利用CRISPR/Cas 9編輯紅曲霉pyrG基因對其次生代謝的影響
2022-09-14 04:44:16唐光甫桂艷玲滿海喬趙杰宏
生物技術通報 2022年8期
唐光甫 桂艷玲 滿海喬 趙杰宏
(貴州中醫藥大學 貴州省中藥生藥學重點實驗室,貴陽 550025)
紅曲霉(Monascus)作為我國傳統藥食兩用真菌,已有上千年歷史[1],目前常利用紅曲霉發酵生產多種食品,如娃哈哈紅曲藜麥餅干、茅臺不老紅曲酒、紅曲腐乳和紅曲腌漬魚等,紅曲色素也被廣泛用作食品添加劑。現代科學證明紅曲霉能生產麥角固醇、洛伐他汀、γ-氨基丁酸等多種功能活性物質[2],已有脂必妥、血脂康等藥品和納豆紅曲、靈芝紅曲等保健品的上市銷售。但自從紅曲產品中檢測出桔霉素這種真菌毒素[3]以來,紅曲產品的生產和應用受到很大影響[4]。因此深入研究紅曲霉次生代謝產物的遺傳調控、加快選育安全、高效的紅曲霉菌種非常重要。
已報道通過同源重組技術先后開展了CntA/Orf2[5]、CtnB/Orf4[6]、CtnE[7]、CtnF[8]、CtnG[9]、CtnH[10]、Orf1[11]、Orf3[12]、Orf6[13]、Orf7[14]和pksCT[15]等基因的敲除實驗,初步證實了多個桔霉素合成相關基因。有研究報道尿嘧啶核苷的天然產物能夠成為新型小分子的抑制劑[16],而黑曲霉pyrG基因(乳清酸核苷-5'-磷酸脫羧酶基因)的破壞能增強糖酵解通量,并顯著提高檸檬酸鹽及其前體的量[17],可見尿嘧啶核苷及其合成關鍵基因pyrG在真菌物質代謝上有重要功能。本文通過構建紅曲霉Cas 9底盤菌株,利用體外轉錄合成的sgRNA轉化其原生質體實現對pyrG基因的定向編輯,為紅曲霉構建了快速基因編輯體系,并證實pyrG基因對紅曲霉的次生代謝有重要影響。
1 材料與方法
1.1 材料
1.1.1 菌株 紫色紅曲霉(Monascus purpureus)菌株由貴州中醫藥大學生藥學實驗室提供,農桿菌(Agrobacterium tumefaciens)EHA 105和 大 腸 桿 菌(Escherichia coli)TOP 10感受態(購自北京華越洋)。
1.1.2 試劑 pGM-T克隆試劑盒和Genegreen核酸染料(購自北京天根生化),sgRNA體外轉錄試劑盒(購自重慶英茂盛業),T7EI核酸內切酶(購自北京唯尚立德),尿嘧啶核苷和5-氟乳清酸(5-FOA)(購自上海生工),纖維素酶、溶壁酶和蝸牛酶(購自北京索萊寶),pDHt/sk-PC(購自Addgene),桔霉素Elisa檢測試劑盒(購自上海紀寧)。
1.2 方法
