水產(chǎn)動物小分子熱休克蛋白家族研究進展
2022-09-15 07:49:26李婷婷孫繼紅韓森榮劉麗鄒雯靜宋堅常亞青湛垚垚
水產(chǎn)學雜志 2022年4期
李婷婷,孫繼紅,韓森榮,劉麗,鄒雯靜,宋堅,常亞青,湛垚垚
(大連海洋大學農(nóng)業(yè)農(nóng)村部北方海水增養(yǎng)殖重點實驗室,遼寧 大連 116023)
熱休克蛋白(Heat shock proteins,HSPs),又稱熱應(yīng)激蛋白,是一類廣泛存在于原核和真核生物體內(nèi)具有高度保守性的生物大分子[1]。根據(jù)相對分子質(zhì)量,可將熱休克蛋白分為HSP100、HSP90、HSP70、HSP60 和小分子熱休克蛋白(Small heat shock proteins,sHSPs5 個主要家族[2]。sHSPs 是指相對分子質(zhì)量介于12~30 kDa 的熱休克蛋白[3]。根據(jù)其結(jié)構(gòu)、功能及表達部位,又進一步分為Ⅰ類和Ⅱ類兩個主要亞家族。Ⅰ類亞家族成員無組織特異性表達,能參與泛素-蛋白酶體系統(tǒng)(Ubiqutin-proteasome system,UPS)介導的蛋白質(zhì)降解過程,包括HSPB1、HSPB5、HSPB6 和HSPB8;Ⅱ類亞家族成員具有組織表達特異性,在哺乳動物的肌源性組織、睪丸組織和眼晶狀體等部位特異性表達,包括HSPB2、HSPB3、HSPB4、HSPB7、HSPB9 和HSPB10[4,5]。除了典型Ⅰ類和Ⅱ類兩類亞家族成員外,在后來的研究中又先后鑒定出了HSPB11、HSPB12 和HSPB15,3個非典型成員(表1)。
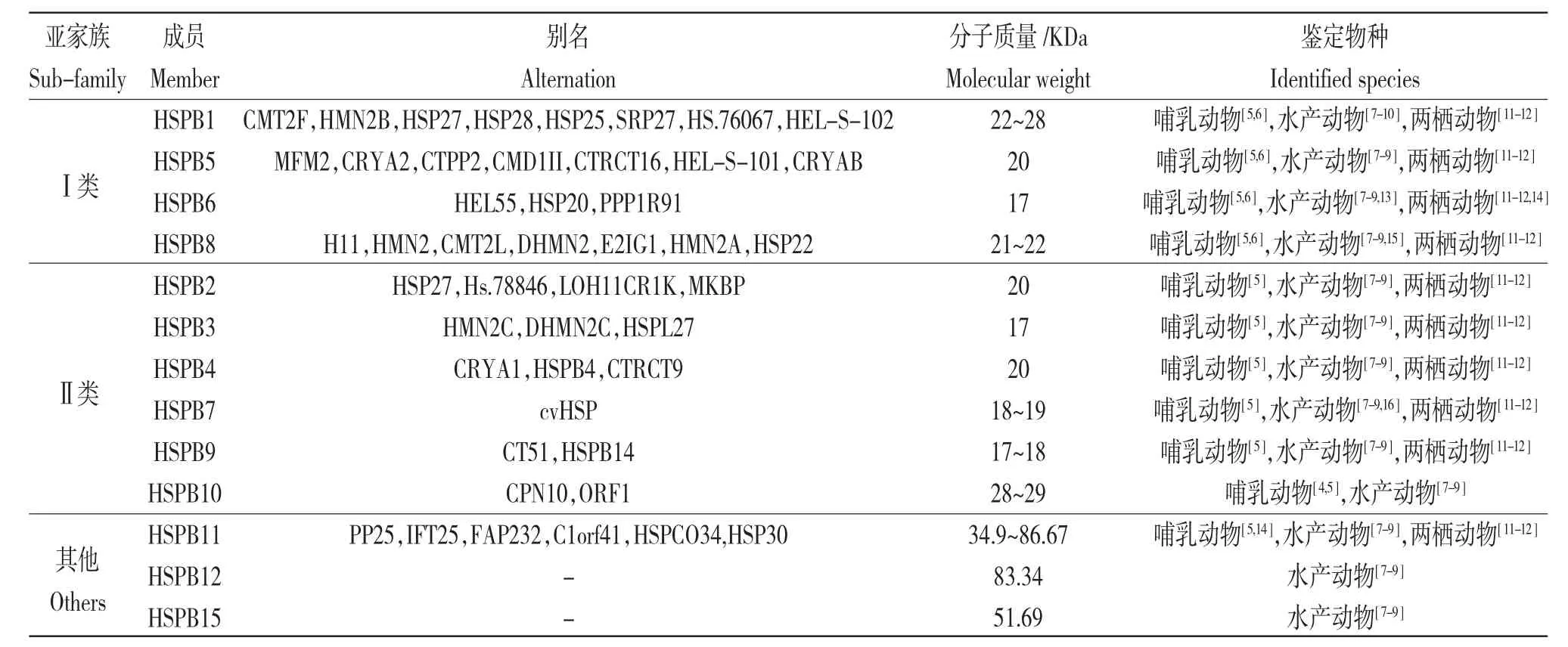
表1 常見小分子熱休克蛋白家族成員統(tǒng)計Tab.1 Summary of members in common small heat shock proteins sub-family
截至2021 年4 月,美國國立生物技術(shù)信息中心(National Center for Biotechnology Information,NCBI)數(shù)據(jù)庫中登錄的sHSPs 家族基因序列信息多達10 858 條,其中所占比例較高的物種為哺乳動物(992 條;占比9.14%)和水產(chǎn)動物(814 條;占比7.50%)。