植物脅迫相關microRNA研究進展
2022-09-14 04:43:04王楠楠王文佳朱強
生物技術通報 2022年8期
王楠楠 王文佳 朱強
(福建農林大學林學院,福州 350000)
miRNA是一種植物體內廣泛存在的內源性非編碼RNA。1993年,Lee等[1]在秀麗隱桿線蟲中首次發現動物miRNA lin-4,隨后在2002年,Reinhart等[2]在擬南芥中也發現了16條miRNA。隨著高通量測序和生物信息技術的迅速發展,越來越多miRNA被發 現,截 至2019年,miRBase(http://mirbase.org/)已經收錄了來自271個物種的38 598條miRNA前體和48 860條成熟體[3]。近年來,越來越多的研究表明miRNA在植物的諸多生物學過程中都起著重要作用,其中miRNA在植物生長發育過程中的重要作用已經由前人較為詳細的總結[4-5]。本文側重闡述挖掘miRNA在植物脅迫響應過程中的作用。
1 miRNA的產生及作用方式
1.1 miRNA的產生
miRNA的基因(MIR)被RNA聚合酶II(Pol II)轉錄成具有莖環結構的初級miRNA(pri-miRNA)。pri-miRNA在RNase III家族酶DICER-LIKE1(DCL1)的作用下加工形成miRNA/miRNA*雙鏈體。雙鏈體的3'端被甲基轉移酶HUA ENHANCER1(HEN1)2'-O-甲基化。之后,雙鏈體中的一條鏈被整合到ARGONAUTE1(AGO1)中以形成RNA誘導的沉默復合物(RISC)。以下將詳細介紹miRNA復雜的產生過程。
1.1.1MIR轉錄與轉錄調控 Mediator作為一種通用的轉錄共激活因子,有助于將Pol II募集到MIR基因座[6]。還存在一些其他幫助MIR轉錄的蛋白,包括NEGATIVE ON TATA LESS2(NOT2)、含有MYB結構域的DNA結合蛋白CELL DIVISION CYCLE 5(CDC5)和被認為有助于轉錄延伸的Elongator復合物[7-9]。NOT2、CDC5和Elongator都 與Pol II以 及miRNA前體加工復合物相互作用。MIR轉錄中的Pol II活性還受到磷酸化調節[10]。
1.1.2 miRNA前體加工 pri-miRNA由包含DCL1、HYPONASTIC LEAVES1(HYL1)和SERRATE(SE)作為核心成分的切割復合物加工,以產生成熟的miRNA/miRNA*雙鏈體[11]。DCL1起到剪切的作用,將pri-miRNA剪切成21-nt的miRNA/miRNA*雙鏈體,HYL1與DCL1相互作用以促進有效和精確的primiRNA的加工[12]。SE被認為是擬南芥miRNA加工復合體的核心成員,SE突變導致成熟miRNA水平降低、pri-miRNA水平增加以及pri-miRNA剪切缺陷[13]。
1.1.3 miRNA穩定化和RISC形成 miRNA/miRNA*雙鏈體的3'-末端被HEN1 2'-O-甲基化以穩定。HEN1最初是在擬南芥中作為甲基轉移酶被發現的,它可以特異性地甲基化小RNA[14]。擬南芥HEN1-small RNA復合物的晶體結構表明小RNA雙鏈體與HEN1雙鏈RNA結合域(dsRBD)結合,其中一個末端位于甲基轉移酶(MTase)活性位點并依賴于Mg2+甲基化[15]。
隨后,miRNA雙鏈體的一條鏈被選為引導鏈,而另一條鏈則被降解。RISC的形成包括4個部分。第一步,AGO1和HEAT SHOCK PROTEIN 90(HSP90)的二聚體形成復合物。第二步,三磷酸腺苷(ATP)與HSP90的結合引起AGO1構象變化,從而允許雙鏈體結合到AGO1-HSP90蛋白復合物中。第三步,ATP水解誘導HSP90從AGO1解離。最后,HSP90解離引起的AGO1構象變化去除非引導鏈并形成成熟RISC[16]。