1.2.1 CRISPR/Cas9遺傳轉化 將pDHt/sk-PC(含Ppdc-toCas9-Tpdc表達框)質粒經凍融法轉化農桿菌EHA 105感受態,涂布在含50 mg/L Kan的LB培養基上篩選,使用Cas 9檢測引物(表1)對抗性菌落分別PCR鑒定,將獲得的農桿菌陽性菌株在含50 mg/L kan液體LB培養基中培養,取菌液離心去除上清,用IM培養基稀釋至OD600值為0.15-0.2,再在相同條件下培養4-6 h,至其OD600值為0.6-0.8。用此菌液稀釋紅曲霉孢子至106個/mL,取200 μL涂布于鋪有0.45 μm硝酸纖維素膜的Co-IM培養基[18]平板上,25℃暗處共培養3 d,將長出的菌落和硝酸纖維素膜一起揭下倒扣在含20 mg/L潮霉素和300 mg/L頭孢霉素的沙氏培養基上篩選,2 d后揭下硝酸纖維素膜,挑取抗性紅曲霉單菌落使用Cas 9檢測引物分別進行PCR分析。

表1 PCR檢測引物和sgRNA序列Table 1 Primers and sgRNA sequences for PCR detection
1.2.2 原生質體制備 將紅曲霉底盤菌株接種在含潮霉素的PDA平板上培養15 d后,用無菌水沖洗孢子,制成1×108個/mL的孢子懸液涂布于鋪玻璃紙的PDA平板上,40 h后刮取菌絲,按比例(M∶V=1∶30)加入酶裂解液[19](用1.0 mol/L MgSO4配制,含1.0 g/L纖維素酶、3.0 g/L的溶壁酶和10.0 g/L的蝸牛酶),置于60 r/min和30℃搖床中酶解2.5-3 h,然后在顯微鏡下統計原生質體的數量。
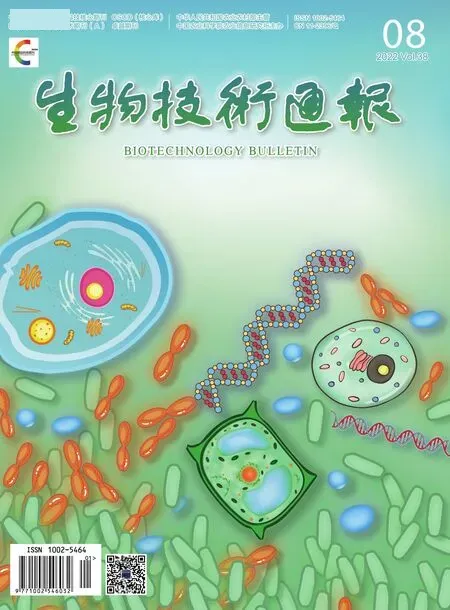
1.2.3 sgRNA合成和轉化 將橙色紅曲pyrG基因(GenBank:GU723506.1)序列在NCBI 與紫色紅曲基因組blast,將共有序列提交到http://crispor.tefor.net/在線設計pyrG的sgRNA(圖1)及其檢測引物對(表1)送北京六合華大基因公司合成。利用體外轉錄試劑盒合成sgRNA,并經電泳檢測。取100 μL原生質體加入10 μL sgRNA,冰浴30 min后加入1 mL PTC溶 液(PEG 4000 40%、pH 8.0 Tris 50 mmol/L、CaCl250 mmol/L),室溫放置40-60 min,取300 μL涂布在含20 mg/L潮霉素、20 g/L尿嘧啶核苷和1.5 g/L 5-FOA的CM再生培養基(蔗糖210.38 g/L、酵母膏6 g/L、酪氨酸6 g/L、瓊脂20 g/L)上30℃恒溫培養4-7 d。利用Cas 9引物對長出的抗性紅曲霉菌株進行PCR擴增、T7EI酶切和測序鑒定。
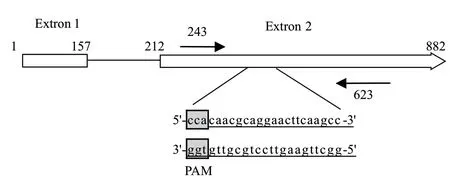
圖1 pyrG基因sgRNA靶標序列Fig. 1 Target sequence of sgRNA for gene pyrG
1.2.4 化學成分測定 將液體沙氏培養基二級接種培養14 d后的菌絲體過濾,45℃干燥后研碎。測定紅曲色素,精密稱取0.10 g菌絲粉末加入10 mL 70%乙醇搖勻,60℃水浴1 h,然后補足10 mL濾過,將濾液倒入25 mL容量瓶中定容,用紫外分光光度計檢測505 nm吸光度,計算紅曲色素的色價(色價U/g=吸光度×稀釋倍數×浸提溶劑體積/樣品質量)。測定洛伐他汀,取1 mL發酵液加入9 mL 70%乙醇混勻,放入恒溫振蕩器28℃、130 r/min振蕩1 h,然后4 200 r/min離心5 min,取上清液檢測237 nm處吸光度值來判斷含量高低[20]。測定桔霉素,稱取菌絲體粉末0.30 g,加入2 mL 70%甲醇超聲10 min后,取1 mL懸濁液12 000 r/min離心10 min,取上清液用桔霉素 ELISA試劑盒進行測定,同步制作桔霉素標準曲線(圖2)。

