谷子SiPRR73基因的光溫調控模式 及非生物脅迫響應特性
2022-09-14 04:22:16閆留延李劍峰張世文王永芳張小梅祖超凡王振山桑璐曼何占祥賈小平董志平
華北農學報 2022年4期
閆留延,李劍峰,張世文,張 博,王永芳,張小梅,祖超凡,王振山, 桑璐曼,何占祥,賈小平,董志平
(1.河南科技大學 農學院,河南 洛陽 471023;2.河北省農林科學院 谷子研究所,國家谷子改良中心,河北 石家莊 050035)
PRRs(Pseudo-response regulators)家族基因廣泛參與植物花期調控、生物量積累及對逆境脅迫的抵御作用[1-4]。作為PRR家族的重要成員,PRR73是擬南芥(Arabidopsisthaliana)PRR7的直系同源基因,目前在C4作物中報道較少,只在玉米(Zeamays)中利用圖位克隆法獲得了該基因[5]。作為我國重要的雜糧作物,谷子(Setariaitalica)遺傳資源豐富,地理分布廣泛,存在較多的光溫敏感自然變異類型,是C4禾谷類作物光溫敏感調控機制研究的理想模型[6-9]。研究谷子PRR73基因對光溫及非生物脅迫的響應特點,可以了解光周期和溫度如何共同調控PRR73基因表達來實現谷子對不同光溫環境的適應性調節,揭示PRR73基因參與非生物脅迫調節的可能機制,具有重要意義。
PRRs家族蛋白質包含3個區域:PRR結構域、CCT結構域和可變域,分別位于蛋白質的N端、C端和中部[10-13]。被子植物PRRs基因的PRR、CCT結構域高度保守,但PRR結構域中個別氨基酸變化可以導致基因功能改變[14-15]。對玉米ZmPRR73的研究表明,在不保守的可變域中氨基酸的變化可能也影響光周期敏感性[5]。擬南芥PRR7通過CONSTANS依賴途徑控制開花時間,可以阻遏CCA1和LHY啟動子的活性,調節CCA1和LHY響應環境溫度和調節細胞內活性氧的水平[12-13,16-20]。此外,PRR7能夠負調控抗冷基因CBF和調節ABA來響應冷脅迫和干旱脅迫,參與植物對鐵過量的適應性反應[20-24]。谷子和其他禾本科作物一樣,光周期調控開花研究已經有所報道,但光周期和溫度如何共同調控生長發育的研究則極少涉及[25-33]。事實上光周期和溫度通過復雜的互作效應影響谷子生長發育,一些光周期途徑的關鍵基因如SiCCT參與了谷子光溫互作調控[34]。
作為光周期調控途徑的另一個關鍵基因,PRR73在C4作物光溫互作調節以及應對非生物脅迫中是否起到一定作用是值得研究的一個問題。本研究從谷子中克隆SiPRR73基因,系統分析其對光周期、光溫組合以及5種非生物脅迫的響應特性,以揭示光周期和溫度對SiPRR73基因的調控模式以及這種調控對谷子適應不同光溫環境的潛在意義,并揭示SiPRR73基因參與抵抗非生物脅迫的調節機制。
1 材料和方法
1.1 試驗材料
所用的谷子品種延谷11號來自陜西,是對光溫較敏感的春谷品種。
1.2 材料種植及樣品采集
采用盆栽種植,所用塑料盆規格為10 cm×10 cm,盆內裝有營養土,種植120盆,每盆播種5~8粒種子,置于25 ℃、14 h光/10 h暗的培養室長至三葉期定苗,其中部分植株在五葉期采集嫩葉用于克隆SiPRR73基因;另有部分植株繼續培養,抽穗后采集根、莖稈、次頂葉、頂葉、穗頸、幼穗于液氮中保存。設置長日照(25 ℃、15 h光/9 h暗,LD)、短日照(25 ℃、9 h光/15 h暗,SD)2個不同光周期處理,每個處理24盆,其中,長日照設定21:00—6:00為黑暗, 6:00—21:00為光照;短日照設定15:00—6:00為黑暗, 6:00—15:00為光照(下同)。在六葉期時于6:00開始取樣,取頂端嫩葉,每隔2 h取樣一次,連續48 h取樣,此外從三葉期開始于9:00取樣,每隔3 d取樣一次,短日照取至八葉期(抽穗),長日照取至十二葉期,所有樣品于液氮保存。設置4種光溫組合處理:低溫長日照(溫度22 ℃、光周期15 h光/9 h暗,LTLD)、低溫短日照(溫度22 ℃、光周期9 h光/15 h暗,LTSD)、高溫長日照(溫度27 ℃、光周期15 h光/9 h暗,HTLD)、高溫短日照(溫度27 ℃、光周期9 h光/15 h暗,HTSD);每個處理16盆,培養至六葉期從6:00開始取樣,每隔3 h取樣一次,連續24 h取樣,所采頂端嫩葉于液氮中保存。
用于非生物脅迫處理的谷苗種植條件同上,每個脅迫處理和對照處理均8盆谷苗,在培養室(溫度為25 ℃,光照條件為14 h光/10 h暗)培養21 d,隨后進行脅迫處理,NaCl、ABA、EDTA-Fe、PEG6000的處理濃度分別為200 mmol/L、100 μmol/L、600 μmol/L、20%,低溫處理溫度為15 ℃。對照組(溫度為25 ℃,光照條件為14 h光/10 h暗,CK)和每個處理組取樣時間均為處理前(0 h),脅迫處理后0.5,1,2,4,8,16,24 h,所取的頂端嫩葉于液氮中保存。
1.3 SiPRR73的克隆及生物信息學分析
葉片總RNA的提取及反轉錄分別用康為世紀RNA提取試劑盒及寶生物公司第一鏈cDNA合成試劑盒完成,基因特異引物信息見表1。基因擴增體系為20 μL,包括cDNA模板1 μL,2×Es Taq MasterMix(Dye)10 μL,引物(正向與反向混合)0.5 μL,剩余體積用ddH2O補足。擴增循環程序: 94 ℃預變性4 min;變性(94 ℃/30 s)、退火(58 ℃/30 s)、延伸(72 ℃/90 s),循環35次;最后72 ℃延伸 8 min。PCR產物用全式金的膠回收試劑盒純化回收,回收產物連入pBM16A載體,轉化DH5α感受態細胞,挑選陽性克隆進行測序。
利用NCBI的BlastP程序(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)搜索保守結構域;從公共數據庫下載其他物種的43個PRR73蛋白序列,利用 ClustalW 1.8軟件對包含谷子SiPRR73蛋白的44條PRR73蛋白序列進行多序列比對,系統進化樹構建采用MEGA 6.05軟件。
1.4 谷子SiPRR73基因的表達分析
按照1.3步驟完成樣品RNA的提取及第一鏈cDNA的反轉錄合成,谷子內參基因SiActin和SiPRR73基因分別用表1中對應的特異引物(SiActin-FQ、SiPRR73-FQ)進行擴增。擴增體系按照TB GreenTM 試劑盒(寶生物公司)說明書準備,使用羅氏熒光定量PCR儀擴增,兩步法擴增程序:預變性(95 ℃/30 s);40個循環包括變性(95 ℃/30 s)、退火(58 ℃/30 s)。基因相對表達量的計算用2-ΔΔCt方法。

