昆蟲幾丁質合成關鍵酶功能及其RNAi技術在害蟲防治中的研究進展
2022-09-15 14:08:26許靜靜李思琪任夢圓薛雨欣李永強
陜西農業科學 2022年8期
關鍵詞:研究
許靜靜,李思琪,任夢圓, 薛雨欣,李永強
(西北農林科技大學 植物保護學院, 農業部西北黃土高原作物有害生物綜合治理重點實驗室, 陜西 楊凌 712100)
幾丁質是昆蟲體內一種天然含量僅次于纖維素的氨基多糖[1],主要存在于昆蟲表皮、氣管和圍食膜中,起支撐和滲透屏障的作用,使蟲體減少或免于機械損傷和毒素及病原菌的侵染[2~3],因此對昆蟲的正常生長發育有著至關重要的作用[3]。由于植物和脊椎動物體內不含幾丁質,調控幾丁質合成與代謝相關酶便為新型高效殺蟲劑的設計和開發提供了潛在的優良靶標。針對這些關鍵酶的化學抑制劑以及基于RNA干擾(RNA interference, RNAi)技術的害蟲防控新途徑的研究和開發,一直是近些年農業害蟲防治領域的研究前沿和熱點[2,4]。苯甲酰基脲類是上個世紀70年代初研究最早、最具代表性的一種昆蟲幾丁質合成抑制劑,通過結構的不斷修飾,已有多個品種實現了商品化[5]。近年隨著RNAi技術介導的害蟲防治策略的提出[6],憑借其專一性強、潛在靶標多、環境安全等諸多優點,其在害蟲防控方面的研究和應用不斷取得進展。本文對近些年利用RNAi技術針對幾丁質合成關鍵酶在害蟲防控研究及應用中的重要進展、存在問題及潛在的解決途徑進行了較為系統的總結和歸納,期望為利用RNAi技術進行害蟲防治研究提供有價值的理論指導。
1 幾丁質結構與功能
幾丁質是N-乙酰氨基葡萄糖組成的線性聚合物,其化學結構與植物纖維素相似,因此也被稱為動物纖維素[7]。通過X射線分析發現,幾丁質具有α、β和γ三種晶體類型,α晶型中幾丁質鏈以反向平行的方式排列,β晶型中以平行的方式排列,γ晶型則以平行和反向平行交替的方式排列。由于幾丁質鏈反向平行排列產生了大量的氫鍵,形成的緊密微纖維有助于提高機械強度和穩定性,因此α晶型主要存在于昆蟲的體壁中。相比之下,β晶型和γ晶型鏈間氫鍵較少、緊密度較低,但二者能與水形成大量氫鍵,使形成的微纖維更具有可塑性和柔軟性,因此其主要存在于昆蟲圍食膜等中[8]。
幾丁質主要與蛋白質等共同形成昆蟲的表皮及圍食膜,從而起到保持昆蟲形態和滲透屏障的作用[3]。但幾丁質形成的堅硬體壁在阻礙外源物侵害的同時,也阻礙了昆蟲的生長發育。因此,昆蟲必須進行幾丁質有規律的合成與代謝,保證周期性的正常蛻皮,以適應正常生長發育的需求[9]。在研究果蠅的幾丁質合成酶1突變中發現,幾丁質的正常合成對表皮的完整性、形態的維持至關重要,同時還發現幾丁質合成酶1還參與了角質層色素的沉著過程[10]。而有些昆蟲在饑餓或蛻皮時,圍食膜可能完全停止形成,直至昆蟲再次取食時,并排出舊的圍食膜或重吸收產生新的圍食膜[3],因此昆蟲的生長發育嚴格依賴于體內幾丁質的含量。有研究表明,昆蟲蛻皮的干物質中幾丁質含量高達40%,而圍食膜中的幾丁質含量通常在3%~13%之間[3]。
2 幾丁質合成途徑關鍵酶
2.1 海藻糖酶
