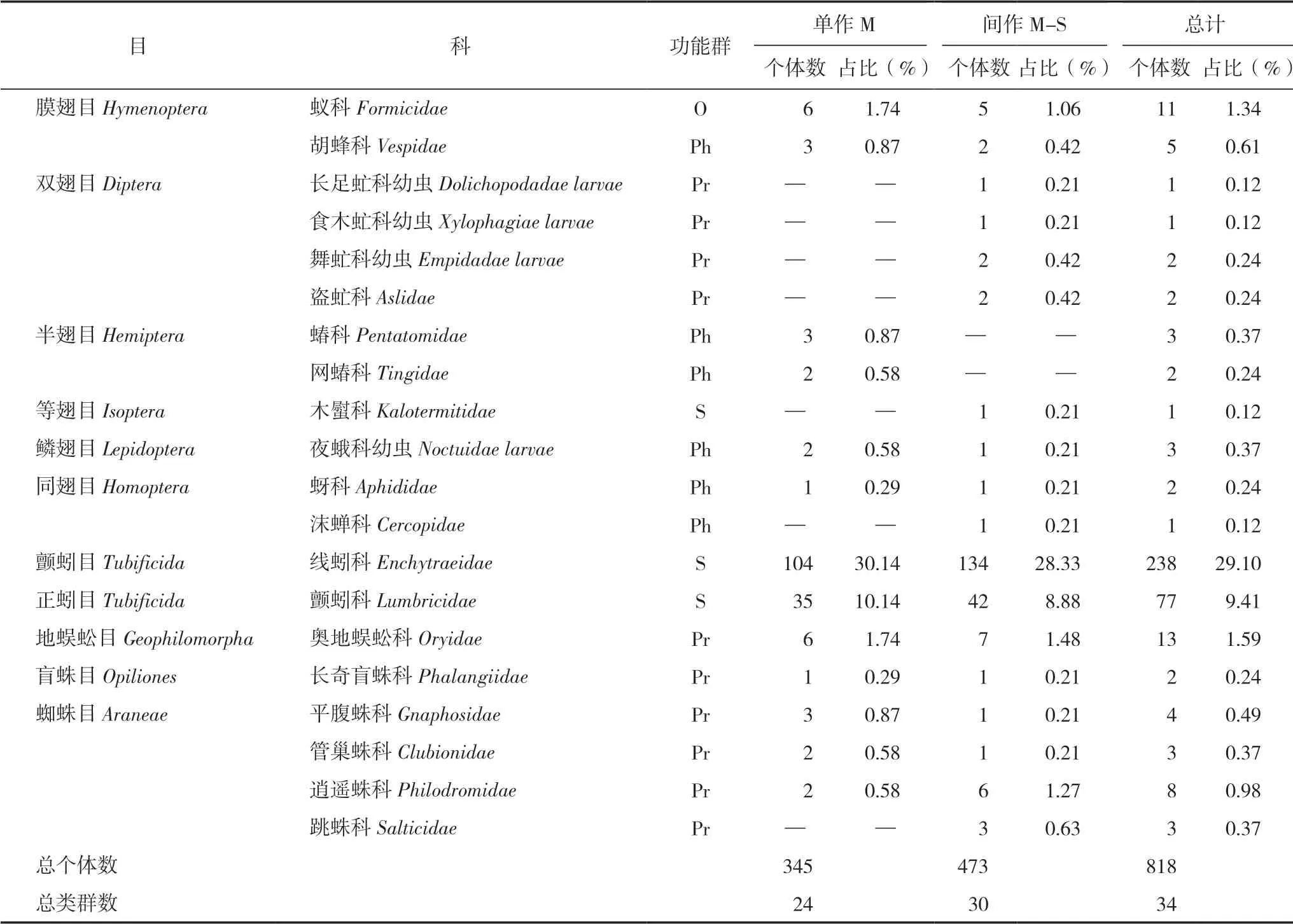
續表
2.2 大型土壤動物個體數和類群數動態特征
2.2.1 不同種植模式下大型土壤動物個體數和類群數動態變化
玉米單作和玉米/大豆間作種植下大型土壤動物個體數均隨玉米生長呈增加的趨勢,個體數表現為6月<7月和8月<9月(圖1a,P<0.05);玉米各生育時期類群數在月份間無顯著差異(圖1b,P>0.05)。7月玉米/大豆間作種植下個體數顯著高于玉米單作種植(P<0.05);玉米生育期內玉米/大豆間作種植和玉米單作種植下類群數均無顯著差異(P>0.05)。

圖1 土壤大型動物個體數和類群數的動態變化
2.2.2 大型土壤動物個體數和類群數垂直動態特征
不同月份0~10、10~20和20~30 cm 3個土層的個體數均在玉米各生育期內呈增加的趨勢,6月與9月中3個土層垂直分布的個體數有顯著變化(圖2a,P<0.05);7~9月大型土壤動物個體數均在10~20 cm土層最多,7~9月不同土層個體數變化中,10~20 cm > 0~10 cm 與20~30 cm 土層(圖2a,P<0.05)。
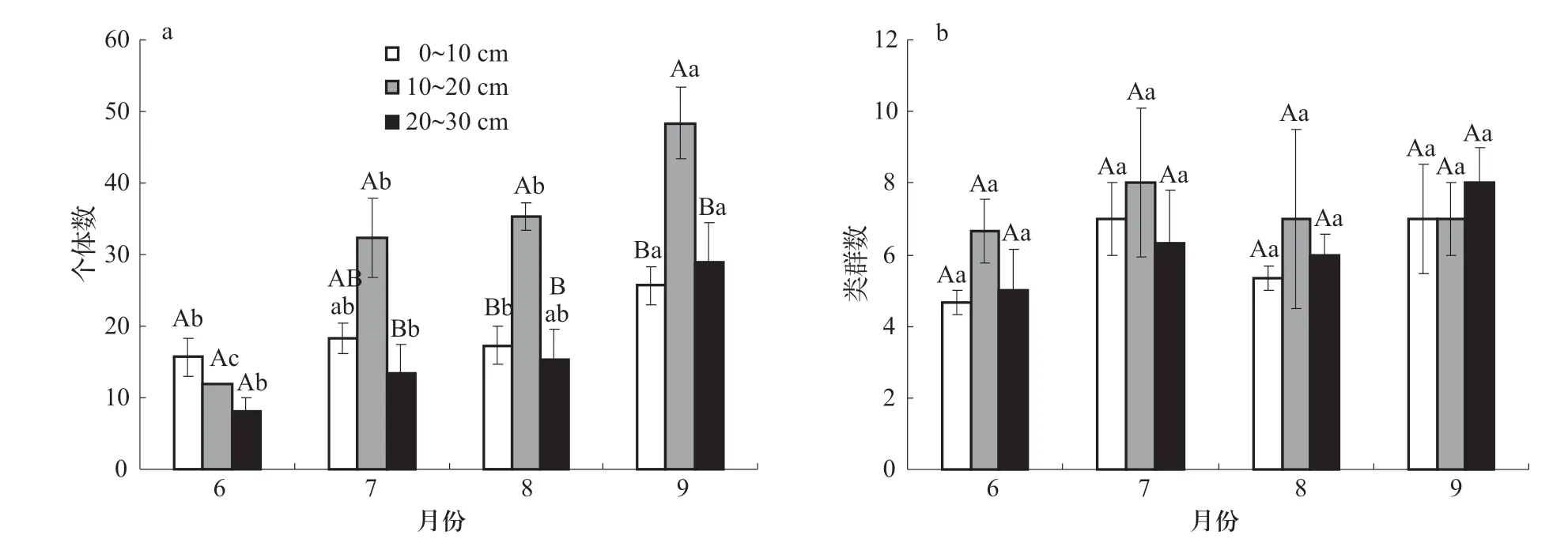
圖2 個體數和類群數垂直分布的動態特征
在玉米各生育期相同土層不同月份間大型土壤動物類群數均無顯著變化(P>0.05);不同土層相同月份間大型土壤動物類群數同樣無顯著差異(P>0.05)。
2.3 大型土壤動物群落多樣性動態特征
2.3.1 不同種植模式下大型土壤動物群落多樣性動態變化
