植物氮素吸收利用相關NPF基因家族研究進展
2022-09-19 09:27:26王化敦馬鴻翔
植物營養與肥料學報 2022年8期
王化敦,張 鵬*,馬鴻翔
(1 江蘇省農業科學院 / 江蘇省農業生物學重點實驗室,江蘇南京 210014;2 揚州大學農學院 / 江蘇省糧食作物現代產業技術協同創新中心/江蘇省作物基因組學與分子育種重點實驗室,江蘇揚州 225009)
氮(N)作為生物大分子如核酸、蛋白質的基本組分,是植物生長發育需要量最多的礦質營養元素,也是大多數農業生態系統中作物產量的限制因子[1]。在通氣土壤中,不同形態的人工合成氮肥在微生物(主要是硝化細菌)作用下轉變為硝態氮(NO3--N),成為無機態氮素的主要存在形式[2];在淹水條件下,作物(如水稻)根系通過泌氧和根系分泌物形成的根際微環境可將不同形態氮轉化為硝態氮,亦是植物利用的重要氮素形態[3]。
目前,植物中已報道參與硝態氮吸收和運輸的轉運蛋白基因家族包括:NPF(nitratetransporter1/peptidetransporterfamily)、NRT2(nitratetransporter2)、CLC(chloridechannels)和SLAC1/SLAH(slowanion channel-associated1homologues)[4-6]。在以上4個基因家族中,NRT2、CLC和SLAC1/SLAH家族成員數量較少(5~7個),其中NRT2家族編碼高親和力(highaffinity)轉運蛋白,與伴侶蛋白NAR2 (nitrate assimilation related protein)結合,在對低氮環境的響應中具有重要功能[4];CLC家族編碼氯離子(Cl-)通道蛋白,后來發現與在液泡中的儲存和運輸有關[7-8];SLAC1/SLAH家族編碼一類對電壓反應遲緩類型(slow type)的離子通道,通過向保衛細胞運輸Cl-和引發氣孔閉合過程,其家族成員SLAH3主要在中柱表達,可特異性轉運,與在根與地上部之間的長距離運輸有關[9-10]。
NPF家族包括NRT1(nitratetransporter1)和PTR(peptidetransporter)兩類基因,前者一般認為編碼低親和力(low-affinity) NO3-轉運蛋白,后者編碼寡肽轉運蛋白[11],由于二者序列相似性較高,在進化關系上處于同一分支,將NRT1/PTR基因統一命名為NPF(NRT1PTRfamily)[12]。研究表明,NPF基因除了作為硝態氮轉運蛋白或寡肽轉運蛋白轉運和寡肽之外,還可轉運其它多種底物(、Cl-、生長素、脫落酸、赤霉素、茉莉酸、硫代葡萄糖苷、砷酸二甲酯等),參與多種生物與非生物脅迫響應[13-14]。與其它轉運蛋白家族相比,NPF家族成員眾多,模式植物擬南芥含有53個NPF基因,糧食作物水稻、玉米和小麥中分別含有93、79和331個NPF基因[12,15-16]。