一株海南菌草“綠洲一號”內生細菌的分離鑒定及其生物學特性研究
2022-09-14 08:45:18李望豪周瑞鵬許鈺瀅趙琪琪王榮波葉文雨
廣東農業科學 2022年7期
關鍵詞:植物
李望豪,周瑞鵬,許鈺瀅,趙琪琪,劉 娟,王榮波,葉文雨
(1.福建農林大學國家菌草工程技術研究中心,福建 福州 350002;2.福建農林大學生命科學學院/植物與微生物相互作用福建省高校重點實驗室,福建 福州 350002;3.福建農林大學園藝學院,福建 福州 350002;4.福建省作物有害生物監測與治理重點實驗室,福建 福州 350013)
【研究意義】菌草可作為靈芝等食、藥用菌培養基的成分。海南菌草“綠洲一號”是由國家菌草工程技術研究中心選育出的菌草品種之一。該品種在水土保持,治理鹽堿地,抑制病原菌及作為食、藥用菌新型培養基原料等方面都有廣闊的發展前景[1-5]。“綠洲一號”是菌草中較為重要的品種,明確“綠洲一號”內生細菌的生物學特性及其更深層次的機理,可以為菌草資源利用及其可持續發展提供理論支持,也為植物病原真菌的生物防治及植物促生機理的研究奠定基礎。【前人研究進展】植物內生細菌是指可以長期在植物體內生存,且會對植物的生長發育產生一定益生作用的一類細菌[6-9]。目前,從植物組織分離的內生細菌是當今微生物領域研究的一大熱點,其主要研究方向為內生細菌的促生能力和拮抗能力。Khanna等[10]從鷹嘴豆(Cicer arietinumL.)中分離出對其枯萎病具有拮抗作用且可促進鷹嘴豆植株生長的菌株;曹鵬飛等[11]從楊梅等植物的組織中分離出對楊梅枯萎病病菌拮抗效果良好的內生細菌;白潔等[12]從歐李根部分離得到7株內生細菌,均具有一定的溶磷和產IAA 能力,從而對植物產生促生作用;錢鑫等[13]從健康的黃瓜植株中分離出對菊苣假單胞菌(Pseudonwnas cichorii)有抑菌作用的內生細菌,該菌具有很好的固氮、溶磷等促生作用,在抵抗病原真菌的同時還可以為黃瓜提供生長所需的微量元素,從而促進黃瓜生長。