Shannon-Weiner 指數在相同月份不同種植模式下無顯著差異(圖3a,P>0.05),相同種植模式下不同月份均表現為7月該指數最高,顯著高于6月(P<0.05)。Margalef 指數在相同月份不同種植模式下無顯著差異,相同種植模式下均表現為7月最高,且玉米/大豆間作種植下7月指數顯著高于9月(圖3b,P<0.05)。Pielou指數在相同月份不同種植模式下無顯著差異,6和9月玉米單作種植高于玉米/大豆間作種植,7和8月玉米/大豆間作種植高于玉米單作種植,相同種植模式下月份間同樣均無顯著差異(圖3c,P>0.05)。Simpson 指數在相同月份不同種植模式下無顯著差異;相同種植模式下月份間均在6月該指數最高,且玉米單作種植顯著高于7月(圖3d,P<0.05)。
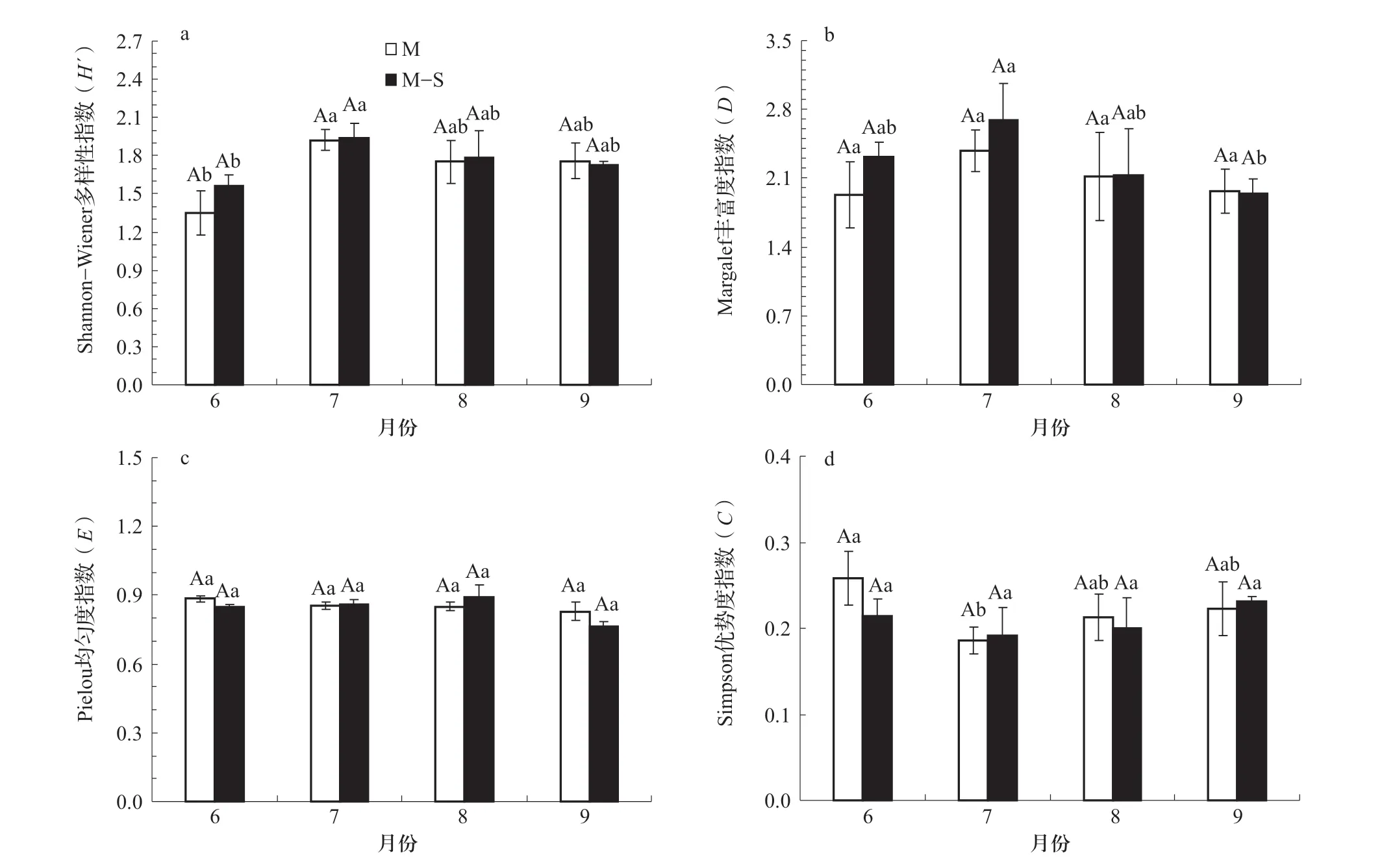
圖3 不同種植模式下大型土壤動物群落多樣性的變化
2.3.2 大型土壤動物群落多樣性垂直分布動態特征
Shannon-Weiner 指數在相同月份不同土層間,僅在6月0~10 cm 土層大于10~20和20~30cm土層,其余7~9月該指數均呈現0~10 <10~20 < 20~30 cm土 層 的 規 律,且 在8月20~30 cm土層該指數顯著高于0~10 cm土層(圖4a,P<0.05);在相同土層6~9月中0~10 cm 土層該指數呈現先下降后上升的規律,10~20 cm土層該指數呈現緩慢上升的規律,20~30 cm土層該指數呈現先上升后下降再上升的規律(7月該指數最高)。