海藻糖酶(Trehalase,Tre)是幾丁質生物合成的第一個催化酶,也是昆蟲體內唯一一類水解海藻糖的非還原性酶[11],參與昆蟲的幾丁質的合成和能量代謝過程[12]。昆蟲Tre屬于糖苷水解酶家族,理論分子量約為63.4~74.2 kD,理論等電點為酸性(4.66~6.23),含有兩個氨基酸標簽序列:“PGGRFREFYYWDSY”和“QWDYPNAWPP”,以及一個甘氨酸富集區“GGGGEY”。昆蟲含有兩種海藻糖酶,即可溶性海藻糖酶(Tre1)和膜結合型海藻糖酶(Tre2)。將部分已知昆蟲Tre的氨基酸序列利用MEGA6軟件中的Align功能進行比對,再通過Neighbor-Joining進行系統進化分析,可以得到已知的昆蟲Tre基因的系統進化樹(圖1)。從該系統發育樹可以看出,不同目昆蟲的Tre基因均聚為兩大類,即Tre1和Tre2。同一種昆蟲中二者之間氨基酸序列相似性較低,但二者在不同昆蟲中卻分別存在較高的相似性。例如鱗翅目昆蟲的Tre1和Tre2在系統發育樹上各聚為一大類,表明二者之間同源性較低;而同屬鱗翅目昆蟲的草地貪夜蛾、斜紋夜蛾和甜菜夜蛾的Tre1在系統發育樹上所處的位置卻較近,表明序列同源性較高(圖1)。
通過RNAi技術對海藻糖酶功能進行研究,結果表明Tre1主要與昆蟲表皮中幾丁質含量高低有關,而Tre 2主要與昆蟲圍食膜幾丁質含量高低有關[13-14],二者的基因沉默會直接影響昆蟲的正常生長發育。Chen等[14]通過顯微注射法對甜菜夜蛾海藻糖酶SeTre1和SeTre2進行RNAi研究發現,基因轉錄水平降低60%和80%,且注射dsSeTre1的昆蟲角質層幾丁質含量顯著下降約30%,注射dsSeTre2的昆蟲中腸幾丁質含量下降約25%。張倩等[13]通過飼喂法干擾灰飛虱海藻糖酶LSTre1和LSTre2,基因轉錄水平分別降低49%和41%,并導致昆蟲體重減輕、死亡率顯著升高。由此可見,通過dsRNA干擾海藻糖酶,打破了幾丁質合成的平衡可能是導致昆蟲蛻皮困難甚至死亡的主要原因[15]。

圖1 部分已知昆蟲海藻糖酶的系統進化樹
2.2 幾丁質合成酶
幾丁質合成酶(Chitin synthetase,CHS)是幾丁質生物合成的最后一個催化酶。大多數昆蟲含有兩種幾丁質合成酶,即幾丁質合成酶1(CHS1)和幾丁質合成酶2(CHS2)[16],但在褐飛虱、豌豆蚜和大豆蚜等半翅目昆蟲中,只發現了CHS1。將部分已知昆蟲CHS的氨基酸序列使用MEGA6軟件進行同源比對,再使用Neighbor-Joining法進行系統進化分析,從而得到昆蟲CHS基因的系統進化樹(圖2)。從CHS系統發育樹可以看出,昆蟲的CHS基因也被分成了兩大類,即CHS1和CHS2。例如,所有的半翅目昆蟲的CHS1形成了一個亞分支,其中桔蚜、大豆蚜和豌豆蚜CHS1同源性較近(圖2)。幾丁質合成酶是高度保守的大分子跨膜蛋白,屬于糖基轉移酶家族,理論分子量約為160 ~180 kD,具有約14 ~17個跨膜螺旋,理論等電點偏酸性(5.43~7.08)。CHS含有3個結構域,分別為A、B、C結構域。其中位于中心的結構域B相對較為保守,含有兩個特有的識別序列“EDR”和“QRRRW”。