但是,相較于哺乳動物,水產(chǎn)動物sHSPs 家族成員的研究相對滯后,主要集中在魚類、腔腸動物及軟體動物。研究表明,sHSPs 不僅在水產(chǎn)動物的生長發(fā)育過程中發(fā)揮重要作用,還可能參與介導水產(chǎn)動物的免疫防御反應(yīng)。為進一步了解水產(chǎn)動物sHSPs 家族成員的系統(tǒng)進化關(guān)系及生物學功能,本文綜述了近年來水產(chǎn)動物sHSPs 家族成員的序列特征、進化特點及生物學功能進行,旨在豐富水產(chǎn)動物sHSPs 家族基因的研究數(shù)據(jù),為深入了解水產(chǎn)動物sHSPs 家族基因成員的生物學信息提供參考。
1 sHSPs 家族成員的特征
1.1 sHSPs 家族成員的分子結(jié)構(gòu)特征
sHSPs 家族成員都具有可變的N 端結(jié)構(gòu)域(N-terminal domain,NTD),靈活的C端結(jié)構(gòu)域(C-terminal domain,CTD)和高度保守的α-晶狀體結(jié)構(gòu)域(alpha-crystalline domain,ACD)[17-19]。其中,NTD 由24~247 個氨基酸殘基組成,富含芳香族氨基酸殘基;NTD 的主要作用是影響sHSPs 低聚物的結(jié)構(gòu)和發(fā)揮分子伴侶(Molecular chaperone)功能[18]。CTD 的氨基酸殘基個數(shù)大都小于20 個,高度保守的IXI/V 結(jié)構(gòu)將CTD 分為“尾巴”和“延伸”結(jié)構(gòu)。CTD 的主要功能是增加sHSPs 四級結(jié)構(gòu)的穩(wěn)定性和復(fù)合物的溶解性。ACD 是由90~100 個氨基酸殘基組成的β-三明治夾心結(jié)構(gòu),由7 或8 個反向平行的β-折疊片層構(gòu)成,哺乳動物的sHSPs 缺乏β6 鏈,ATD 的主要功能是形成sHSPs 二聚體[19](圖1)。
1.2 sHSPs 家族成員的表達特征
sHSPs 為最常見的分子伴侶,在發(fā)揮作用時會形成大型動態(tài)低聚物。該低聚物呈空心球狀,在沒有與目標蛋白結(jié)合前以非活性的形式存在[1]。