圖2 桔霉素標準曲線Fig. 2 Standard curve of citrinin
1.2.5 數據統計及顯著性分析 實驗數據以Graghpad prism軟件分析,以“平均值±標準偏差”(Mean±SD)表示。分析方法為“單因素方差分析-多重比較-Turkey檢驗”,*P<0.05表示有顯著性差異,**P<0.01有較顯著差異,***P<0.005有極顯著差異。
2 結果
2.1 制備紅曲霉Cas 9底盤菌株
將含Ppdc-toCas9-Tpdc表達框的pDHt/sk-PC質粒經凍融法轉化農桿菌EHA 105感受態,對Kan平板上的抗性農桿菌菌落PCR檢測(圖3-A),獲得陽性轉化菌株,然后與紫色紅曲霉孢子懸液共培養(圖3-B),再轉接到添加Hyg和Cef的平板上進行抗性篩選,使用Cas 9引物對抗性紅曲霉菌落進行PCR檢測(圖3-C),獲得了轉化成功的紅曲霉Cas 9底盤菌株。
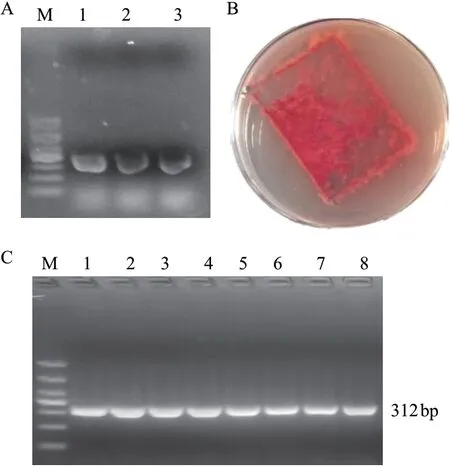
圖3 農桿菌介導Cas 9轉化紅曲霉Fig. 3 Transformation of Cas 9 into monascus mediated by A. tumefaciens
2.2 體外sgRNA轉化原生質體
基因pyrG是否變異可以通過在添加5-FOA和尿嘧啶核苷的培養基上篩選,能夠正常生長的菌株,推測其pyrG基因發生突變并喪失功能,因此需建立5-FOA對紫色紅曲霉的最佳篩選濃度。通過比較不同濃度5-FOA對紅曲霉生長的抑制作用,發現當5-FOA濃度達到1.5 g/L時,紅曲霉完全被抑制(圖4-A),故選擇1.5 g/L的5-FOA作為紅曲霉篩選濃度[21]。使用體外轉錄試劑盒合成pyrG基因的sgRNA,電泳檢測表明條帶大小正確(圖4-B),經PEG介導將sgRNA轉化到紅曲霉原生質體中(圖4-C),在含20 mg/L潮霉素、20 g/L尿嘧啶核苷和1.5 g/L 5-FOA的培養基上篩選,獲得了抗性紅曲霉菌株(圖4-D)。
圖4 體外sgRNA轉化紅曲霉原生質體Fig. 4 Transformation of monascus protoplasts by sgRNA in vitro
2.3 突變株pyrG基因變異分析