Margalef指數在相同月份不同土層下6月0~10 cm土層顯著高于10~20 cm土層(圖4b,P<0.05),9月0~10 cm土層該指數顯著高于10~20和20~30 cm土層(P<0.05);在相同土層6~9月中0~10 cm 土層和20~30 cm土層呈現規律相反,0~10 cm 土層該指數呈現緩慢上升的規律,20~30 cm土層呈現緩慢下降的規律,而10~20 cm土層呈現先上升后下降的規律(7月該指數最高)。Pielou 指數在相同月份不同土層下6~8月20~30 cm土層顯著高于10~20 cm土層(圖4c,P<0.05);在相同土層6~9月中0~10和10~20 cm土層該指數無明顯變化規律,20~30 cm土層呈現緩慢下降的規律。Simpson 指數在相同月份不同土層下6月10~20 cm土層顯著高于0~10 cm土層(圖4d,P<0.05),8和9月20~30 cm土層該指數顯著高于0~10 cm土層(P<0.05)。
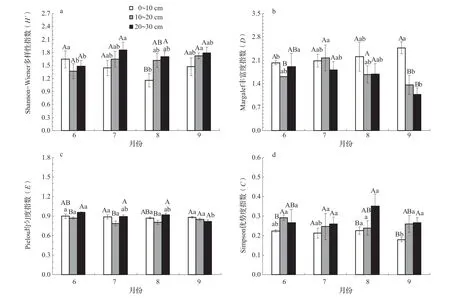
圖4 大型土壤動物群落多樣性垂直分布變化
2.4 大型土壤動物群落功能群動態特征
2.4.1 不同種植模式下大型土壤動物群落功能群動態變化
植食性大型土壤動物個體數在相同月份不同種植模式下,僅在8月間作種植低于單作種植,其余6、7和9月間作植食性大型土壤動物個體數較單作種植高,但無顯著差異(圖5a,P>0.05);同一種植模式6~9月,植食性大型土壤動物個體數均呈增加的趨勢,9月植食性大型土壤動物個體數與6和7月有顯著差異(圖5a,P<0.05)。捕食性大型土壤動物個體數在7月不同種植模式下,間作種植顯著高于單作種植(圖5b,P<0.05),并且在玉米各生育期7月間作種植捕食性個體數最多,與6月捕食性個體數有顯著差異(P<0.05)。