表1 基因擴增及熒光定量PCR引物Tab.1 Primers used for gene amplification and fluorescence quantitative PCR
2 結果與分析
2.1 SiPRR73基因結構域、進化關系分析
從谷子葉片提取高質量總RNA(圖1-A),反轉錄合成第一鏈cDNA,利用RT-PCR技術分2段擴增SiPRR73基因,擴增產物SiPRR73-G1、SiPRR73-G2經電泳檢測與目標片段相符(圖1-B)。回收片段經測序后拼接得到2 928 bp的cDNA序列,其中CDS長度為2 283 bp,推定的蛋白質氨基酸數量為760個。SiPRR73 蛋白含有REC、CCT等2個結構域(圖2)。系統進化分析表明,C4作物家族的谷子、高粱(Sorghumbicolor)、糜子(Panicummiliaceum)、玉米(Zeamays)、哈氏黍(Panicumhallii)PRR73蛋白具有較近的進化關系(圖3)。

A.谷子總RNA電泳:1,2.兩管RNA。B.SiPRR73基因的RT-PCR產物電泳:1.DL2000 Marker;2,3.分段擴增產物SiPRR73-G1、SiPRR73-G2。
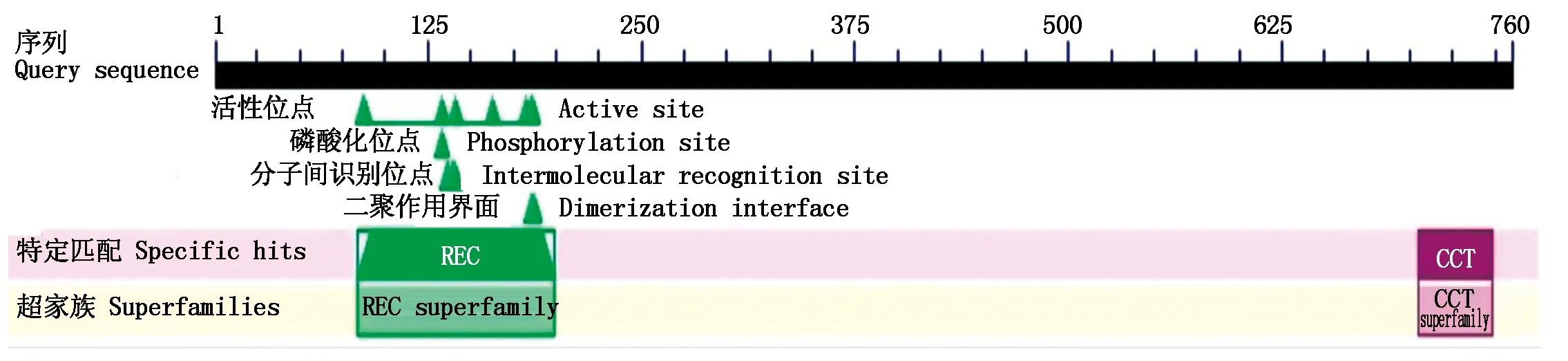
圖2 SiPRR73蛋白所包含的結構域Fig.2 The domains contained in SiPRR73 protein