通過RNAi技術對幾丁質合成酶的功能進行研究,結果表明CHS1主要負責昆蟲的表皮中幾丁質的合成,而CHS2主要負責圍食膜的幾丁質形成[3]。Zhang等[17]對東亞飛蝗幾丁質合成酶LmCHS1進行基因沉默后,發現LmCHS1的轉錄水平降低了約80%;若蟲表現為發育遲緩、身體扭曲,無法完成正常的蛻皮,即使完成了蛻皮,新形成的體壁顏色較淺且后足因彎曲而無法正常行走。余志濤等[18]對中華稻蝗OcCHS1進行基因沉默后,若蟲齡期普遍增長,多數試蟲由于無法完成蛻皮而死亡,少數進入下一齡期的若蟲也因無法羽化為成蟲而死亡,死亡率高達85%。劉曉健等[19]對東亞飛蝗若蟲幾丁質合成酶LmCHS2進行基因沉默后,中腸幾丁質含量的降低使圍食膜殘缺不全,進食難以消化和吸收,最終因為饑餓而死亡。此外還發現,CHS2基因在昆蟲進食期間高表達,而蛹等非進食期間低表達[20]。這些實驗結果均表明兩種幾丁質合成酶分布和功能的差異性。
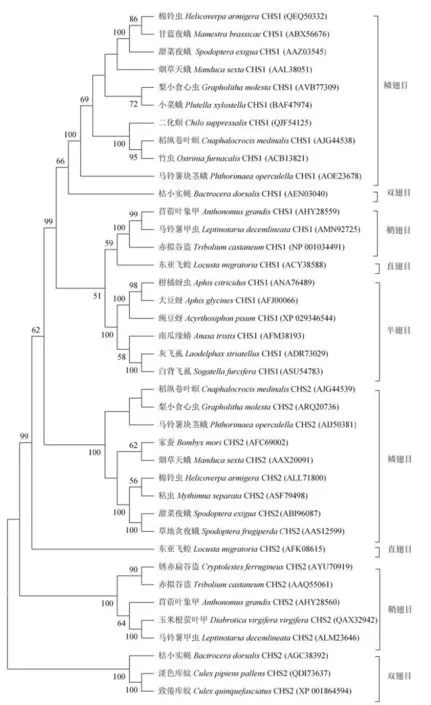
圖2 部分已知昆蟲幾丁質合成酶的系統進化樹
除此之外,許多昆蟲CHS1基因具有的可變剪接已經被報道,包括褐飛虱[21]、東亞飛蝗[17]和赤擬谷盜[22]等。值得注意的是,迄今尚未發現CHS2有可變剪切機制[22]。CHS1的兩個交替外顯子編碼的多肽長度均為59個氨基酸,且各編碼一個高度保守的跨膜結構域。CHS1a主要負責昆蟲表皮幾丁質的合成,而CHS1b主要負責氣管幾丁質的合成[12,21]。例如,張傳溪等[21]對稻褐飛虱幾丁質合成酶NlCHS1基因研究結果表明,使用dsNlCHS1處理的五齡若蟲在羽化前全部死亡,而dsNlCHS1a處理的五齡若蟲,45%在羽化前死亡,42%的個體在轉化成成蟲后死亡,僅有13%的個體存活;但NlCHS1b卻有高達73%的成蟲存活率,表明dsRNA能夠降低特定序列表達從而產生特定的效應。在東亞飛蝗中,干擾LmCHS1和其兩個可變剪接,也得到了相似的結果[17]。
2.3 幾丁質合成途徑其它催化酶
幾丁質的生物合成是一類高度復雜的生理生化過程,由一系列的酶催化完成,該過程共涉及8種酶,目前幾丁質合成通路中研究較多的是Tre和CHS,這兩類酶在多種昆蟲中已經完成了基因克隆和功能分析。相對而言,與幾丁質合成有關的其它酶,例如果糖-6-磷酸轉氨酶(Gructose-6-phosphate aminotransferase,Gfat)、葡糖胺-6-磷酸-N-乙酰轉移酶(Glucosamine-6-phosphate N-acetyltransferase,GNA)和UDP-N-乙酰葡糖胺焦磷酸化酶(UDP-N-acetylglucosamine pyrophosphorylase,UAP)等酶研究的則相對較少。