腐食性大型土壤動物個體數在相同月份不同種植模式下間作種植高于單作種植,且在7月有顯著差異(圖5c,P<0.05);同一種植模式6~9月中,腐食性大型土壤動物個體數均呈增加的趨勢,9月腐食性個體數與6月腐食性個體數有顯著差異(P<0.05)。雜食性個體數在單作種植6月與7、8和9月的個體數有顯著差異(圖5d,P<0.05)。
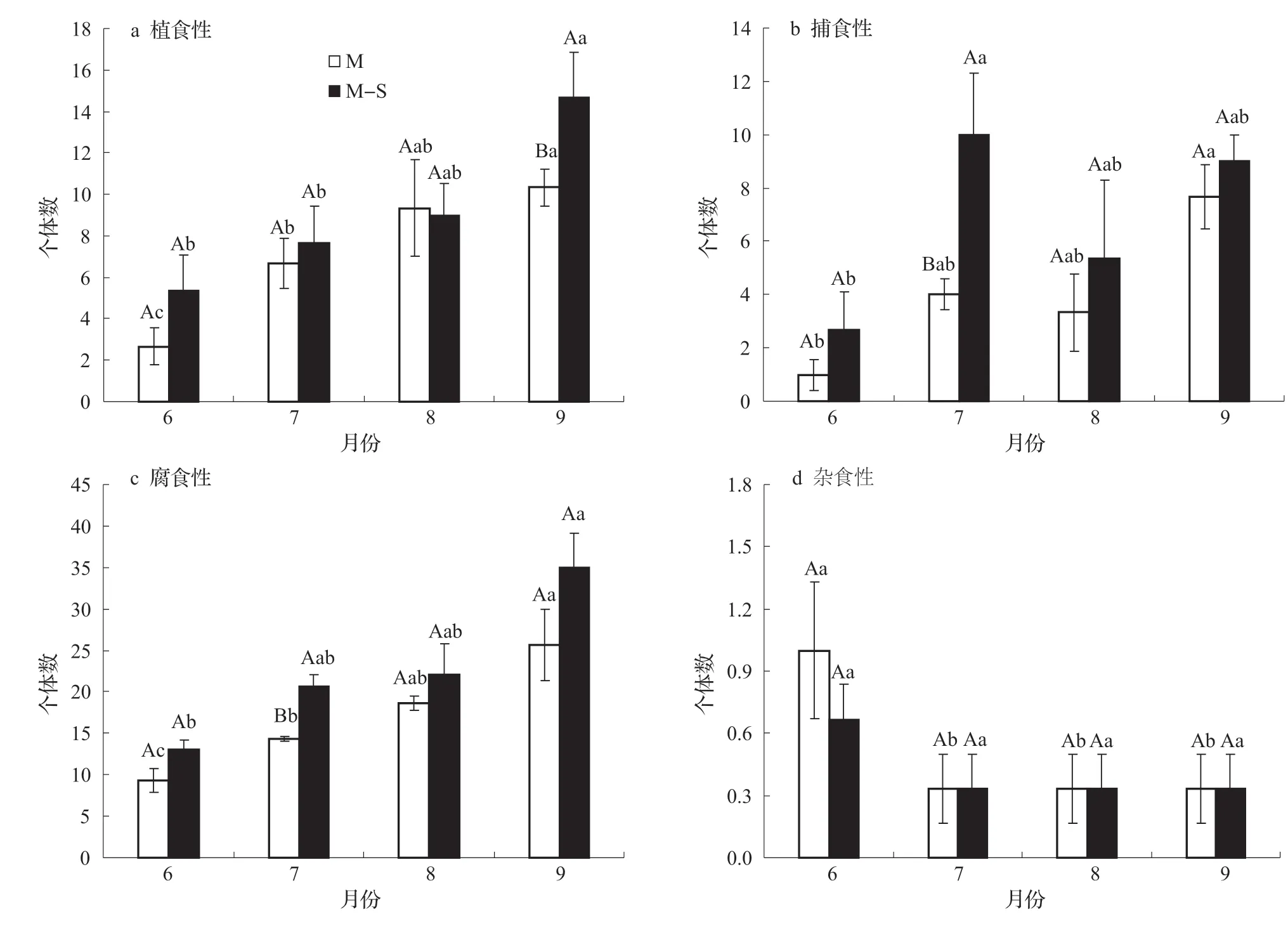
圖5 不同種植模式對大型土壤動物群落功能群的影響
2.4.2 大型土壤動物群落功能群垂直分布動態變化
植食性大型土壤動物個體數在相同月份不同土層下均表現為10~20 > 0~10 > 20~30 cm土層,并且6月10~20 cm土層植食性個體數與20~30 cm土層有顯著差異(圖6a,P<0.05),9月10~20 cm土層植食性個體數與0~10和20~30 cm土層有顯著差異(P<0.05);在相同土層6~9月中,植食性個體數呈增加趨勢。捕食性個體數在相同月份不同土層下只有9月的10~20與20~30 cm 土層有顯著差異(圖6b,P<0.05),其余月份無顯著變化;在相同土層6~9月中,同樣只有在9月中0~10和10~20 cm土層個體數分別與6月有顯著差異(P<0.05)。腐食性個體數在6月0~10 cm土層個體數最多,7~9月均是10~20 cm土層個體數最多,7月10~20 cm土層個體數與20~30 cm土層有顯著差異(P<0.05),8和9月10~20 cm土層個體數與0~10和20~30 cm土層均有顯著差異(圖6c,P<0.05);在相同土層6~9月中10~20和20~30 cm土層個體數呈現持續增加的趨勢,且10~20 cm土層個體數在各月均有顯著變化(P<0.05)。