目前尚不清楚miRNA是否在RISC形成之前被輸送至細胞質。在早期模型中,miRNA/miRNA*雙鏈體通過HASTY運輸至細胞質,然后裝載到AGO1上[17]。然而,最近提出了另一種模型,即AGO1的裝載發生在細胞核中[18]。
1.2 miRNA的作用方式
植物miRNA通過兩種主要機制在轉錄后水平調節靶基因:mRNA的剪切和翻譯抑制[19]。在植物中,miRNA與靶標mRNA具有近乎完美的互補性,因此,對轉錄本剪切被認為是植物miRNA的主要作用方式。然而,這是一種誤解。雖然高度互補的序列有利于對mRNA的切割,但它也同樣發揮翻譯抑制的作用。事實上,經過實驗驗證,靶基因也會經歷miRNA介導的翻譯抑制[20]。除了mRNA切割和翻譯抑制外,一些miRNA還觸發一些特殊靶基因轉錄本產生次級siRNA(phasiRNAs),成熟的phasiRNA通過堿基互補配對原理在轉錄后水平發揮負向調控作用,這是植物中普遍且保守的現象[21]。
2 miRNA參與脅迫響應
研究表明,miRNA在多種生物學過程中起著重要的調節作用,包括植物的發育、代謝、非生物脅迫以及病原體防御等過程(圖1)。環境脅迫能誘導植物在表型和生理生化上產生一系列的變化以適應其正常生長,例如在表型上調整植株的大小、葉片大小及卷曲程度、根的生長情況、氣孔開合以及細胞大小等;生理生化上表現在次級代謝物積累、光合作用的抑制、呼吸作用的加強以及一些抗氧化酶的激活等。并且為了適應環境,植物也進化出各種抗逆機制,例如脫落酸(abscisic acid,ABA)合成、活性氧(reactive oxygen species,ROS)清除、離子平衡、乙烯反應和調節下游防御反應的絲裂原活化蛋白激酶(MAPK)途徑等。探究miRNA在植物響應環境脅迫過程中的分子機制,有利于未來利用基因工程技術提高植物抗性、改良植物品質。
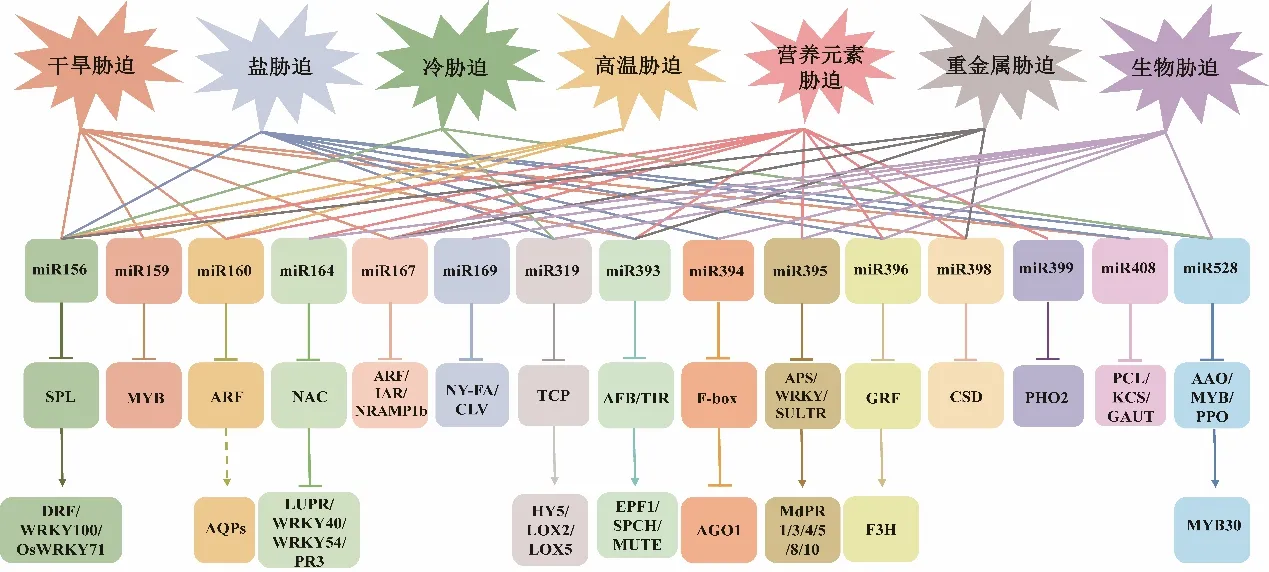
圖1 miRNA在脅迫響應過程中的作用Fig. 1 Role of microRNA in plant stress responses
2.1 非生物脅迫
2.1.1 干旱脅迫 干旱是決定植物生產力和分布的重要環境因素之一。miR156是植物中最早被發現的一種miRNA,它與SQUAMOSA-promoter binding like(SPL)轉錄因子的相互關系在植物中普遍保守。玉米miR156在煙草內過表達能提高轉基因植物的抗旱性,進一步研究表明,抗旱性的提高與脯氨酸等次級代謝物的積累以及抗氧化酶表達量升高有關[22]。番茄miR159也可通過調節MYB33的轉錄水平誘導脯氨酸和腐胺的積累從而幫助植物提高抗旱性[23]。花青素作為一種次級代謝產物通過清除ROS以保護植物免受脅迫。在擬南芥、水稻、紫花苜蓿以及楊樹中,都存在miR156/SPL通過調節花青素的積累水平響應植物干旱脅迫的機制[24-26]。其中,苜蓿SPL13可以直接與編碼花青素生物合成過程關鍵酶基因DIHYDROFLAVONOL-4-REDUCTASE(DFR)的啟動子區域相結合調節其表達。
1.1.1納入標準 (1)有高血壓病史,或者正在服用降壓藥者。(2)明顯心律不齊或者心房顫動者。(3)血脂異常者。(4)吸煙者。(5)無體育運動者。(6)糖尿病者。(7)BMI(身體指數)在26kg/m2以上者。(8)有腦卒中家族史者。