Gfat研究主要集中在酵母菌等的重要生物學功能和蛋白修飾方式方面,Gfat在昆蟲幾丁質合成途徑中的作用方面研究較少[23]。張歡歡等[23]研究表明飛蝗的果糖-6-磷酸轉氨酶基因LmGfat在蟲體表皮高表達,且LmGfat基因沉默后,試蟲主要因蛻皮失敗而死亡,推測飛蝗LmGfat與幾丁質的合成密切相關。而飛蝗的葡糖胺-6-磷酸-N-乙酰轉移酶LmGNA在蝗蟲各個組織均有表達且干擾后未出現特殊表型,通過與CHS1相似的表達模式,推測可能與幾丁質的合成有關。陳潔等[24]對5齡甜菜夜蛾UDP-N-乙酰葡糖胺焦磷酸化酶SeUAP進行干擾,甜菜夜蛾幾丁質合成酶SeCHS2表達水平明顯的下調,并出現了畸形蛹和死亡的現象。陸承聰等[25]對西花薊馬UDP-N-乙酰葡糖胺焦磷酸化酶FoccUAP進行了基因沉默,西花薊馬的羽化率和存活率均顯著降低,并出現了翅膀和腹部等畸形現象。這些試驗結果表明幾丁質合成相關酶的基因沉默會對昆蟲的生長發育產生顯著的影響,但這些酶的具體功能還有待進一步研究和闡明。
3 幾丁質合成關鍵酶調控在害蟲防治中的研究進展
3.1 針對幾丁質合成途徑的化學抑制劑
幾丁質合成抑制劑的作用相當于生長調節劑,通過對昆蟲的幾丁質合成關鍵酶活性產生重要影響,或通過和底物競爭性與Tre或CHS結合,阻斷昆蟲幾丁質的正常合成,從而導致蟲體出現畸形和死亡[3]。幾丁質合成酶最具代表性的化學抑制劑是苯甲酰基脲類和肽基核苷類化合物(表1),其中苯甲酰基脲類主要抑制昆蟲的生長發育,是幾丁質合成抑制劑中商品種類最多的一類[26],其致毒方式特別,具有選擇性強、作用范圍大和殘留少等特點,已在害蟲防治中得到廣泛應用[7]。苯甲酰基脲類殺蟲劑主要包括除蟲脲、氟蟲脲、噻嗪酮和抑食肼等。另一類肽基核苷類化合物,例如多樣霉素和尼可霉素,其結構類似于UDP-GlcNAc,作用于CHS的催化部位,具有極高的殺菌活性[27],被認為是有效的殺菌劑。雖然,目前并不認為核苷肽類化合物是殺蟲劑,但有研究發現,多氧霉素使蝗蟲在羽化時出現死亡,還導致紅頭麗蠅的圍食膜重量減輕[28]。此外,研究還發現,由于這類化合物均是極性分子,較難通過昆蟲體壁或取食后被降解,但通過注射,具有顯著的抑制效果[26]。
目前,尚未見專門針對昆蟲海藻糖酶的化學抑制劑正式登記。不過,有研究發現井岡霉素和海藻唑啉等均被發現具有顯著抑制昆蟲Tre活性的作用,因此被認為是潛在、有效的海藻糖酶抑制劑型殺蟲劑[29-30]。研究發現使用含有井岡羥胺A(Validoxylamine A)的飼料飼喂棉鈴蟲,試蟲表現為取食量減少,體重減輕,繁殖率與存活率下降等癥狀,表明對棉鈴蟲的生長發育產生了明顯的抑制作用[31]。此外,研究還發現注射了海藻唑啉的蝗蟲,首先試蟲活動減少,隨后取食量減少[29]。盡管作為有效的殺菌劑的井岡霉素和海藻唑啉,對昆蟲的Tre離體活性很高,但進一步研究發現對昆蟲活體致死效果很差。因而,研究人員希望通過對Tre抑制劑結構的改造來保證其應用效果,從而開發出具有實際應用意義的產品[32]。昆蟲生長調節劑的適當應用有利于減少環境污染,促進綠色食品生產,在農業害蟲防治中具有重要意義。
3.2 基于RNAi技術的幾丁質合成調控