以線蚓科和隱翅甲科為主要的腐食性類群個體數主要分布在10~20 cm 土層中,距離玉米成熟期越接近,表層0~10 cm 土層的個體數比例越少。雜食性個體數在玉米生育期內垂直分布和月動態特征無明顯變化規律(圖6d,P>0.05)。
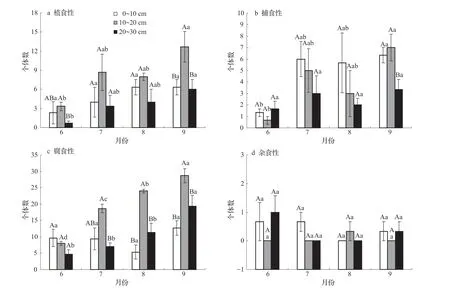
圖6 大型土壤動物群落功能群的垂直分布變化
3 討論與結論
3.1 玉米間作種植下土壤動物群落多樣性變化
土壤動物類群數和個體數的變化主要受土壤環境因子的影響,當自然環境變化或人為干擾強烈時,土壤動物群落結構通常發生相應改變[17]。農業生產中為了合理利用土壤資源,采取間作種植的措施,通過改變土地利用空間進而改變土壤的理化性狀。本研究表明,玉米/大豆間作種植在玉米的出苗、拔節、抽穗和成熟期均提高了大型土壤動物群落個體數(P>0.05),有研究表明,農田土壤動物個體數隨著外源氮濃度增加呈遞增的趨勢[15],而豆科-禾本科作物間作種植促進了作物氮素的吸收,導致土壤動物類群數和個體數增加;也有研究表明,長期施用氮肥增加了土壤微生物的種類和豐富度[18],能夠給土壤動物提供更多食物[19],從而有利于土壤動物個體數的增加;楊文亭等[20]在紅壤地定位試驗研究發現,種植模式和施氮水平對土壤動物平均密度和類群數均有一定的影響,施氮和間作種植有利于土壤動物生長。