圖3 PRR73蛋白的分子系統進化樹Fig.3 Molecular phylogenetic tree of PRR73 proteins
2.2 谷子不同組織部位SiPRR73基因的表達特性
SiPRR73基因具有明顯的組織表達特異性,在次頂葉表達水平最高,其次為頂葉,隨后依次為穗頸、幼穗、莖稈、根(圖4)。
2.3 SiPRR73基因對光周期的響應特點
兩晝夜基因表達模式一致,因此,圖5-A中只展示24 h表達模式。短日照條件下,SiPRR73基因在光照8 h后(14:00)表達量達到峰值,隨后開始下降,在整個晝夜中呈節律性表達,只出現一個表達峰。長日照條件下,SiPRR73基因在8:00表達量迅速升高,在10:00和14:00出現表達峰值,14:00后表達量緩慢下降。總之,SiPRR73基因呈現出光依賴性晝夜節律表達模式,短日照下只有一個表達峰,長日照下有2個表達峰,且無論長日照還是短日照光照期表達水平總體高于黑暗期。從圖5-B可以看出,總體上短日照條件下從三葉期到八葉期(抽穗)SiPRR73基因的表達水平要低于長日照,長日照條件下八到十二葉期仍維持營養生長,SiPRR73在十一、十二葉期表達量升至較高水平。

不同小寫字母表示處理間差異顯著(P<0.05)。圖5—7同。 Different lowercase letters indicate significant differences between treatments at P<0.05. The same as Fig.5—7.

A.晝夜表達規律;B.不同葉期表達規律。 A. Diurnal expression pattern;B. Expression pattern at different leaf stages.
2.4 光周期和溫度對SiPRR73基因表達的調控作用
溫度對SiPRR73基因表達峰的數目有明顯影響,在高溫(27 ℃)條件下的光照期SiPRR73出現2個表達峰,而在低溫(22 ℃)條件下的光照期只出現1個表達峰,光周期的改變并不影響高、低溫間表達峰數目的差異;光周期控制SiPRR73表達峰出現的早晚,溫度的改變并不影響SiPRR73在短日照條件下的表達峰比長日照下提前的特點。此外,SiPRR73表現出一定光誘導表達特性,黑暗期表達水平相對較低(圖6)。
2.5 非生物脅迫條件下SiPRR73基因的表達模式
NaCl處理1 h內SiPRR73基因受誘導表達,隨后的時間除了8 h誘導表達,其余時間點基因表達均處于抑制狀態(圖7-A)。SiPRR73基因在ABA處理1,8 h受誘導表達,其余時間點表達基本處于抑制狀態(圖7-B)。PEG模擬干旱脅迫除了脅迫0.5,24 hSiPRR73表達受到抑制,其余時間點均處于誘導表達狀態,特別是處理8,12 h,基因表達最強(圖7-C)。SiPRR73在低溫(15 ℃)處理24 h內均受誘導表達,4,8 h表達最強,說明SiPRR73在應對低溫脅迫中可能發揮功能(圖7-D)。在EDTA-Fe 處理下,SiPRR73在早期(2 h內)表達受到抑制,4~18 h均處于誘導表達狀態,24 h表達再次受到抑制,推測其在鐵脅迫過程中發揮特定作用(圖7-E)。5種非生物脅迫中,PEG和低溫總體上誘導SiPRR73表達,而NaCl在脅迫早期誘導基因表達,后期總體表現為抑制狀態,Fe脅迫則早期抑制基因表達,后期總體誘導基因表達。SiPRR73對ABA脅迫的響應特點接近NaCl,主要差異在于脅迫0.5 h NaCl誘導基因表達而ABA抑制基因表達。

圖6 光溫互作對SiPRR73基因晝夜表達模式的影響Fig.6 The effect of photo-thermal interaction on diurnal expression pattern of SiPRR73