RNAi是指利用雙鏈RNA(doubled-stranded RNA, dsRNA)或者小干擾RNA(small interfering RNA, siRNA)特異性地高效誘導害蟲體內重要功能基因的mRNA降解碎片化,而導致該靶基因表達沉默(即后續的翻譯表達無法正常進行),產生相應的生理功能缺失,最終導致害蟲生長發育受阻甚至致死的一種技術。該現象包含的機制最早于1998年在秀麗隱桿線蟲中得以闡明[33]。在2001年,RNAi技術被《Science》雜志評為十大科學進展之一,并位于2002年十大科學進展之首。隨后,RNAi成為了昆蟲基因功能研究的熱點和前沿,越來越多昆蟲的功能基因被克隆和鑒定,Mao等[34]在2007年提出將RNAi技術應用于害蟲防治中,而Price等[6]在2008年提出將RNAi技術作為新型害蟲防治策略,并將之視為第四代殺蟲劑。
RNAi介導的害蟲靶基因沉默途徑具有特異、高效的優點,以及技術過程的簡便性,現已被廣泛用于重要農業害蟲綠色防控新技術研究和開發中。已有研究表明,通過注射或飼喂的方法,均可降低昆蟲幾丁質的含量,產生預期的效果(表2)。鑒于幾丁質合成關鍵酶的環境安全性和靶標專一性,其做為潛在的開發新型高效綠色殺蟲劑的靶標具有巨大的應用潛力。
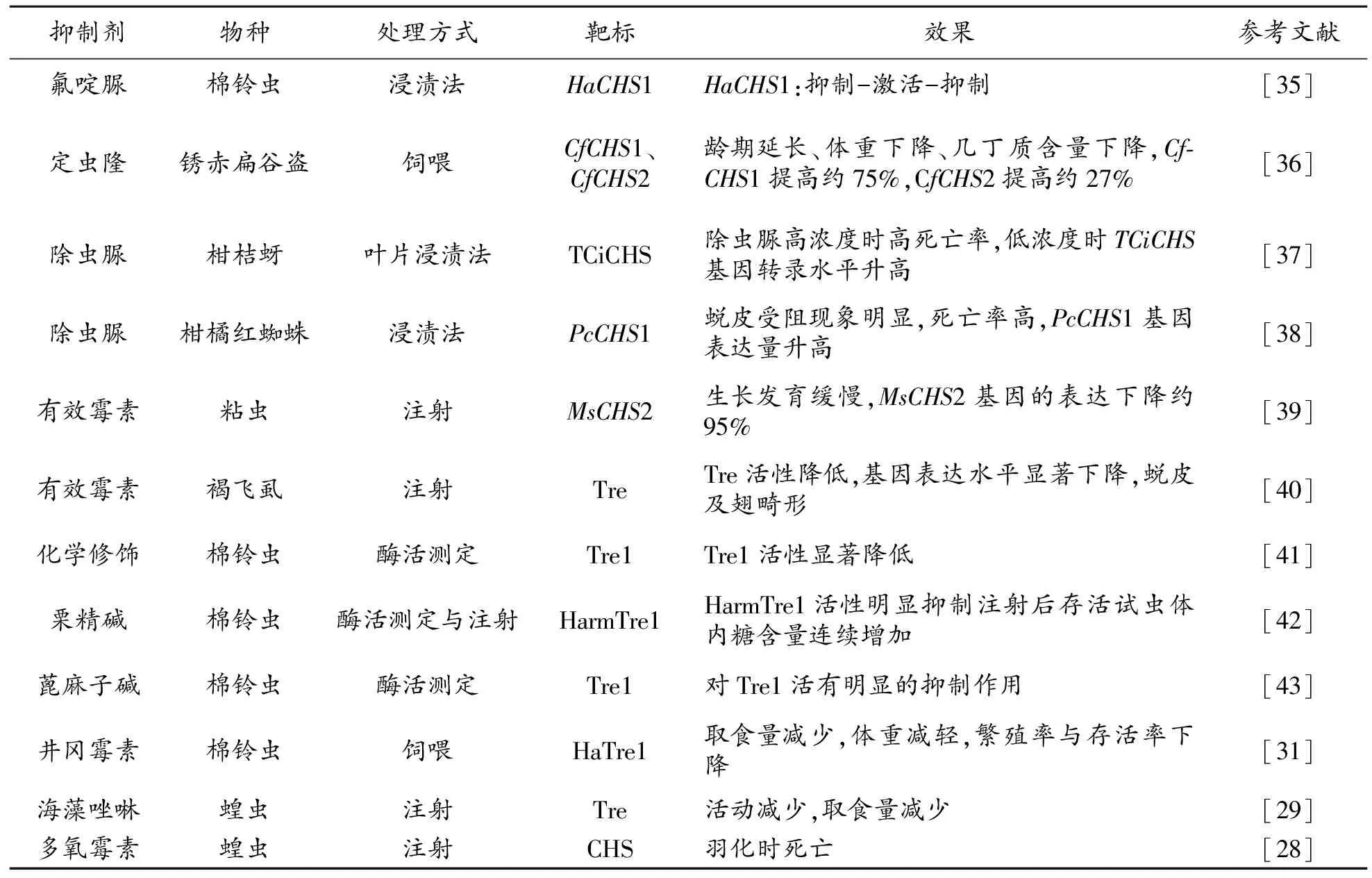
表1 針對幾丁質合成的化學抑制劑對昆蟲生長發育的影響
3.2.1 植物介導的RNAi(表達dsRNA的轉基因抗蟲作物)在害蟲防治中的應用 RNAi技術應用于害蟲防治的研究已取得較大進展。寄主誘導的基因沉默(host-induced gene silencing,HIGS)技術已在稻縱卷葉螟[44~47]、甜菜夜蛾[48]和蚜蟲[49~50]等害蟲中取得了較大進展(表2)。例如,Tian等[48]在甜菜夜蛾中報道通過飼喂在體壁和氣管特異性表達的SeCHS1的dsRNA后,顯著抑制甜菜夜蛾的生長發育,并導致昆蟲死亡。Zhao等[50]構建了含麥長管蚜幾丁質合成酶CHS1基因片段的轉基因抗蚜小麥品系,研究發現取食第三代(T3)小麥品系的試蟲,其CHS1基因表達水平下降了50%左右,存活和羽化的試蟲數量均顯著下降。Ye等[51]對豌豆蚜幾丁質合成酶ApisCHS進行RNAi研究發現,取食dsApisCHS7 2 h后導致44.7%的試蟲死亡,只有51.3%的試蟲成功蛻皮。王爽[52]構建了含稻縱卷葉螟海藻糖酶CmTre基因片段的轉基因水稻品種,室外活體實驗結果發現轉基因水稻的卷葉率和白葉率均明顯降低,表明含有CmTre的轉基因水稻對稻縱卷葉螟具有一定的防治效果。
3.2.2 dsRNA核酸農藥 上述研究表明,通過在植物中表達來增強抗蟲性是害蟲防治的重要手段。但相對于植物轉基因技術,直接體外噴灑dsRNA的方法更為簡便、快捷。大田噴施核酸用藥能夠針對不同害蟲、不同的發育時期設計特定的dsRNA,而且也能夠同時以多個基因為靶標,混合應用或者交替應用提高基因干擾和致死效率[53]。結合實際應用,噴灑及灌根是最為簡單、通用的方式。雖然,截止目前尚未有dsRNA在大田噴灑使用的報道,但室內已有用dsRNA重組菌株的菌液涂在蘿卜葉片上飼喂小菜蛾,造成其幼蟲的死亡、產卵量降低等現象,表明菌液確實對小菜蛾產生了干擾效應[57]。Hunter等[58]通過浸根和注射樹干的方式處理果樹,在7周后仍能在樹苗中檢測到dsRNA。Li等[59]通過浸根的方式處理水稻和玉米,飼喂飛虱和玉米螟,昆蟲的死亡率明顯提高。這些研究結果表明,基于RNAi技術,開發直接噴灑外用的核酸農藥(dsRNA)具有很大潛力。