【本研究切入點】前人對經濟作物和糧食作物內生細菌的研究較為深入,近年來也逐漸重視對菌草等植物內生細菌的研究。鑒于“綠洲一號”菌草用途廣泛,其內生細菌功能多樣,故對其內生細菌進行相關研究。蘇云金芽孢桿菌(Bacillus thuringiensis)可作為微生物殺蟲劑,其在植物病蟲害防治中起到積極作用,并且近年來得到更深入的研究與應用[14-15]。目前蘇云金芽孢桿菌大多是從土壤和部分植物組織中分離得到,而從菌草分離得到蘇云金芽孢桿菌的研究鮮有報道。【擬解決的問題】本研究從海南菌草“綠洲一號”分離得到1 株內生細菌,并對其生理生化特性、拮抗病原真菌以及其促生性能等方面展開研究,為植物病原真菌的生物防治及植物促生機理研究提供基礎理論。
1 材料與方法
1.1 試驗材料
供試植物為海南菌草基地的“綠洲一號”(Juncao“Oasis No.1”),2020 年10 月采其葉片作為內生細菌的分離材料。
供試病原真菌為稻瘟病菌(Magnaporthe oryzae,Guy11)、香蕉枯萎病菌4 號(Foctropical race4,Foc4),病原真菌均保存在植物與微生物相互作用福建省高校重點實驗室。
供試培養基包括細菌生長培養基:LB 固(液)體培養基[16];真菌菌絲生長培養基:PDA 培養基[17];生理生化培養基:淀粉水解培養基[18]、明膠培養基[19]、脫脂奶粉培養基、檸檬酸鹽利用培養基、酯酶檢測培養基、纖維素水解培養基;促生性能培養基:有機磷培養基[20]、無機磷培養基[21]、CAS 檢測培養基[22]、硅酸鹽細菌培養基[23]、King 培養基[24]。
1.2 試驗方法