用pyrG引物對獲得的抗性紅曲霉菌株PCR擴增,部分PCR產物的序列有明顯的局部套峰(圖5-A),說明序列上存在變異導致錯峰。雖然5、6、7、8號菌株PCR產物是單條帶(圖5-B),但用T7EI核酸內切酶檢測pyrG序列與野生型的雜合性時,結果顯示5號和6號在突變株泳道及混合泳道上均切出了第2條帶,可見5號和6號菌株有變異位點,屬于雜合基因型;7號菌株僅在混合泳道切出了第2條帶,屬于純合基因型(圖5-C)。經測序證實,pyrG基因序列被CRISRP/Cas9成功編輯。4個突變菌株中,5號和8號在相同位點均缺失2個堿基,6號和7號在臨近位點各缺失1個堿基,變異均發生在PAM位點上游2-5 bp范圍(圖5-D)。
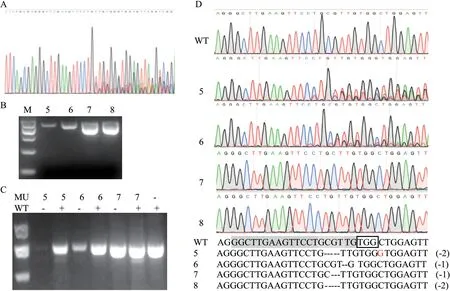
圖5 紅曲霉突變株pyrG基因的變異分析Fig. 5 Variation analysis of pyrG gene of monascus mutant
2.4 突變株的形態特征
通過CRISPR/Cas 9技術把pyrG基因編輯成功的紅曲霉菌株接種在含潮霉素、5-FOA和尿嘧啶核苷的培養基上能夠生長,而野生型不能生長,可見突變株的pyrG基因失去了功能,不受5-FOA的影響,產生對尿嘧啶核苷的營養依賴性。對突變株的菌落形態、顯微結構(400×)和生長速率進行觀察,除了pyrG突變株菌體的正面顏色略有變淡外,菌落、分生孢子和閉囊殼的形態特征沒有較明顯的變化(圖6),說明pyrG基因變異未導致菌株出現很明顯的生長發育差異甚至畸形,推測pyrG基因可能對紅曲霉的次生代謝有影響。

圖6 部分突變株的菌落形態和顯微特征Fig. 6 Colony morphology and microscopic characteristics of partial mutant strains
2.5 突變株化學成分的含量變化
比較突變株與野生型主要化學成分的含量,結果顯示(圖7),培養基中添加尿嘧啶核苷的野生型比不添加的對照菌株中洛伐他汀和紅曲色素含量極顯著增加,桔霉素含量極顯著減少。可見,尿嘧啶核苷對紅曲霉的次生代謝有重要影響。當在添加尿嘧啶核苷的培養條件下比較突變株與野生型菌株,結果顯示,洛伐他汀含量在9個變異株中均有增加(圖7-A);紅曲色素含量在4個變異株中有增加,在5個菌株中不同程度減少,增減比較隨機(圖7-B);桔霉素在7個變異株中顯著增加,另2個變異株的桔霉素含量未增反降(圖7-C)。因為洛伐他汀、紅曲色素和桔霉素皆屬于聚酮類化合物,其主干代謝途徑較一致,代謝流之間相互關聯,因此pyrG基因變異造成的影響比較復雜。
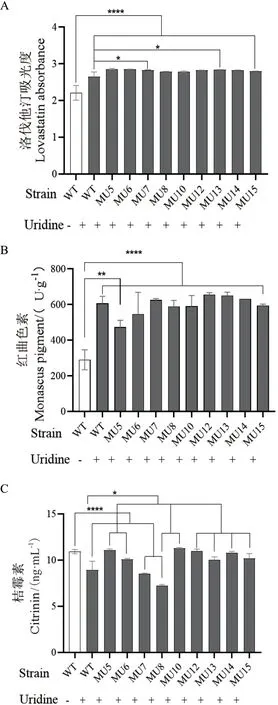
圖7 培養基中添加尿嘧啶核苷對不同菌株化學成分含量的影響Fig. 7 Effect of uracil riboside on the chemical composition content of different strains in medium
3 討論