當機體受到外源刺激或脅迫(如高溫、低氧或磁場干擾等)而出現(xiàn)蛋白質(zhì)折疊紊亂時,機體內(nèi)的sHSPs 從非活性狀態(tài)轉(zhuǎn)變?yōu)榧せ顮顟B(tài),以不依賴ATP 的形式與錯誤折疊的蛋白(目標蛋白)相結(jié)合,將其隔離在低聚復(fù)合物中,并通過依賴ATP 的大分子熱休克蛋白促進錯誤折疊蛋白的重新折疊,保證應(yīng)激狀態(tài)下機體內(nèi)空間結(jié)構(gòu)的穩(wěn)定性[20];當外源刺激或脅迫消失后,隨著錯誤折疊蛋白的減少,sHSPs 又逐漸恢復(fù)到非活性狀態(tài)[9,15,16]。除了非活性狀態(tài)到激活狀態(tài)的轉(zhuǎn)變之外,sHSPs 相對表達量的顯著增加也是機體內(nèi)sHSPs 家族成員響應(yīng)外源刺激或脅迫的策略之一[6,21,22]。Pedro 等[21]對搖蚊幼蟲Chironomus riparius進行了熱休克(35℃)和異源物(鎘,Cd)處理。結(jié)果顯示,與對照組相比,熱休克組和異源物處理組中搖蚊幼蟲的hsp27 基因的相對表達量都顯著增加;還有研究證實,鼠海馬體神經(jīng)元細胞的hspb1、hspb5、hspb6、hspb8 和hspb11 基因在亞砷酸、氧化和應(yīng)激條件下的相對表達量都顯著增加[6]。
2 水產(chǎn)動物中sHSPs 的研究進展
2.1 水產(chǎn)動物sHSPs 家族的主要種類及系統(tǒng)進化分析
目前,水產(chǎn)動物的sHSPs 家族成員主要集中在Ⅰ類亞家族(占比52.17%),而歸屬于Ⅱ類亞家族(占比33.33%)和其他亞家族(占比14.50%)的成員較少(表2)。將18 種水產(chǎn)動物中歸屬于Ⅰ類亞家族的sHSPs 的核苷酸序列與5 種哺乳動物和1 種兩棲動物中相應(yīng)的sHSPs 的核苷酸序列進行系統(tǒng)進化分析發(fā)現(xiàn)(圖2),水產(chǎn)動物的HSPB1(除斑點叉尾Ictalurus punctatus 和克氏原螯蝦Procambarus clarkii 外)聚為一支,最終與哺乳動物和兩棲動物聚為一大支,表明HSPB1 在物種進化過程中相對保守。哺乳動物和兩棲動物的HSPB8 亞型聚為較近的一支,最終與水產(chǎn)動物中的魚類聚為一大支;水產(chǎn)動物中棘皮類的HSPB8 亞型和細粒棘球蚴Echinococcus granulosus 單獨聚為一支,提示水產(chǎn)動物的HSPB8 亞型種內(nèi)的差異性較大,且存在明顯的種間分化現(xiàn)象。哺乳動物和兩棲動物的Ⅰ類亞家族中的HSPB6 亞型聚為較近的一支,但是,水產(chǎn)動物中的魚類和棘皮類的HSPB6 亞型則分別聚為一支。