1.2.1 內生細菌的分離與純化 將新鮮葉片用自來水沖洗,洗凈后用75%酒精浸泡5 min,然后用無菌水沖洗3 次以上,將最后一次無菌水涂布于LB 培養基中,以確定表面是否消毒徹底。將葉片剪碎后放入研缽,加入無菌水充分研磨,吸取上清液并將上清液稀釋至10-1、10-2、10-3倍,然后分別涂布于LB 培養基上。培養1 d 后挑取合適的菌落,純化3 次后加甘油,以菌液︰甘油為3︰1 的比例對分離純化得到的內生細菌進行保藏。
1.2.2 內生細菌拮抗病原真菌的篩選 采取平板對峙法,將提前活化的稻瘟病菌和香蕉枯萎病菌4 號切成直徑5 mm 的菌餅,菌絲向下接種在培養基中心。再將提前活化16 h 的內生細菌分別添加在距離中心菌餅3.5 cm 處,待對照組病原真菌基本長滿平板時記錄對照組和試驗組病原真菌菌落半徑,計算抑菌率:

1.2.3 內生細菌的分子鑒定 活化內生細菌16 h 后,利用菌液作為模板,采用引物27F 和1492R[25],PCR 條件及反應體系參考劉彥超等[25]的方法,PCR 產物送至生物工程(上海)股份有限公司進行測序。將測序結果與NCBI 數據庫中的已知序列進行BLAST 比對,利用 MEGA7.0 軟件采用近鄰相接法構建出系統發育樹。
1.2.4 內生細菌的生理生化特性分析 將內生細菌活化16 h 后進行生理生化特性鑒定[26-28],相關試驗包括革蘭氏染色、檸檬酸鹽利用試驗、纖維素利用試驗、明膠液化試驗、酯酶試驗、淀粉水解試驗、蛋白酶試驗和接觸酶試驗。
1.2.5 內生細菌的促生能力分析 將內生細菌活化16 h 后進行該菌對植物的促生能力分析[29-32],相關試驗包括溶磷能力鑒定、合成鐵載體能力鑒定、降解硅酸鹽能力鑒定以及分泌IAA 能力鑒定。
2 結果與分析
2.1 內生細菌HLY3-2 的形態學特征
從海南菌草“綠洲一號”葉片上分離得到1株內生細菌(編號:HLY3-2)。從圖1A 可見,HLY3-2 菌落呈紅色圓形,邊緣規則、表面光滑、濕潤、粘稠狀,菌落直徑為1~2 mm。經革蘭氏染色為陽性,菌體呈桿狀(圖1B),初步鑒定該內生細菌為芽孢桿菌(Bacillussp.)。