CRISPR/Cas 9是一種基因靶向修飾技術,通過sgRNA與靶向基因序列結合,引導Cas 9核酸內切酶對靶序列進行切割,然后細胞在DNA修復過程中產生的InDel導致目的基因移碼和功能缺失[22]。該技術作為一種簡單、快速的基因編輯工具[23],主要通過Cas 9核酸酶和靶向sgRNA共同發揮作用,一般需要針對不同靶向序列設計和構建Cas 9和不同sgRNA的共表達載體,再對真菌遺傳轉化才能實現基因的定向編輯,導致基因功能研究耗時費力。如果有表達Cas 9的底盤菌株,研究基因功能時僅需導入不同的sgRNA即可實現各種基因的編輯目的,從而簡化實驗。
本研究通過農桿菌介導遺傳轉化獲得了Cas 9組成型表達的紅曲霉底盤菌株,再把人工合成的pyrG基因靶向sgRNA通過PEG介導法轉化Cas 9底盤菌株原生質體,構建了體外sgRNA對紅曲霉Cas 9底盤菌株中pyrG基因的定向編輯。pyrG基因是尿嘧啶核苷合成中的關鍵酶,能把5-FOA轉化為有很強細胞毒性的 5-氟尿嘧啶核苷酸,從而抑制野生型菌株的生長[24],因此pyrG突變后即獲得了5-FOA抗性,并對尿嘧啶核苷形成依賴。實驗結果表明,獲得的4株pyrG突變株能夠在添加5-FOA的培養基上正常生長,并且未出現明顯的生長發育差異,這樣突變株與生產用菌株的形態差異很小,比較容易被生產上接受。經化學成分分析顯示,添加尿嘧啶核苷能夠顯著增加野生型紅曲霉的洛伐他汀和色素含量,同時顯著降低桔霉素含量,符合紅曲霉菌種對這3種成分的研發要求。
因pyrG突變會導致內源性尿嘧啶核苷的合成減少,理論上突變株將比野生型的洛伐他汀和紅曲色素含量減少,桔霉素含量增加。當在添加尿嘧啶核苷的培養條件下比較突變型與野生型菌株,結果顯示:(1)洛伐他汀在9個變異株中均有增加,與推測結果相反,可能與添加20 g/L尿嘧啶核苷的劑量過高有關,導致突變株的洛伐他汀含量未減少,同時來自紅曲色素、桔霉素等其他聚酮類化合物的代謝流可能也有彌補作用。(2)紅曲色素含量在4個變異株中有增加,在5個菌株中不同程度減少,增減比較隨機。考慮到紅曲色素包含安卡黃素、夢那紅、潘紅胺等多種組分,且易受光照等環境因素的影響,因此推測突變株中紅曲色素的含量變化與pyrG基因突變沒有很明顯的相關性。(3)桔霉素含量在7個變異株中顯著增加,與推測結果一致,但另外2個變異株的桔霉素含量未增反降,可能與sgRNA的非特異性打靶有關,另外,不同變異菌株之間的個體差異也會產生影響。因為洛伐他汀、紅曲色素和桔霉素皆屬于聚酮類化合物,其主干代謝途徑較一致,代謝流之間存在相互聯系,而尿嘧啶核苷對三者的作用卻并不相同,因此推測尿嘧啶核苷對代謝流的影響可能發生在桔霉素的分支代謝途徑上,導致桔霉素減少時,增加了洛伐他汀的代謝流。
本研究成功構建了體外sgRNA對紅曲霉Cas 9底盤菌株定向基因編輯的技術體系,并獲得pyrG突變株,為大量研究紅曲霉的基因功能奠定了基礎,也為研究其他真菌提供參考。補充尿嘧啶核苷能夠提高洛伐他汀和紅曲色素的含量,同時降低了桔霉素的含量,因此該研究對紅曲產品的生產有一定的指導價值。
4 結論
利用表達Cas 9的紅曲霉作為底盤菌株,通過原生質體導入外源sgRNA可以實現快速基因編輯,為重復使用底盤菌株開展大規模基因研究提供了技術參考。通過pyrG基因編輯發現,外源或內源尿嘧啶核苷的含量變化對紅曲霉桔霉素的含量有較大影響。