氣孔是植物葉片表皮中的“窗口”,在植物與環境之間的氣體交換中起核心作用,氣孔的開關受環境信號和內源性激素調節進而影響植物對干旱脅迫的反應和耐受性。而ABA作為一種內源性植物激素,是調節水分流失和氣孔開合的關鍵激素。有研究表明Auxin response factors 10(ARF10)作為miR160的靶基因通過影響氣孔開合進而參與番茄干旱脅迫過程,35S∶mSlARF10轉基因番茄表現出葉片變窄、氣孔變大等表型,SlARF10不僅影響ABA合成,而且也影響氣孔發育和水通道蛋白基因(AQPs)表達,這兩者協同作用可以控制番茄葉片水分的流失[27]。miR393-OE(miR393 overexpressing transgenic line)表現出氣孔密度的增加以及保衛細胞長度的減少,而缺失型突變體表現出相反的表型,這可能與miR393靶基因ARF5以及氣孔發育相關基因EPIDERMAL PATTERNING FACTOR 1(EPF1)、SPEECHLESS(SPCH)以及MUTE的沉默有關。miR393-OE對干旱處理更為敏感,能積累多于野生型的丙二醛(malondialdehyde,MDA)和過氧化氫(H2O2),同時還能抑制ABA在葉片的積累。這些結果也證明了miR393通過與ABA之間的相互作用以及調控氣孔密度從而響應植物干旱脅迫[28]。Zhao等[29]構建的過表達Osa-miR393a的轉基因匍匐翦股穎表現出與氣孔密度和表皮致密相關的干旱脅迫耐受性的增強,還確定了miR393的兩個靶標Auxin signalling F-BOX 2(AsAFB2)和TRANSPORT INHIBITOR RESPONSE 1(AsTIR1)。獨腳金內酯是植物地上部分適應干旱脅迫所需的一種植物激素。對番茄施加外源的獨腳金內酯誘導miR156的積累,miR156-OE和獨腳金內酯的處理都會使植物氣孔導度降低以及ABA敏感性增加[30]。而miR398c卻負向調控大豆的抗旱性,過表達miR398c通過下調過氧化物酶體相關基因(GmCSD1a/b、GmCSD2a/b/c和GmCCS)的表達削弱其清除ROS的能力,引起電解質泄漏和氣孔打開[31]。
植物通過表型的改變以適應干旱脅迫,例如改變根構型以更好的吸收土壤中的水分、調整葉片大小及卷曲程度以減輕水分蒸發等。研究人員在擬南芥中鑒定出miR167的新靶點IAA-Ala Resistant 3(IAR3),IAR3具有將非活性形式的生長素水解并釋放出活性形式的功能。干旱脅迫下miR167表達量下降導致IAR3積累從而提高生長素(auxin)水平以促進側根生成[32]。Yang等[33]在擬南芥中研究了miR160和miR165/166之間的相互作用及其對擬南芥葉片發育和耐旱性的影響。發現STTM160蓮座葉呈現明顯的鋸齒狀,STTM165/166呈現出嚴重的向上卷曲,而STTM160×165/166呈現出輕微的向下卷曲。研究結果還顯示,miR160指導ARF通過生長素信號途徑促進葉片發育,而miR165/166介導的HD-ZIP IIIs通過ABA信號途徑賦予植物耐旱性。在干旱脅迫下,異源表達OsmiR408的轉基因黑麥草與野生型相比,表現出葉片卷曲、氣孔凹陷等表型,這與其保持較高的葉片相對含水量、較低的電解質滲透率和較少的脂質氧化相關,使轉基因植株表現出更強的抗旱能力[34]。