圖7 非生物脅迫條件下SiPRR73基因的表達模式Fig.7 Expression pattern of SiPRR73 under abiotic stress conditions
3 結論與討論
通過對擬南芥、水稻、高粱、玉米4種植物CCT結構域基因進化分析,發現水稻OsPRR73基因劃分到第4組,該組包括擬南芥2號、5號染色體上的一些CCT結構域基因[35]。本研究發現,谷子SiPRR73與水稻、玉米、小麥、高粱等禾本科家族PRR73基因聚為1組,但在組內又分為3個亞組,谷子與玉米、糜子、高粱等C4作物PRR73基因聚為1個亞組,水稻、小麥PRR73基因分別聚為另外2個亞組。組織特異性表達分析發現,谷子SiPRR73和玉米ZmPRR73、水稻OsPRR73一樣,均在葉片中的表達量最高,受光調控,而小麥TaPRR73基因則在根部表達量較高,說明PRR73基因在禾本科家族內部進化過程中功能發生了分化[4-5,36]。谷子SiPRR73在長、短日照條件下的晝夜表達模式與水稻OsPRR73、小麥TaPRR73相似,即在溫度不變的情況下,長日照有2個表達峰,短日照有1個表達峰,與玉米ZmPRR73長短日照下均只有1個表達峰存在差異[4-5,37],說明盡管同為C4作物,谷子和玉米PRR73基因對光周期的響應模式存在一定差異。溫度是光周期外影響植物生長的另一個重要環境因子,二者存在復雜的互作效應。對擬南芥的研究發現,提高溫度能抵消短日照對開花的抑制作用[38];而大麥和大豆都是在誘導開花的光周期條件下高溫促進開花,抑制開花的光周期條件下高溫抑制開花[39-42]。谷子光溫互作模式與大豆相似,短日照和高溫對抽穗具有正向作用,長日照和高溫對抽穗具有負向效應[34]。本研究發現,光周期和溫度均對SiPRR73的表達模式產生了影響,但影響方式不同,光周期主要影響了基因表達峰出現時間的早晚,溫度主要影響了表達峰的數目,而這種影響會導致基因功能發生改變,從而影響谷子抽穗開花,這表明SiPRR73可能參與谷子對不同光周期和溫度環境的適應性調節過程。
PRRs家族基因與植物對非生物脅迫的抵御也有密切關系[43]。CBF與擬南芥對低溫脅迫的抵御能力有關,PRR9通過調節CBF的表達參與抵御低溫脅迫[44-45]。本研究發現,低溫(15 ℃)脅迫的整個過程SiPRR73基因呈現強誘導表達模式,說明該基因可能參與谷子對低溫脅迫的抵抗作用。PRRs家族基因與ABA調控基因有部分重疊,ABAR、ABI3通過ABA調控表達參與植物抗逆性,這些基因也受PRRs家族基因TOC1的負向調節,同時對TOC1有反向調節作用,通過這種調節環的方式參與ABA信號傳導[46]。SiPRR73基因表達明顯受到ABA調節,且在ABA處理不同時間表達模式不同,表明SiPRR73可能參與ABA介導的信號傳遞途徑。擬南芥PRR5基因參與鹽脅迫調節[47],本研究發現,SiPRR73對NaCl脅迫的響應特點接近ABA,因此可能與鹽脅迫應答有關。水稻OsPRR73的表達受到干旱抑制[2],而本研究發現,谷子SiPRR73基因明顯被PEG模擬的干旱脅迫所誘導,說明水稻與谷子PRR73基因存在不同的干旱脅迫應對機制。植物鐵蛋白具有儲鐵功能,可以緩解鐵過量,擬南芥PRR7抑制鐵蛋白基因的表達,與鐵脅迫應答有關[20,24]。本研究發現,鐵脅迫初期SiPRR73基因的表達受到抑制,說明鐵脅迫需要大量鐵蛋白發揮儲鐵功能,脅迫信號傳遞給SiPRR73基因,使其表達減弱,有利于鐵蛋白基因的表達,而后期SiPRR73表達增強說明隨著脅迫減弱,鐵蛋白需求減少,SiPRR73開始抑制基因表達,因此,推測SiPRR73和擬南芥PRR7一樣,參與了作物抵御鐵脅迫。
本研究首次克隆得到谷子SiPRR73基因包含完整編碼區的cDNA序列,該基因CDS長度2 283 bp,編碼760個氨基酸;SiPRR73基因在谷子次頂葉表達水平最高,受光誘導和光周期調控;光周期和溫度共同調節SiPRR73基因表達,通過改變表達峰出現時間、表達峰數目影響基因表達模式,從而可能參與了谷子對不同光溫環境的適應性調節;SiPRR73基因也參與了谷子對鹽脅迫、低溫脅迫、ABA脅迫、干旱脅迫和鐵脅迫的應答反應。