本研究在7月時,間作種植大型土壤動物個體數顯著高于單作種植,可能是由于7月土壤水熱條件較適合土壤生物生長發育[21],間作種植模式較單作種植具有更好的小氣候環境[22],適合大型土壤動物的生存。而本研究中相對于玉米單作種植,玉米/大豆間作種植在玉米的出苗、拔節和成熟期均提高了大型土壤動物的群落類群數,玉米抽穗期單作種植高于間作種植,可能是由于不同種植模式在玉米生長過程中的小氣候環境存在差異[22]。
土壤動物通常出現明顯的垂直分布,表現出表聚特性,土壤動物個體數和類群數隨土壤深度加深而降低[23],不同土壤類型降減速率可能不同[24],但在沙地或農業用地中土壤動物的垂直分布可能會出現逆向分布的現象[24-25]。農業生產活動促使大型土壤節肢動物向土壤下層移動,其中7月下移最為明顯[26]。本研究中單作與間作種植均采用翻耕耕作方式,玉米所有生育期內大型土壤動物類群數和個體數大體上是中層(10~20 cm)土層多于表層(0~10 cm)和下層(20~30 cm)土層;朱強根等[27]研究表明,耕作方式能夠對土壤動物群落結構產生一定影響,翻耕使10~20 cm土層的土壤動物數量增加,免耕秸稈覆蓋樣地在拔節期有助于提高土壤動物的多樣性[28]。王振海等[23]在長白山苔原帶研究表明,7月土壤動物優勢度指數降低,多樣性指數、豐富度指數和均勻度指數最高。本研究發現單作和間作種植下均在玉米拔節期(7月)土壤動物多樣性指數和豐富度指數最高,優勢度指數最低,其多樣性指數和豐富度指數高的原因可能是土壤溫度與水分適中,促進土壤微生物活動,適合土壤動物生存與繁殖,增加了多樣性。而楊旭等[29]在松嫩平原黑土耕作區研究表明,8月土壤動物均勻度指數和豐富度指數最高,優勢度指數最低。