2.1.2 鹽脅迫 土壤鹽堿化是植物面臨的一種非常普遍的非生物脅迫,它影響了世界上約6%的土地和23%的耕地。鹽堿化嚴重影響了作物的生存環境和產量,造成了相當大的經濟損失。植物受到鹽脅迫會顯著影響多種內源性信號分子的水平,包括ABA、乙烯、赤霉素(gibberellin,GA)、ROS、一氧化氮(NO)等。這些信號分子及其下游信號成分已被證明在植物鹽脅迫響應中發揮著重要作用。擬南芥miR394及其靶基因LEAF CURLING RESPONSIVENESS(LCR)將ABA與鹽脅迫響應聯系起來。miR394受ABA誘導而積累,LCR則相反。鹽脅迫實驗表明miR394作為植物鹽脅迫響應的負調節因子,過表達促進ABA引起的氣孔關閉、ROS的積累以及ABA響應基因的表達[35]。過表達Sm-MIR408的轉基因煙草則能在促進種子萌發的同時減少鹽脅迫下ROS的積累,并且增強抗氧化基因的轉錄水平及其酶活性[36]。乙烯信號途徑主要通過調節ROS產生和ROS清除機制來響應鹽脅迫反應。Liu等[37]在柳枝稷耐鹽性實驗研究中發現,miR319-OE轉基因柳枝稷通過抑制靶基因PvPCF5的表達進而下調蛋氨酸循環中關鍵基因的表達,從而促進乙烯合成相關基因表達以提高耐鹽性。
在研究匍匐剪股穎植物響應鹽脅迫的分子機制時發現,過表達Osa-miR528的轉基因匍匐翦股穎節間縮短、分蘗數增加,通過改善植物保水性、細胞膜完整性、葉綠素含量,維持鉀離子穩態、過氧化氫酶活性以及降低抗壞血酸氧化酶(ascorbic acid oxidase,AAO)活性而提高植物對鹽脅迫的耐受性[38]。過表達Osa-miR393a的轉基因匍匐翦股穎通過正向調控離子轉運相關基因(AsNHX1)的表達,促進轉基因植物鉀離子的吸收以達到耐鹽性增強的作用[29]。Yuan等[39]對miR396與耐鹽性之間機制進行研究時發現,Osa-miR396c轉基因植物與野生型相比,表現出生物量減少、節間減短、葉片面積減少、葉脈數量減少、單位面積表皮細胞減少的表型,而在高鹽脅迫下表現出保水性增強、葉綠素含量增加、細胞膜完整性高和鈉離子外排增強等耐鹽性等表型。WRKY轉錄因子通過參與ABA合成、ROS清除、離子平衡和乙烯反應途徑參與鹽脅迫耐受相關的生物學途徑以保護植物免受鹽脅迫損害。然而關于WRKY上游的調控機制尚未可知。近期在蘋果鹽脅迫與miR156的研究中發現,miR156-OE減弱植物耐鹽性,其靶基因SPL13的過表達能增強耐鹽性,同時揭示了SPL13直接與WRKY100相互作用以增強植物耐鹽性[40]。
2.1.3 低溫脅迫 低溫脅迫通過影響光合作用、呼吸作用以及ROS的產生和積累而對植物產生一系列負面作用。miR319已被證明可靶向TEOSINTE BRANCHED/CYCLOIDEA/PCF(TCP)轉 錄 因 子,TCP基因家族是高等植物特有的轉錄因子,這類轉錄因子通過控制細胞增殖以參與調節植物生長發育的多個過程,它們的特征是在N末端具有高度保守的59個氨基酸構成的TCP結構域。OsamiR319的表達受低溫誘導下調,其靶基因OsPCF6和OsTCP21則相反。Osa-miR319過表達導致植物對低溫脅迫耐受性增強,這可能與脯氨酸的積累、OsPCF6和OsTCP21調 節 的ROS的 清 除 以 及以一些低溫響應基因表達量升高有關[41]。在甜瓜中,低溫誘導miR319c表達量降低,引起TCP2介導的ELONGATED HIPOCOTYL 5(HY5)水平增加,HY5轉錄因子通過調節花青素生物合成途徑中關鍵成分積累從而響應低溫脅迫[42]。番茄miR319d過表達也增強其冷脅迫耐受性,具體表現在電解質滲透率、MDA濃度、O2-濃度和H2O2濃度的降低以及葉綠素含量和Fv/Fm值的增高,這可能與FeSOD、CuZnSOD和過氧化氫酶(CAT)基因表達水平升高以及GAMYB-like1表達水平降低有關[43]。