近年來,NPF基因的功能獲得了較多關注和深入研究,模式植物擬南芥中已有超過一半(31/53)NPF家族成員的生物學功能被解析,糧食作物水稻中亦有16個NPF基因的生物學功能被報道(表1、圖1)。大量研究表明,NPF基因廣泛參與植物對氮素的吸收和利用過程,在改良和提高作物氮素利用率及產量相關性狀中具有重要作用和應用價值。本文主要針對模式植物擬南芥和糧食作物中已報道NPF基因在氮素吸收利用中的生物學功能進行綜述,以期深入理解植物高效吸收和利用氮素的機理,為提高作物氮素利用率相關研究以及作物氮高效育種實踐提供參考。
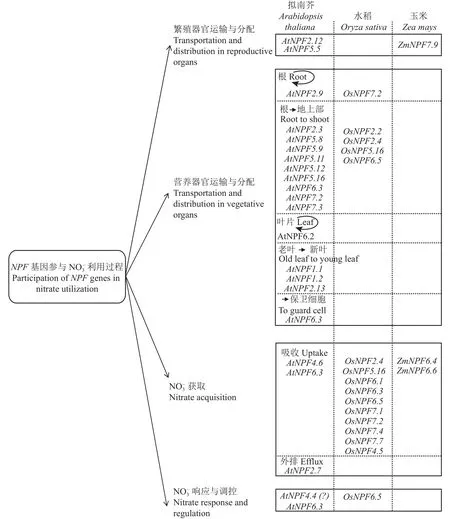
圖1 模式植物擬南芥和糧食作物中NPF基因在參與利用中的功能Fig.1 Diverse functions of NPF genes in nitrate utilization in Arabidopsis and main food crops

表1 模式植物擬南芥和主要糧食作物中已報道生物學功能的NPF基因Table 1 Functionally characterized NPF genes in Arabidopsis and main food crops

續表1 Table 1 continued

續表1 Table 1 continued

續表1 Table 1 continued
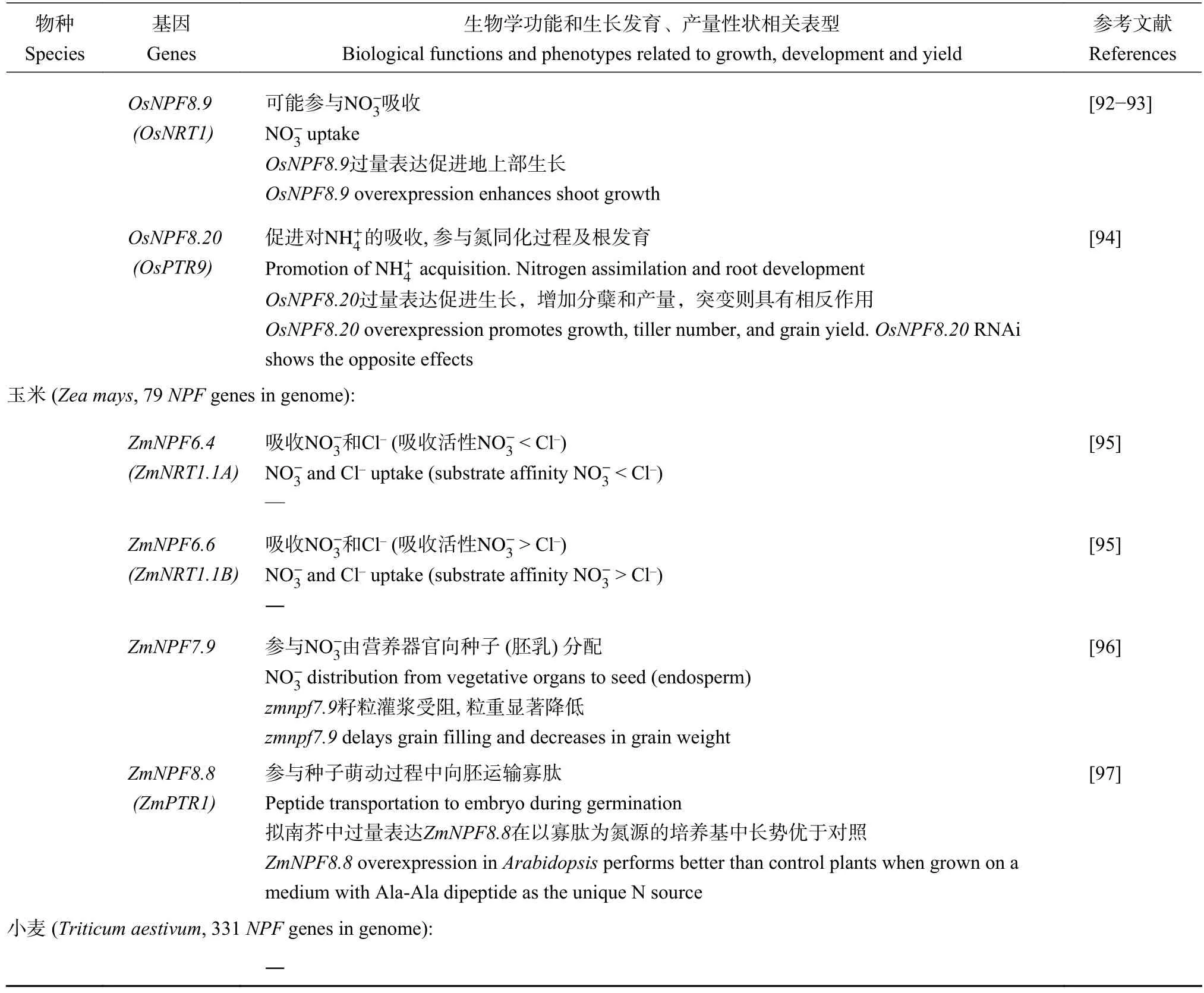
續表1 Table 1 continued
1 模式植物擬南芥中NPF基因在氮素吸收和利用中的生物學功能
1.1 參與氮素吸收與外排
AtNPF6.3(NRT1.1/CHL1)是植物中第一個被克隆的硝態氮(NO3--N)轉運蛋白基因,主要在根中表達,受誘導表達顯著上調[48]。AtNPF6.3兼具低親和力、高親和力(即dual-affinity)吸收特性,由其第101位蘇氨酸(Thr)是否磷酸化決定[98]。蛋白晶體結構分析表明,當環境中濃度充足時,AtNPF6.3中Thr101去磷酸化形成二聚體,降低了蛋白結構的靈活性,對具有低親和力吸收特性;當環境中缺乏時,Thr101磷酸化使AtNPF6.3由二聚體解離為單體,提高了蛋白結構的靈活性,對具有高親和力吸收特性[99-100]。不同于AtNPF6.3,AtNPF4.6(AtNRT1.2)僅編碼低親和力轉運蛋白,參與吸收過程[36]。AtNPF4.6主要在根毛和根表皮細胞組成型表達,并且在atnpf6.3中抑制AtNPF4.6的表達進一步降低了對的吸收,說明AtNPF6.3介導的誘導型雙親和力吸收功能和AtNPF4.6介導的組成型低親和力吸收功能相對獨立[36]。植物對的獲取是根中吸收與外排活動的綜合結果[101]。