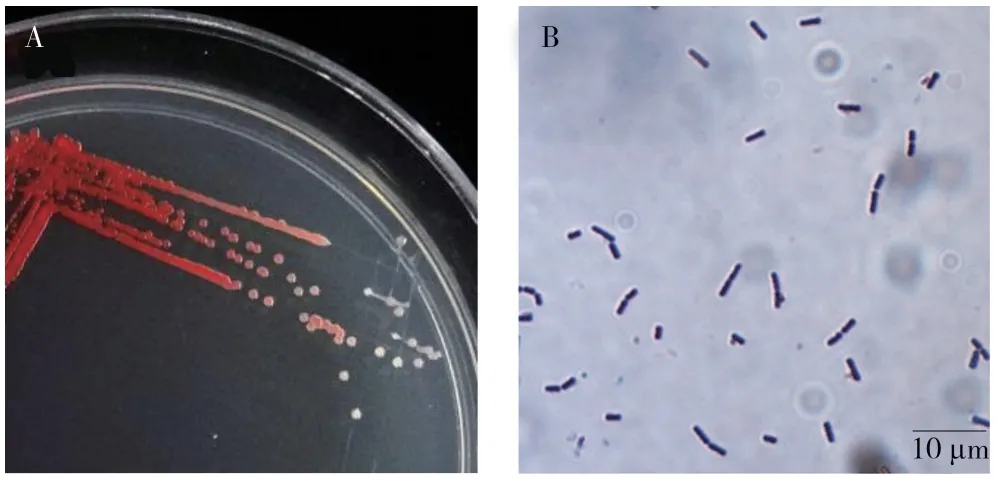
圖1 內生細菌HLY3-2 的形態學特征Fig.1 Morphological characteristics of endophytic bacterium HLY3-2
2.2 內生細菌HLY3-2 拮抗病原真菌的篩選
內生細菌HLY3-2 拮抗病原真菌篩選試驗結果表明,該菌對稻瘟病菌和香蕉枯萎病菌4 號兩種病原真菌均具有一定的抑菌作用(圖2)。通過測量對照組病原真菌菌落直徑與試驗組病原真菌菌落直徑,得出內生細菌HLY3-2 的抑菌率分別為 18.45%和30.65%(圖3)。
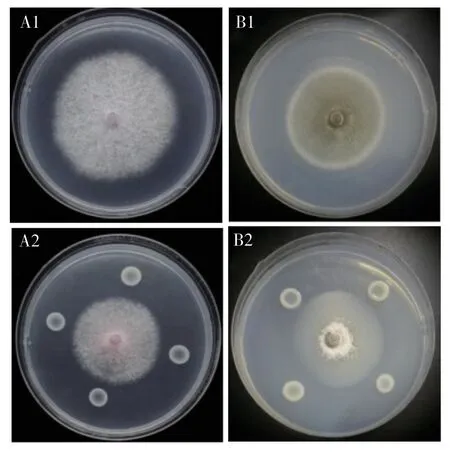
圖2 內生細菌HLY3-2 對病原真菌的拮抗作用Fig.2 Antagonism of endophytic bacterium HLY3-2 against pathogenic fungi
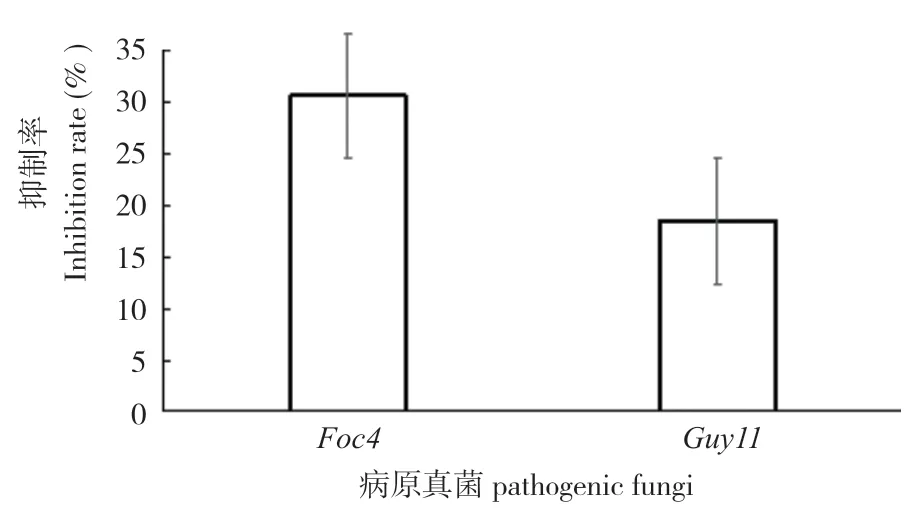
圖3 內生細菌HLY3-2 對病原真菌的抑制率Fig.3 Inhibition rates of endophytic bacterium HLY3-2 against pathogenic fungi
2.3 內生細菌HLY3-2 的分子鑒定
采用通用引物對菌液進行PCR 擴增,經1.2%瓊脂糖凝膠電泳,結果(圖4A)顯示,HLY3-2菌株的16SrDNA 基因片段大小約1 600 bp。測序得到16S rDNA 基因序列后,將序列與NCBI 數據庫中的已知序列進行BLAST 比對,將具有同源性高的序列下載,利用MEGA7.0 軟件進行同源比對,采用鄰近相接法構建系統發育樹,結果(圖4B)表明,內生細菌HLY3-2 與蘇云金芽孢桿菌(Bacillus thuringiensis)匯在一支且相似度高達到99%,故初步判斷該菌為蘇云金芽孢桿菌。