楊文亭等[20]在紅壤地研究發現,種植模式和玉米生育期的交互作用顯著影響了土壤動物密度類群和豐富度指數。綜上所述,空間、時間和管理措施的差異對土壤動物多樣性動態影響并不一致。
3.2 玉米間作種植下土壤動物群落功能群變化
玉米/大豆間作種植改變了農田不同食性土壤動物的個體數量、類群數以及多樣性。不同食性的土壤動物功能群是構成土壤地下食物網的基礎,在調節物質循環過程中起到重要作用。一般認為,相對穩定的土壤環境,其土壤動物的各食性比例保持一個穩定狀態[30]。植食性動物作為生態系統的初級消費者,其種類和數量等通常都高于其他幾個營養級的消費者[31],本研究表明,玉米/大豆間作種植對各食性土壤動物均有所提高。Quinn等[32]研究認為,施用氮肥后植物根組織氮濃度增加可能是導致植食性土壤動物數量增加的主要原因,在本研究中玉米/大豆間作種植模式下增加了玉米出苗、拔節和成熟期植食性土壤動物的個體數,且在玉米成熟期顯著提高了植食性土壤動物的個體數,植食性土壤動物的個體數主要分布在0~10和10~20 cm土層中。捕食性類群作為生態系統的次級消費者,在7月間作種植顯著提高了其個體數。以線蚓科和隱翅甲科為主要的腐食性類群隨玉米生長個體數增加,并且主要分布在10~20 cm 土層中,距離玉米成熟期越接近,表層(0~10 cm)土層的個體數比例越少。有研究表明,線蚓的數量能夠反映群落環境特點和土壤質地狀況,通常情況下土壤含水量高、有機質含量高的土壤中線蚓科的數量就多[33]。同時,腐食性類群能夠加快農田土壤生態系統物質循環和能量轉換的過程[34]。