OsmiR528過表達增強了擬南芥、松樹和水稻的冷脅迫耐受性。OsmiR528通過靶向F-box進而降低轉錄因子OsMYB30的表達。據報道,OsMYB30促 進β淀 粉 酶(β-amylase,BMY)基 因OsBMY2、OsBMY6和OsBMY10表達從而增加細胞中糖含量以增強植物冷脅迫耐受性[44]。香蕉miR528在冷脅迫下顯著下調,其靶基因多酚氧化酶(Polyphenol oxidase,PPO)基因的表達量則上調100倍以上,這引起ROS含量激增,使得香蕉果實果皮褐變[45]。水稻miRNA156過表達導致擬南芥、松樹和水稻在冷脅迫下細胞活力和生長速率增加。OsmiR156通過靶向OsSPL3增加植物耐寒性,OsSPL3正向調節OsWRKY71的表達,而OsWRKY71是轉錄因子OsMYB2和OsMYB3R-2的 負 調 節 因 子。OsMYB2和OsMYB3R-2分別通過激活OsLEA3、OsRab16A、OsDREB2A和OsKNOLLE2、OsCTP1、OsCycB1.1、OsCycB2.1和OsCDC20.1表達來抵消低溫脅迫所帶來的負面影響[46]。
2.1.4 高溫脅迫 高溫作為一個重要的環境因素,會對植物生長、發育、繁殖等過程產生負面影響。植物響應高溫脅迫是一個復雜的過程,涉及各種代謝和生化過程。苜蓿中的相關研究證明miR156參與植物熱脅迫響應過程,mi156-OE轉基因植物通過抑制靶基因SPL13從而增強其耐熱性[47]。Arshad等[48]測量了熱脅迫下野生型和miR156-OE轉基因植物的各項生理參數,并收集了葉組織進行蛋白組分析,發現miR156-OE植物中的脯氨酸和抗氧化劑含量高于野生型,miR156-OE植物特有的蛋白質與多種功能有關,包括代謝、光合作用、應激反應和植物防御。
miR159/MYB在小麥和白菜的熱應激過程中都起重要作用,小麥miR159過表達與靶基因GAMYB表達的下調相結合會導致其耐熱性的降低,但單獨過表達GAMYB卻沒有提高幼苗的耐熱性,所以,miR159/GAMYB途徑與熱應激耐受性之間的因果關系還需要進一步研究[49-50]。此外,過表達miR160可以增加棉花對高溫脅迫的敏感性,這與ARF10和ARF17表達受到抑制從而激活導致花藥開裂的Auxin信號通路有關[51]。
2.1.5 養分脅迫 氮是植物中許多重要化合物的主要成分,參與植物一系列生化反應,并在作物的生物量積累和產量提升中發揮重要作用。鉀則參與滲透調節、光合作用、物質運輸等過程,可以提高植物的抗逆性。因此,缺乏這兩種元素會導致植物生長受到限制。對花生的營養缺乏機制研究中發現,氮和鉀缺乏會影響植物的生長,使植物表現出地上和地下部分組織干重降低,根長、根表面積、根體積、根活力降低和根呼吸減弱。氮缺乏顯著降低初生根長度和側根數量,這可能與miR160、miR167、miR393和miR396的上調以及AFB3和GRF的下調有關。主根和側根對缺鉀的反應與缺氮條件相反,表現為miR156、miR390、NAMATAF-CUC 4(NAC4)、ARF2和AFB3的 上 調,以 及miR160、miR164、miR393和SPL10的下調,這些可能有助于缺鉀條件下初生根和側根的生長[52]。
磷是植物不可或缺的一種常量營養元素,在發育和代謝過程中都很重要。雖然土壤中磷含量豐富,但大多數土壤中磷與鈣、鐵和鋁等元素結合形成不溶性的化合物,限制了植物對磷的獲取。缺磷是植物生長和發育過程中的常見脅迫。miR156/SPL3在擬南芥磷吸收過程中發揮著重要作用,過表達miR156的轉基因植株與野生型相比磷吸收速度加快,并在缺磷狀態下能減少根際酸化和花青素積累[53]。據報道,miR399是磷饑餓信號途徑中的重要組分。最早在擬南芥中闡明miR399在磷饑餓信號傳導中的功能,在磷饑餓條件下,miR399的表達被顯著提高,而靶基因ubiquitin-conjugating E2 enzyme(PHO2)表達降低。為進一步研究miR399的功能,Hu等[54]對OsmiR399過表達植物進行了基因芯片分析發現,除了磷饑餓反應基因外,許多參與鐵、鉀、鈉和鈣吸收的基因也被顯著上調,鐵、鉀、鈉和鈣的濃度也有所增加。此外,OsmiR399下游靶標LTN1(OsPHO2)的功能喪失也導致這些營養元素濃度增加以及相關基因的上調。miR399-PHO2途徑參與植物對磷缺乏的適應性反應同樣在玉米中存在[55]。