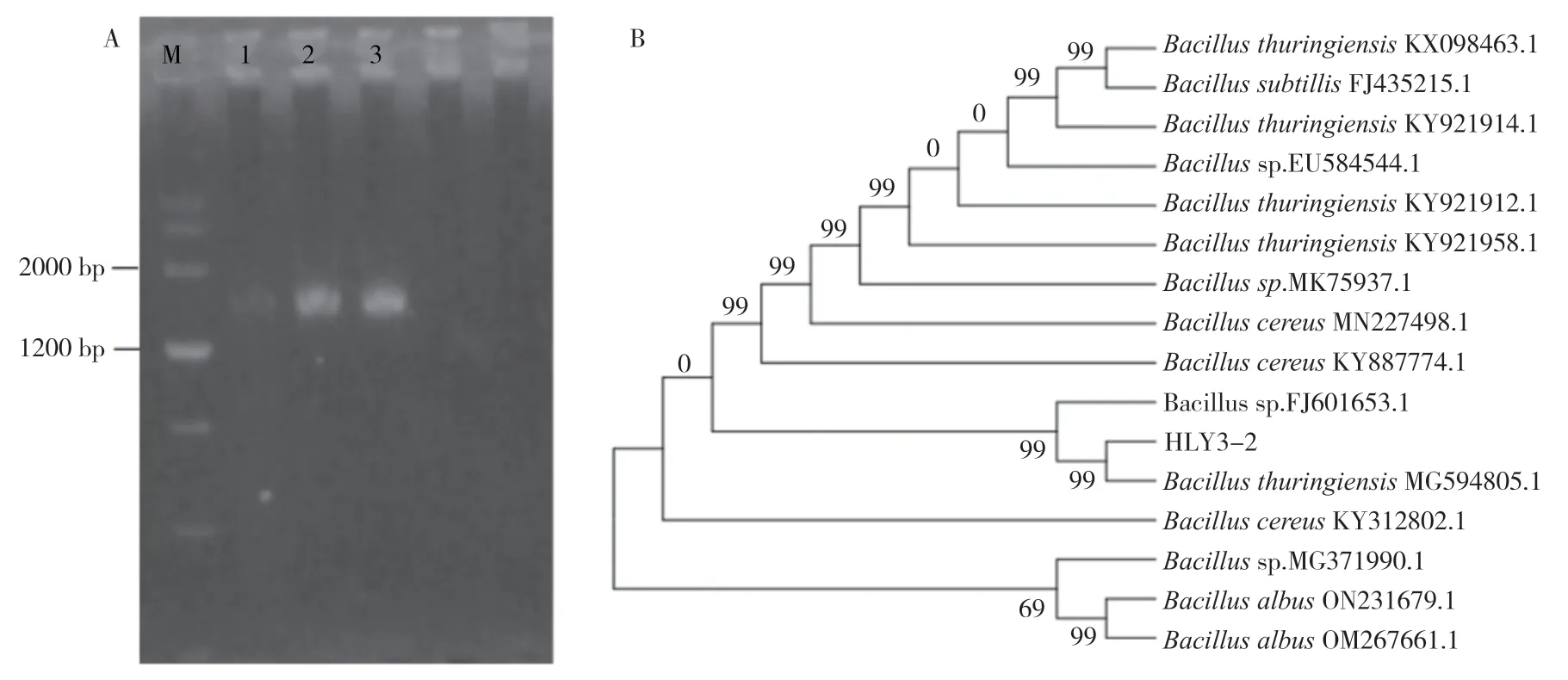
圖4 內生細菌HLIY3-2 的分子鑒定Fig.4 Molecular identification of endophytic bacterium HLY3-2
2.4 內生細菌HLY3-2 的生理生化特性
在檸檬酸鹽利用試驗培養基中,菌落周圍沒有產生變化,說明HLY3-2 菌株無法利用檸檬酸鹽(圖5A);向纖維素利用試驗培養基中加入剛果紅后,菌落被沖散且無透明圈產生,表明該菌株不具有利用纖維素的能力,從而導致菌落沒有很好地固定在培養基上(圖5B);向裝有明膠培養基的EP 管中接種菌液,明膠沒有液化,故該菌株沒有使明膠液化的能力(圖5C);菌落在酯酶培養基上沒有產生透明暈圈,可見其為不產酯酶菌株(圖5D);向接種該菌株且恒溫培養3 d 的淀粉培養基中加入碘液,2 min 后菌落周圍沒有產生透明圈,故其為不產淀粉水解酶菌株(圖5E);在奶粉培養基上,菌落周圍產生了明顯的透明圈,故該菌株可以產生蛋白酶(圖5F);向涂有該菌落的載玻片上滴入過氧化氫溶液后,迅速產生大量氣泡,有氧氣產生,故該菌株可以產生過氧化氫酶(圖5G)。
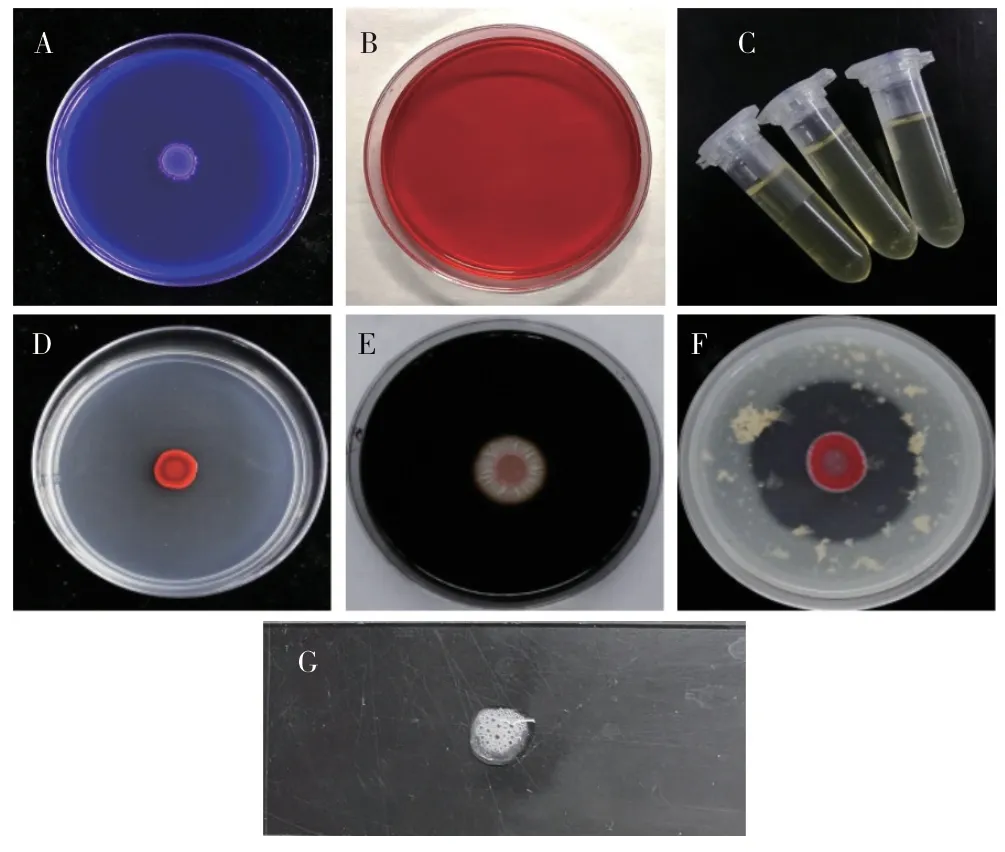
圖5 內生細菌HLIY3-2 的生理生化特性分析Fig.5 The physiological and biochemical characteristics of endophytic bacterium HLY3-2
2.5 內生細菌HLY3-2 的促生能力