值得注意的是,哺乳動物和兩棲動物的HSPB6 亞型最終沒有與水產(chǎn)動物的HSPB6 亞型聚為一大支,而是與所有動物的HSPB5 亞型聚為一大支。這一結(jié)果在一定程度上證實了水產(chǎn)動物的HSPB6 亞型種內(nèi)差異性較大,種間分化明顯,與哺乳動物和兩棲動物的HSPB6 亞型親緣關(guān)系較遠,而與哺乳動物和兩棲動物的HSPB5 亞型親緣關(guān)系較為密切。
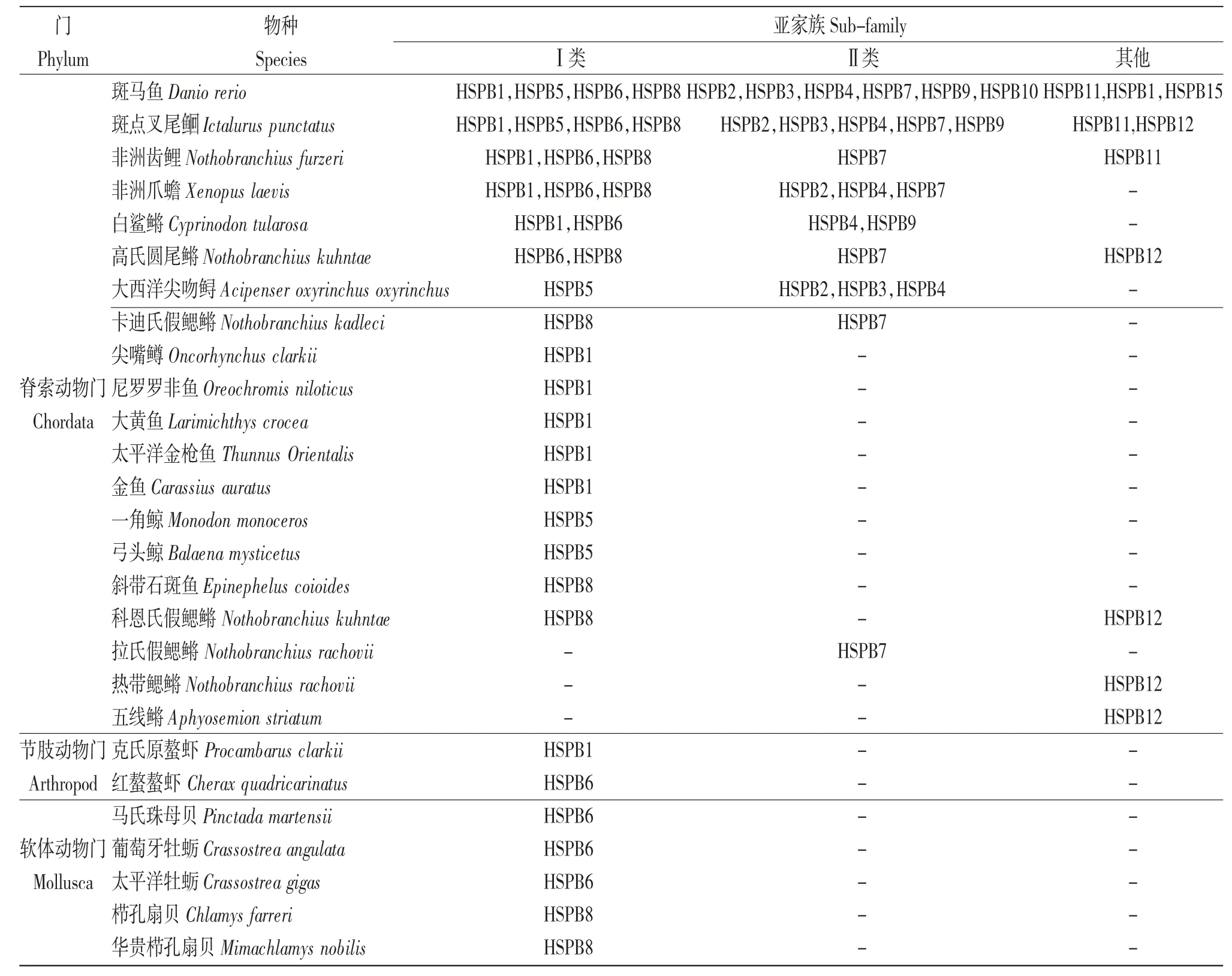
表2 NCBI 數(shù)據(jù)庫中27 種水產(chǎn)動物小分子熱休克蛋白家族成員核苷酸序列統(tǒng)計(截至2021 年3 月)Tab.2 Statistics of the nucleotide sequences of the members of the small-molecule heat shock protein family in 27 species aquatic animals in the NCBI database(up to March 2021)
將8 種水產(chǎn)動物歸屬于Ⅱ類亞家族的sHSPs的核苷酸序列與3 種哺乳動物和1 種兩棲動物中歸屬于Ⅱ類亞家族的sHSPs 核苷酸序列進行系統(tǒng)進化分析發(fā)現(xiàn)(圖3),所有分析物種(除斑點叉尾外)中的HSPB2 和HSPB3 亞型聚為一大支,HSPB9和HSPB10 亞型聚為一大支,表明HSPB2 和HSPB3亞型,HSPB9 和HSP10 亞型的核苷酸序列較相近,親緣關(guān)系較密切。所有動物的HSPB7 亞型聚為較近的一大支。此結(jié)果表明,水產(chǎn)動物、哺乳動物和兩棲動物的HSPB7 亞型在進化過程中相對保守。
2.2 水產(chǎn)動物sHSPs 家族的生物學功能
2.2.1 分子伴侶功能
分子伴侶是能夠識別并結(jié)合錯誤折疊或裝配,而本身不參與最終產(chǎn)物形成的一類蛋白[1]。αA-晶狀體蛋白(αA-crystallin)是最早被發(fā)現(xiàn)具有分子伴侶功能的sHSPs[22],隨后也證實,許多其他的sHSPs具有分子伴侶活性[15,16]。如在對小鼠αB-晶狀體蛋白(αB-crystallin)進行敲除后發(fā)現(xiàn),鄰近的hspb2基因也受到破壞,αB-晶狀體蛋白和hspb2 基因功能的喪失導致小鼠骨骼肌的退化[23,24]。