硫元素參與植物發育的許多重要機制和途徑。土壤中的硫元素主要以硫酸鹽(SO4-)形式存在并被吸收入根組織,通過脈管系統輸送至地上部分,被還原成亞硫酸鹽后在葉綠體和線粒體等亞細胞器官中硫化,用于半胱氨酸和蛋氨酸生物合成。而空氣中的SO2是一種常見的污染物,在植物中,SO2主要通過葉組織氣孔吸收,然后水合形成亞硫酸鹽和亞硫酸氫鹽。Yuan等[56]發現Osa-miR395的轉錄水平在硫酸鹽缺乏條件下增加,這與前人在擬南芥中的研究結果相同。將Osa-miR395h在煙草中異源過表達損害了煙草硫酸鹽穩態,NtaSULTR2作為miR395的靶點,屬于低親和力硫酸鹽轉運蛋白。Li等[57]發現擬南芥暴露在SO2中,miR398的表達下調,而靶基因銅或鋅超氧化物歧化酶(Cu or Zn superoxide dismutases,CSD)的轉錄水平增加,同時SOD活性隨著CSD轉錄水平的增加而增強,表明miR398在響應SO2誘導的氧化應激中起重要作用。然而,miR395的表達受SO2影響增加,下游靶基因APS3和APS4以及SULTR2;1的轉錄水平在擬南芥中降低,表明miR395在SO2暴露期間硫酸鹽同化和易位的調節中發揮了重要作用。
2.1.6 重金屬脅迫 重金屬元素被認為是植物的生長和產量的主要限制因素,包括必需金屬元素和非必需金屬元素。多種生理過程需要必需金屬元素,例如鋅、錳和銅等,其他一些金屬,如鎘、鉛或汞則是非必需金屬元素。植物重金屬中毒的顯著特征是酶反應的中斷和氧化應激。天然抗性相關巨噬細胞蛋白(natural resistance-associated macrophage proteins,NRAMP)是植物中不可或缺的膜轉運蛋白,負責轉運必需金屬元素如錳或鐵以及非必需金屬如鎘或鉛。在甘藍型油菜中發現了一個組成型基因BnNRAMP1b,它受鎘脅迫而上調。結合降解組相關信息驗證了BnNRAMP1b作為miR167的靶基因參與金屬脅迫響應[58]。過表達miR156的轉基因擬南芥減少地上部分重金屬鎘積累,增強了植物對鎘脅迫的耐受性。在鎘脅迫下,miR156OE具有初生根增長、生物量和葉綠素含量增高、抗氧化酶活性增強、ROS水平降低以及鎘轉運相關基因表達量降低的表現,而MIM156則相反[59]。
miR393在植物根組織鋁中毒脅迫中具有一定的生物學功能。土壤中鋁離子抑制大麥根尖miR393的表達,miR393的過表達可以消除鋁離子對根伸長的抑制以及ROS積累所導致的細胞死亡。此外,miR393過表達減弱了外源生長素對鋁誘導的根生長抑制的影響,并下調了鋁脅迫下生長素響應基因的表達,這意味著miR393是通過改變大麥中的生長素信號輸出來調節根對鋁的敏感性[60]。
葡萄miR398通過調節靶基因VvCSD1和VvCSD2的表達,調控其對銅脅迫的響應過程。Vv-miR398的表達受到不同濃度銅脅迫抑制,而VvCSD1和VvCSD2基因的表達增強。與野生型相比,VvCSD過表達轉基因植株具有更高的銅耐受性,并且在高濃度銅脅迫條件下降低ROS積累、提高SOD活性,這表明VvCSD增強ROS清除系統從而保護植物免受更多氧化損傷[61]。
2.2 生物脅迫