在含有機磷和無機磷的培養基上,菌落周圍都產生了透明圈,因此HLY3-2 菌株具有溶解有機磷和無機磷的能力(圖6A 和圖6B);在CAS培養基上,菌落周圍培養基沒有產生黃色暈圈,故該菌株沒有產生鐵載體的能力(圖6C);在硅酸鹽細菌培養基上,菌落周圍產生了透明圈,故該菌株可以降解硅酸鹽,為植物提供鉀、硅等微量元素(圖6D);通過標準比色法可知,菌液的發酵液得到的顏色深于50 μg/mL IAA的標準溶液,因此該菌株具有很強的產IAA 能力(圖6E)。綜上可見,HLY3-2 菌株具有一定的促生能力。
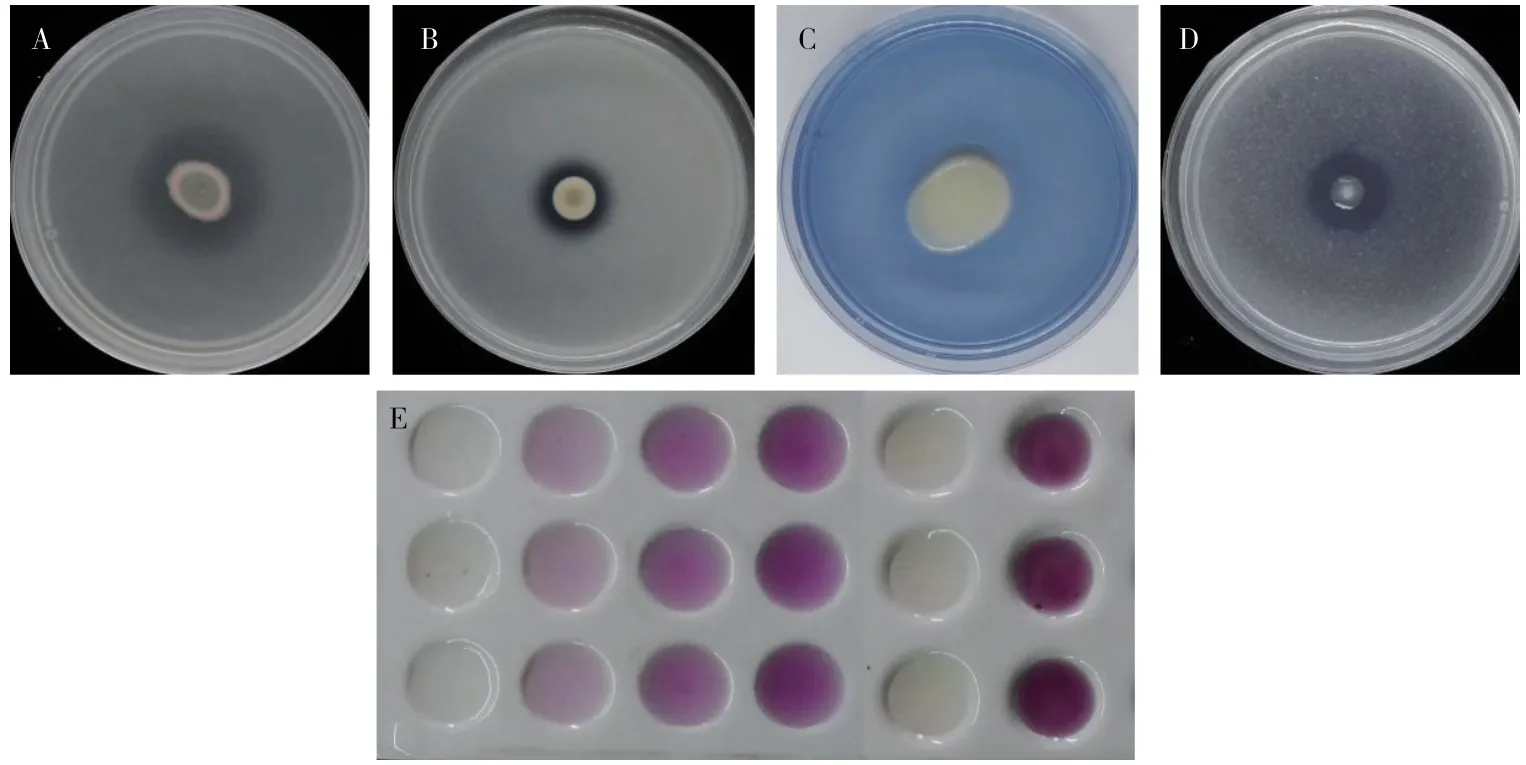
圖6 內生細菌HLY3-2 的促生能力分析Fig.6 Analysis of growth promoting ability of endophytic bacterium HLY3-2
3 討論
在菌草內生細菌研究方面,鄧振山等[33]從巨菌草植株中共分離得到187 株內生細菌,其中86 株具有分泌IAA 的能力,47 株具有溶磷能力。本研究從海南菌草“綠洲一號”葉片中分離得到的內生細菌HLY3-2 具有類似的生物學性能,經過分子與形態學鑒定,該內生細菌為蘇云金芽孢桿菌。該菌株與大多數植物組織中分離得到的內生拮抗促生細菌同為芽孢桿菌屬,但并非同一種。“綠洲一號”植株高大、根系發達、抗逆性強、營養豐富,可以為更多內生細菌的生存提供條件,后續仍需進一步驗證。
在促生機理研究方面,張萍等[34]對蘭科植物內生細菌進行研究,發現內生細菌可以通過溶解磷元素、鉀元素和產生IAA 來促進植物生長。而本研究分離得到的內生細菌HLY3-2 具有溶解無機磷、有機磷及降解硅酸鹽的能力,并且可以產生IAA。因此該內生細菌HLY3-2 可為植株提供充足微量元素和生長素,進而促進植物生長,故該菌株具有開發成為促生菌劑的潛力。
在內生細菌拮抗病原真菌方面,周維等[35]從香蕉中分離出可以對Foc4病原真菌產生拮抗的內生細菌;郭垂寶等[36]從龍牙百合磷莖中分離出對灰葡萄孢菌(Botrytis cinerea)產生強拮抗作用的內生細菌;孫宇晨等[37]從百合的莖分離得到具有促生效應的菌株。以上試驗可以證實植物內生細菌不僅對宿主有促生作用,還可以增強宿主對病害的抵御能力。本研究分離得到1 株拮抗內生細菌屬于蘇云金芽孢桿菌,該菌種的促生、拮抗作用未見報道,因此其可為內生細菌進行生物防治提供新思路和新方法。