Magda 等[15]利用免疫共沉淀(Co-immunoprecipitation,Co-IP)、蛋白質(zhì)免疫印跡(Western blot,WB)等技術(shù)研究了受精120 h 后斑馬魚Danio rerio胚胎中的hspb8 基因。結(jié)果分析表明,HSPB8 可以作為Bag3 分子伴侶,參與形成自噬(autophagy)誘導復(fù)合物。Emily 等[16]使用類轉(zhuǎn)錄激活因子效應(yīng)物核酸酶(Transcription activator-like effector nuclease,TALEN)技術(shù)使斑馬魚hspb7 等位基因發(fā)生突變,hspb7 基因的突變導致細絲蛋白(Filamin-C,F(xiàn)LNC)的功能下降,導致斑馬魚心肌細胞修復(fù)損傷的能力下降,隨后的免疫沉淀(Immunoprecipitation,IP)和WB 實驗進一步證實HSPB7 是FLNC 的分子伴侶。
2.2.2 參與調(diào)節(jié)細胞的生長和分化
隨著對水產(chǎn)動物sHSPs 的不斷深入研究,發(fā)現(xiàn)某些水產(chǎn)動物細胞中的sHSPs 不僅具有分子伴侶功能,還參與維持機體的正常形態(tài)特征以及調(diào)節(jié)機體細胞的生長與分化[15,16]。
Magda 等[15]發(fā)現(xiàn),給受精48 h 后的斑馬魚胚胎注射嗎啉寡核苷酸后能干擾阻斷HSPB8 的翻譯,減少HSPB8 蛋白的表達,導致斑馬魚胚胎身體彎曲、心包水腫和生長速度減慢等形態(tài)變化,證明hspb8具有維持斑馬魚胚胎形態(tài)與生長的重要作用。Emily等[16]研究發(fā)現(xiàn),與對照組相比,敲除了hspb7 基因的斑馬魚心臟形態(tài)畸形。
2.2.3 調(diào)節(jié)細胞凋亡
細胞調(diào)亡(Apoptosis)又叫程序性細胞死亡,是細胞內(nèi)外環(huán)境發(fā)生改變、受到死亡信號的觸發(fā)或在基因調(diào)控下引起的細胞死亡的過程[25,26]。sHSPs 作為細胞凋亡的調(diào)控因子可以通過多種方式調(diào)控細胞凋亡[10,25-27]。