植物病原體包括害蟲、真菌、細菌以及病毒,它們嚴重威脅植物的正常發育。植物免疫系統通過上調或下調miRNA的表達來誘導下游轉錄因子和基因表達以抵抗病原體的入侵。Caruana等[62]發現miR167通過ARF6和ARF8調節植物對病原體的防御反應。miR167響應細菌病原體假單胞桿菌而差異表達,其過表達賦予植物更高的抗菌性。這種抗性與生長素信號轉導、水楊酸信號傳導及氣孔運動有關。此外,miR167-OE轉基因植物的系統性獲得抗性(systemic acquired resistance,SAR)反應也受到嚴重損害,進一步研究表明SAR的激活需要完整的生長素信號轉導過程。總之,miR167在激素平衡以及植物生物防御過程中起著重要作用。
miR169在植物生物脅迫過程同樣發揮著重要的作用。擬南芥miR169靶基因CLAVATA1(CLV1)和CLAVATA2(CLV2)的突變賦予植物更強的抗病性,而miR169的過表達能夠消除clv突變體的抗病性表型[63]。水稻miR169過表達植株對病菌的侵染高度敏感,這與防御相關基因表達量的降低以及感染部位缺乏過氧化氫積累有關。此外,miR169通過抑制核轉錄因子Y(Nuclear factor Y,NF-YA)基因的表達而在對病菌的免疫過程中起負向調節因子的作用[64]。然而,香蕉miR169a和miR169b表達量在被病原體侵染后上調并在香蕉抗性品種中保持較高表達水平[65]。
NAC4作為miR164的靶基因之一,在擬南芥響應生物脅迫的細胞程序性死亡中發揮作用。研究表明NAC4過表達體和miR164沉默型植株對細菌病原體的過敏反應更加強烈,細胞死亡癥狀增強,而NAC4的 下 游 基因LURP1、WRKY40和WRKY54在細胞程序性死亡過程充當負向調節因子[66]。Wang等[67]發現,過表達miR164的靶基因OsNAC60增加細胞程序性死亡、加快離子泄漏、增強ROS積累、胼胝質沉積以及防御相關基因的上調,而增強水稻對稻瘟癥的抵御能力。在上述研究中,miR164在植物生物脅迫過程中發揮消極作用,然而,在最新研究中也發現其具有積極調控作用。Hu等[68]在對棉花黃萎病的研究中發現,ghr-miR164作為正向調節因子可提高棉花對黃萎病的抗性,沉默其靶基因GhNAC100也可增強抗病性。還發現GhNAC100與GhPR3啟動子的CGTA box結合抑制其表達,從而減弱植物抗病性。此外,miR164也在小麥的葉銹病以及楊樹黑斑病防御過程中發揮重要作用[69-70]。
miR319在生物脅迫過程也起重要作用。番茄miR319-TCP4通過影響茉莉酸(jasmonic Acid,JA)合成基因表達和葉片內源JA水平,進而影響植物對根結線蟲的抗性[71]。Zhang等[72]發現米曲霉菌株感染能特異性誘導水稻miR319的表達,抑制其靶基因OsTCP21表達。當miR319b-OE水稻受米曲霉菌感染時,LIPOXYGENASE2(LOX2)和LOX5作為JA合成過程中的關鍵酶被抑制表達[72]。Fan等[73]報告了miR319a/TCP參與毛白楊GA信號轉導以及毛狀體形成過程。miR319a過表達降低靶向的TCP轉錄水平,顯著提高轉基因楊樹的葉毛密度,從而減輕昆蟲危害性。
大蒜受病原體侵害誘導miR394表達而下調F-box和cytochrome P450(CYP450)基 因,因 此miR394及其對靶基因調控強度的差異可作為選擇抗病大蒜品種的標志[74]。miR394作為擬南芥應對細菌感染的負調控因子,其過表達通過影響LCR以及許多參與植物miRNA代謝的關鍵基因AGO1表達從而提高植物對細菌的易感性[75]。Zhang等[76]在番茄中的研究也驗證了miR394參與負向調控生物脅迫,miR394的過表達抑制其靶基因LCR的表達,進而抑制JA合成相關基因,從而降低番茄對致病疫霉的抗性。