在蘇云金芽孢桿菌抗病研究方面,李輝等[38]從疏花水柏枝葉片中篩選出對魔芋軟腐病病菌拮抗效果較好的蘇云金芽孢桿菌,但是對Foc4和Guy11病原真菌的拮抗作用尚未涉及。而本研究分離得到的蘇云金芽孢桿菌對Foc4和Guy11均具有一定的抑菌作用,這可能是由于不同宿主對內生細菌的影響不同,從而產生不同抑菌效果。
在拮抗病原真菌機理方面,劉洋等[39]從核桃根部篩選出拮抗細菌,并對其拮抗病原真菌機理進行了深入研究,表明內生細菌可以通過分泌多種酶(如蛋白酶、纖維素酶)降解病原真菌或抑制病原真菌的生長與繁殖。但是本研究分離得到的內生細菌在生理生化鑒定時表現出可以產生蛋白酶但無法產生纖維素酶的特性,因此其拮抗病原真菌的機理可能與上述研究不同。這可能是由于不同種屬的內生細菌生境不同,拮抗病原真菌的機理也不相同。內生細菌HLY3-2 有望在生物防治、生物農藥開發、菌草產業發展,以及促進國家農業健康發展等方面發揮一定作用。
4 結論
本研究從海南菌草“綠洲一號”中分離得到內生細菌HLY3-2,經鑒定,該菌株為蘇云金芽孢桿菌,具有溶解無機磷、有機磷和降解硅酸鹽的能力,而且可以產生IAA,兼具促進植物生長的能力。此外,該菌株可以產生蛋白酶、過氧化氫酶,可以在一定程度上清除植物體內的活性氧,利于植物健康生長。內生細菌HLY3-2 對病原真菌Foc4和Guy11具有一定拮抗作用。因此本研究分離得到的蘇云金芽孢桿菌可以促進植物生長和抑制病原真菌,為多功能生物菌肥[40]及生防菌的應用提供理論依據。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