斑節(jié)對蝦Penaeus monodon 在感染對蝦白斑綜合癥病毒(White spot syndrome virus,WSSV)2 h 后,其肌肉組織中hsp21 基因的相對表達量顯著降低,隨后脫氧核糖核苷酸末端轉(zhuǎn)移酶介導的缺口末端標記法(Tdt-mediated dutp nick-end labeling,TUNEL)檢測發(fā)現(xiàn),斑節(jié)對蝦肌肉細胞內(nèi)DNA 片段化的細胞數(shù)量增加,其肌肉細胞出現(xiàn)凋亡的癥狀,分析表明,對蝦白斑綜合癥病毒可能通過抑制斑節(jié)對蝦肌肉組織內(nèi)hsp21 基因的相對表達量而誘導斑節(jié)對蝦肌肉細胞的凋亡[27]。
石少婷等[10]發(fā)現(xiàn),在櫛孔扇貝中的“p38MAPKHSP27”通路中,扇貝多肽(Polypeptide from Chlamys farreri,PCF)可通過抑制p38MAPK 的活性,降低hsp27 磷酸化水平,抑制中波紫外線誘導的人角質(zhì)形成細胞HaCaT 細胞凋亡。
2.2.4 免疫調(diào)節(jié)功能
sHSP 是水產(chǎn)動物先天性免疫(Innate immunity)的重要調(diào)節(jié)因子以及宿主和病原菌相互作用的關(guān)鍵參與者。研究證明,sHSPs 可調(diào)控水產(chǎn)動物響應(yīng)病原菌的生理生化過程[28,29]。
Lei 等[29]研究發(fā)現(xiàn),哈維氏弧菌(Vibrio harvey)感染后,太平洋牡蠣Crassostrea gigas hsp20 基因在其血細胞中的相對表達量顯著上升。Li 等[28]通過qPCR 技術(shù)鑒定了鰻鱺弧菌Vibrio anguillarum 對菲律賓蛤仔Ruditapes philippinarum 中的兩種sHSPs亞型——Vpshsp-21 和Vpshsp-2 基因表達的影響。結(jié)果顯示,與對照組相比,VpsHSP-1 基因在6 h 和96 h 時的相對表達量顯著增加,具有統(tǒng)計學差異;VpsHSP-2 基因的相對表達量在48~72 h 內(nèi)急劇增加。兩個基因的不同表達趨勢表明,Vpshsp-21 和Vpshsp-2 基因參與菲律賓蛤仔對感染鰻鱺弧菌的響應(yīng)過程[30-35]。
3 展望