在研究miR396在植物生物脅迫過程中作用時發現,miR396表達量降低賦予植物對真菌病原體的廣泛抗性[77]。水稻過表達miR396靶基因Growth Regulating Factor(GRF)的轉基因植物顯示出更強的抗菌性,表明miR396通過抑制GRF而負向調控水稻對細菌侵染的抗性[78]。Chandran等[78]證明過表達miR396轉基因株系對米曲霉高度敏感,而過表達其靶基因OsGRF6、OsGRF7、OsGRF8和OsGRF9的轉基因植物對米曲霉的抗性增強。進一步研究顯示,MIM miR396和GRF8-OE植物中類黃酮含量均增加,黃酮含量的增加與病原體抗性增強相關,這揭示了miR396-GRF8-F3H-類黃酮途徑介導的新的病原體抗性機制[79]。
Zhang等[80]在對蘋果抗菌性研究中發現,接種葉斑病真菌的葉片上,MdmiR395表達水平升高而其靶基因MdMRKY26、MdWRKYN1表達降低。過表達MdWRKY或抑制MdmiR395可以增加與抗病相關(pathogenesis-related,PR)基因的表達從而增強蘋果的抗病性。Wu等[81]發現,當水稻遭受病毒危害時,miR528表達受抑制,這促進其靶基因L-抗壞血酸氧化酶(L-ascorbate oxidase,AO)表達從而減少AO介導的活性氧的積累。在之后的研究中又發現miR528-AO防御模塊受SPL9的調控,SPL9與miR528啟動子區域內特定基序結合從而激活miR528基因的表達[82]。
3 總結與展望
環境脅迫是影響植物正常生長發育的重要因素之一,不同的環境脅迫會誘導miRNA的差異性表達。miRNA作為調節植物脅迫響應的調控網絡中心,其一舉一動都會引起“蝴蝶效應”,因此對miRNA的研究有助于改良植物的遺傳性狀,提高其抗逆性。研究表明即使是同種miRNA在不同物種中對脅迫響應表現不同。例如,不同物種miR156在鹽脅迫下的表達模式不同,蘋果、玉米相似,擬南芥、甘蔗和水稻相似[24,40,83-85]。木本植物和草本植物在生命周期方面的差異可能解釋了這種現象。一年生草本植物的生命周期比多年生木本植物短,所以當草本植物受到鹽脅迫時,它們會將能量分配給防御機制,于是引起miR156積累抑制SPL表達,從而延長植物的幼年期,使其增強對不利環境條件的耐受性。與擬南芥和其他草本植物不同,多年生木本植物的幼年期較長,其與耐鹽性之間的關系可能比草本植物更復雜。當蘋果受到鹽脅迫時,它通過下調miR156以上調下游靶基因SPL。某些SPL可增強側根伸長和生物量積累,以抵消鹽脅迫引起的能量消耗。其他SPL通過調控下游基因表達以調節植物生理和代謝過程以響應鹽脅迫。除此之外,miR164、miR169在不同物種的生物脅迫方面也起著不同的調控作用[64-68]。即便依據miRNA片段小以及遺傳冗余等特點,研究人員已經開發了Target mimicry(MIM)、Short tandem target mimic(STTM)技術等,但是大量的研究結果顯示,miRNA具有組織和器官特異性。例如miR159在不同組織的表達量不同[86-88]。但目前尚沒有確切的機制解釋這些現象。未來對于單個miRNA的通路研究將可能說明這一現象,這將會成為未來研究趨勢之一,其中單細胞測序技術可能在其中發揮重要作用。
隨著二代測序技術以及miRNA研究技術的不斷發展,越來越多新的miRNA被發現。然而目前的研究大多僅局限于miRNA及其靶基因在生物學過程中的作用,而miRNA是如何接收到上級信號以及通過級聯反應影響下游各種調控途徑的分子機制尚不清楚。因此,建立更為系統的miRNA調控網絡將會成為未來另一個趨勢。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13