表2 針對幾丁質合成途徑關鍵酶的RNAi對昆蟲生長發育的影響
4 RNAi技術應用于防治農業害蟲面臨的主要問題及應對策略
4.1 RNAi技術應用于農業害蟲防治面臨的主要問題
基于害蟲體內關鍵基因的RNAi技術開發dsRNA作為外用殺蟲劑,是新型綠色殺蟲劑創制的一個重要方向。核酸農藥(dsRNA)用于農業害蟲的防治具有以下優點:第一,dsRNA能高效專一控制目標害蟲,對天敵昆蟲無危害;其次,dsRNA的使用,能有效阻止或減緩害蟲抗藥性發生;第三,能顯著降低化學農藥的施用量,保證食品安全;此外,dsRNA屬于核酸,對環境無任何污染,是一種真正環境友好型農藥,這對減少農殘和環境污均具有重要的意義。但是,目前在研究中發現要提高RNAi效率,還有一些瓶頸問題尚需深入研究。
首先,RNAi在不同種類昆蟲中基因沉默效率存在差異性,例如研究發現赤擬谷盜等鞘翅目害蟲、蝗蟲等直翅目害蟲,以及蟑螂等蜚蠊目害蟲對RNAi較為敏感,RNAi后靶基因沉默效率較高;而棉鈴蟲、小菜蛾、草地貪夜蛾等鱗翅目害蟲,以及蚜蟲等半翅目害蟲對RNAi較不敏感,對靶基因沉默效率較低[60]。隨后研究發現,這可能是與昆蟲腸道內含有一種雙鏈RNA酶(dsRNA-specific ribonucleases , dsRNases)有關,因為其可以降解外來的dsRNA。其次,dsRNA進入昆蟲體內后的吸收、轉運和擴散機制目前還不十分清楚隨,這會影響進入昆蟲體內的dsRNA/siRNA的有效到達靶標位置,從而影響基因沉默效果[61]。此外,目前實驗室中通常制備dsRNA是采用商用試劑盒微量合成,而要作為核酸外施噴灑藥劑,則需要制備大量的dsRNA。因此,如何規模化和工廠化生產合成dsRNA,是限制dsRNA作為核酸農藥能否最終得到應用的另一重要因素。綜上,針對害蟲體內的靶標基因的RNAi效率的提高以及如何工廠化大量合成dsRNA還有許多深層問題需要闡明和解決。
4.2 主要解決途徑
4.2.1 高效基因的篩選 昆蟲體內重要靶標基因篩選和鑒定是RNAi技術應用于害蟲防治的先決要素。不是害蟲體內所有的基因都能有效沉默基因,也不是所有的基因干擾(沉默)都能殺死害蟲。根據RNAi作用的基本原理,要做為新一代高效農藥,只有那些對于昆蟲生長發育特別重要的基因才有可能作為潛在的殺蟲靶標基因。因此,在重要害蟲中篩選具有致死作用的靶標基因是將RNAi 技術應用于害蟲控制的關鍵之一[62]。目前普遍認為,作為綠色殺蟲劑設計的靶標生物分子(基因)至少要同時具備如下兩個特征:①對靶標生物的生長發育至關重要,以保證基于靶標設計的dsRNA殺蟲劑具有高效性;②在高等動植物體內不存在或顯著區別于高等動植物體內的同源分子,以保證殺蟲劑的特異性和安全性[63]。