綜上所述,水產(chǎn)動物sHSPs 家族成員可在細胞生長發(fā)育、細胞凋亡及免疫應(yīng)答等多種細胞生理過程中發(fā)揮重要作用,還可作為環(huán)境污染監(jiān)測的分子標記物(表3)。因此,進一步深入研究水產(chǎn)動物sHSPs 家族成員在水產(chǎn)動物生長、發(fā)育和免疫應(yīng)答等方面的功能和作用,不僅進一步加深了對水產(chǎn)動物sHSPs 家族成員及生物學功能的認識和理解,還可為水產(chǎn)動物資源保護、養(yǎng)殖增產(chǎn)以及病害防控等提供一定的分子生物學基礎(chǔ)數(shù)據(jù)。
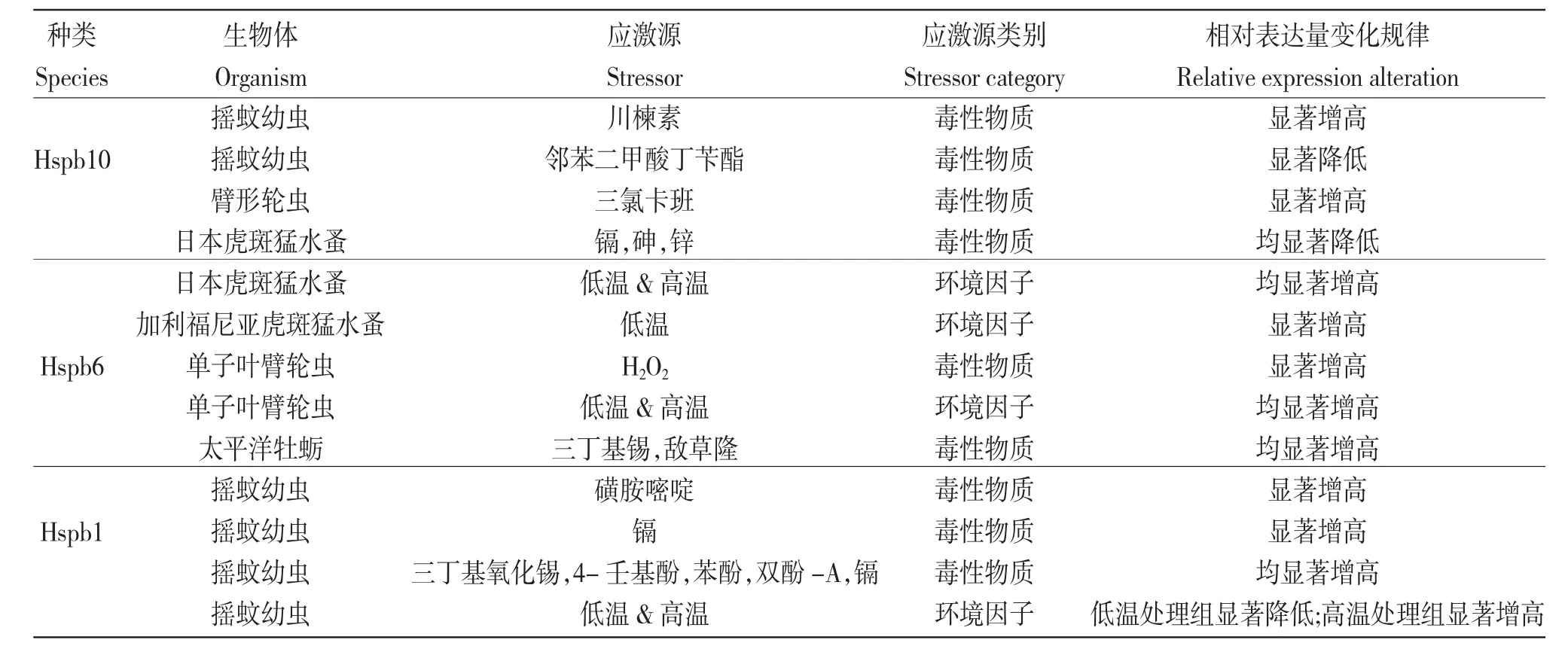
表3 6 種水生生物中小分子熱休克蛋白對應(yīng)激源響應(yīng)表達規(guī)律的總結(jié)Tab.3 Summary of expression patterns of small heat shock proteins in 6 species aquatic organisms in response to stressors
目前水產(chǎn)動物sHSPs 家族成員的序列和結(jié)構(gòu)信息仍來源于少數(shù)物種,且sHSPs 家族成員生物功能鑒定及解析的研究相對匱乏,今后應(yīng)重點開展以下3 方面工作:首先,進步擴大水產(chǎn)動物的種類,豐富和充實水產(chǎn)動物sHSPs 家族成員的序列信息和結(jié)構(gòu)信息,理清水產(chǎn)動物sHSPs 家族成員之間的系統(tǒng)進化地位及相互聯(lián)系;其次,充分利用現(xiàn)代生物技術(shù)(如高通量測序、多組學聯(lián)動分析等)和生物信息數(shù)據(jù)分析技術(shù),深入開展不同種類水產(chǎn)動物sHSPs 家族成員的生物功能研究,進一步理清不同水產(chǎn)動物sHSPs 家族成員表達模式與規(guī)律的共性特征以及種屬特異性特征及其在不同生理或病理條件下的響應(yīng)規(guī)律,全面掌握水產(chǎn)動物sHSPs 家族成員的生物功能;最后,在理論研究的基礎(chǔ)上,系統(tǒng)評估水產(chǎn)養(yǎng)殖動物中sHSPs 家族成員作為種質(zhì)選育標記和養(yǎng)殖水環(huán)境檢測標記物的開發(fā)潛力和應(yīng)用價值,探索水產(chǎn)養(yǎng)殖動物sHSPs 家族成員作為種質(zhì)選育標記和養(yǎng)殖水環(huán)境檢測標記物的有效應(yīng)用方式和方法。