目前主要是從害蟲消化系統(例如唾液腺、消化酶)、生長發育系統(例如幾丁質合成和代謝途徑、昆蟲蛻皮變態過程),以及繁殖系統和免疫系統(例如致育因子、卵黃蛋白)中分離鑒定具有重要功能的靶標基因[64]。通過在蝗蟲、蚜蟲、棉鈴蟲、東方粘蟲等一些昆蟲中的研究,發現除過CHS外,唾液腺蛋白[65](C002)、V-ATP酶[66](V-ATPase)和蛻皮激素受體[67](EcR)等一系列功能蛋白對昆蟲的生長發育、繁殖也起著重要的作用。昆蟲唾液相關蛋白C002是一種水溶性唾液蛋白,在取食過程中發揮著重要作用。例如,通過對麥長管蚜唾液蛋白C002基因的RNAi研究,發現蚜蟲在取食dsRNA后的第八天,死亡率達到64%[65]。Baum等[66]對V-ATPase基因有干擾作用的基因序列(WCR dsRNAs)導入玉米植株中,玉米根葉甲取食轉基因玉米植株后導致幼蟲發育延緩,明顯降低了害蟲取食對玉米生長的影響。Shen等[67]近年對棉花紅蜘蛛的蛻皮激素受體進行基因沉默(RNAi)后,發現若蟲發育為成蟲的過程明顯受阻,并導致試蟲死亡。
有上述可見,隨著今后越來越多的昆蟲基因組、轉錄組、蛋白組等數據被公布,更多的重要功能基因也會被鑒定和挖掘出來,所以將會有數量眾多的高效靶標基因可通過RNAi技術應用于害蟲防治中,這將會加速實現RNAi技術在害蟲防治中的真正應用。
4.2.2 促進RNAi應用的有效途徑 在實際應用中,許多研究發現無論哪種傳遞方式,單個關鍵基因的dsRNA都很難較快達到致死的劑量[62],只能通過調節昆蟲的生長發育使其緩慢死亡。因此,多方面的措施聯用,可增強防控效果。有助于RNAi在害蟲防治中應用的聯用措施主要有以下途徑。
首先是RNAi技術與現有的殺蟲劑聯用。RNAi技術通過靶基因的沉默,打破了昆蟲的正常生長發育,協助化學或生物殺蟲劑突破昆蟲體壁或圍食膜的阻擋,增強殺蟲劑的殺蟲效果。通過dsRNA干擾來沉默非洲瘧蚊幾丁質合成酶AgCHS1基因,使得AgCHS1基因的轉錄水平和幾丁質含量分別降低了62.8%和33.8%,增加了其幼蟲對除蟲脲的敏感性[68]。郭菊馨等[69]對淡色庫蚊海藻糖酶Tre1和幾丁質合成酶CHS1進行基因沉默,結果表明試蟲對溴氰菊酯的敏感性提高。另外研究發現,RNAi技術與Bt農藥聯用,可以累積兩種作用方式的效應。對甜菜夜蛾3齡幼蟲的SeCHS2基因進行沉默后,飼喂Bt毒素發現幼蟲對Cry毒素的敏感性顯著提高,甜菜夜蛾的高死亡率可能是靶基因SeCHS2的RNAi效應和Cry毒性的協同作用的結果[70]。
其次是多靶基因RNAi。利用多個目標基因同時沉默可進一步提高RNAi效率。趙鳳通過農桿菌轉化法獲得轉CmTre和CmCHS基因dsRNA的水稻,以其葉片飼喂稻縱卷葉螟,發現CmTre1、CmTre2、CmCHS1和CmCHS2基因表達量均顯著下降,幼蟲體型較小,畸形甚至死亡[71]。Tzin等[72]人對蚜蟲多個滲透調節基因(水通道蛋白、蔗糖酶、糖轉運蛋白)進行組合,聯合RNAi對血淋巴滲透壓的影響明顯大于單個基因的功能。由此可見,同時干擾多個具有相關作用的靶標基因更能導致害蟲生物功能的改變和有助于殺死害蟲。與現有農藥或多靶標的聯用,不僅能夠減少害蟲防治中對dsRNA濃度的要求,同時能夠減少殺蟲劑的使用劑量,有效防止農藥過量使用對環境和人的危害并延緩抗藥性的產生。
此外,還有納米粒子與dsRNA偶聯,制備納米dsRNA農藥。納米材料具有可修飾性強、水溶液分散性好和光催化降解等特點為dsRNA的傳遞提供了新的思路[73]。為了增強dsRNA在應用中的穩定性,已經研發出了無毒并可降解的納米材料,dsRNA搭載后穩定性明顯提高,噴灑在葉面上不僅耐雨水沖刷且穩定期達30 d[74]。納米材料的應用,使dsRNA不僅可以通過昆蟲取食進入昆蟲體內,還可以直接通過體壁滲透。比如,沈杰教授團隊開發出利用納米材料介導dsRNA從蚜蟲體壁進入體內的方式,從而達到干擾昆蟲生長發育的目的[75]。與通過納米粒子包被增加dsRNA吸收、傳遞效率類似的策略,還有脂質體修飾。研究表明,脂質體修飾的dsRNA在德國小蠊、斑翅果蠅等昆蟲中RNAi能明顯提高靶標基因的沉默效率[76~77]。
4.2.3 利用微生物發酵工程工廠化大規模合成生產dsRNA 利用商用試劑盒合成dsRNA雖然可以得到純度較高的dsRNA,但是成本太高,而且合成量極為有限,且操作復雜,無法滿足田間應用的需求。因此,可以利用大腸桿菌HT115(DE3) 合成多個目的基因的重組dsRNA,這樣可以極大降低合成成本,簡化合成步驟。策略是設計帶有害蟲靶標基因序列的RNAi 載體,再將載體轉入大腸桿菌感受態細胞中,就可以不斷的得到大量目的基因的dsRNA[48,62,78~79]。馬中正[78]通過基因工程菌高效合成了靶向異色瓢蟲基因的dsRNA。Vatanparast[79]通過大腸桿菌(HT115)合成了甜菜夜蛾胰凝乳蛋白酶SeCHY2基因的dsRNA,并通過超聲預處理菌液,從而提高甜菜夜蛾幼蟲口服的死亡率。
5 RNAi技術應用于害蟲防治的小結與展望
RNAi介導的靶基因沉默途徑具有特異、高效的優點,以及技術過程的簡便性,現已被廣泛用于重要農業害蟲綠色防控新技術開發中。昆蟲幾丁質合成酶,是昆蟲幾丁質代謝過程中的關鍵酶之一,由于高等動物及植物體內不存在該酶,而被認為是理想的殺蟲劑靶標[3]。目前已有眾多Tre、CHS等幾丁質合成相關的關鍵酶基因被鑒定和研究。基于幾丁質合成關鍵酶等的RNAi技術在害蟲防治中的應用可通過兩種策略實現:一是植物介導的RNAi; 二是直接體外噴灑dsRNA(核酸農藥)。雖然基于害蟲體內重要基因為靶標的RNAi技術應用于農業害蟲潛力巨大、前景誘人,但是目前還面臨一些瓶頸問題,比如不同昆蟲靶基因的RNAi效率的差異性、dsRNA合成的高成本等問題尚需進一步闡明和解決。此外,納米材料偶聯包被、脂質體修飾、與BT等殺蟲劑聯用,以及多標靶聯用等都需要系統深入開展,以促進RNAi技術在防控害蟲中得到廣泛應用。相信,隨著對幾丁質合成途徑研究的不斷深入,基于該途徑的關鍵酶基因為靶標的RNAi技術在今后害蟲綠色防治中將會發揮重要作用。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