AtNPF2.7(NAXT1)主要在成熟根的皮層細胞表達,其表達受到轉錄后水平的調控,當環境酸化引起細胞質pH降低時,AtNPF2.7在蛋白水平表達顯著增加,參與根中的外排[21],這一生理活動可能反映了植物對外界環境變化(脅迫)的適應性。此外,NPF基因家族部分成員編碼寡肽轉運蛋白(peptide transporter,PTR),參與植物對有機態氮素的吸收過程,如atnpf8.1(ptr1)在以寡肽為氮源的培養基上生長時吸收的氮素顯著降低[71]。
1.2 參與氮素在營養器官中的運輸和分配
NPF基因亦參與在地上部不同組織器官中的運輸和分配過程。AtNPF6.3在氣孔保衛細胞中亦有較強表達,atnpf6.3在含培養條件下氣孔開放受阻,這一現象與脫落酸(abscisic acid)對氣孔開放的抑制作用和細胞內CO2水平無關,研究發現atnpf6.3中保衛細胞濃度顯著降低,誘導的去極化現象消失,說明AtNPF6.3通過向保衛細胞運輸參與了氣孔活動[50]。AtNPF6.2(AtNRT1.4)主要在葉柄中表達,對維持葉片內部(葉柄、葉脈及葉片部位)的動態平衡具有重要作用,該基因突變導致葉柄中濃度顯著降低,而葉片中濃度顯著升高[46]。與幼嫩葉片(新葉)相比,成熟葉片因表面積較大具有較強的蒸騰作用,可以從蒸騰作用驅動的木質部流中獲取更多,而發育中的幼嫩葉片(新葉)相比成熟葉片需要更多的氮素供應,AtNPF1.1(AtNRT1.12)和AtNPF1.2(AtNRT1.11)共同參與了這一生物學過程,二者主要在葉片主脈韌皮部伴胞表達,并且在成熟葉片中的表達量較高,同位素示蹤試驗發現雙突變體atnpf1.1atnpf1.2中直接由根轉運而來的15更多流向已經成熟的葉片而非新葉[17]。當外界供應不足時,體內貯藏的有效動員和再分配對于植物體生長尤其是幼嫩組織的發育具有重要作用,AtNPF2.13(AtNRT1.7)參與此生物學過程,該基因主要在老葉葉脈韌皮部細胞表達,并受氮饑餓誘導表達上調,突變導致老葉中濃度顯著增加,而老葉韌皮部傷流液和新葉濃度顯著降低[29]。進一步研究發現,AtNPF2.13的表達受miR827-NLA模塊的調控,NLA(nitrogenlimitationadaptation)編碼泛素連接酶,介導AtNPF2.13經泛素化途徑降解,氮饑餓條件下NLA的表達受到miR827靶向負調控,促進下游AtNPF2.13表達上調[30]。因此,為了滿足幼嫩組織(葉片)生長發育對氮素的需求,當環境中充足時,AtNPF1.1和AtNPF1.2可以將成熟葉片主脈中的(由根轉運而來)供給新葉;當環境中缺乏時,AtNPF2.13可促進老葉中貯藏的再分配至新葉。
1.3 參與氮素向繁殖器官的運輸和分配
在繁殖生長階段,植物體由根直接吸收的氮素以及營養器官中儲存的氮素大部分將向繁殖器官運輸和分配。在NPF基因家族中,AtNPF8.2(AtPTR5)編碼寡肽轉運蛋白,主要在花粉、胚珠和種子中表達,參與有機態氮素向繁殖器官的運輸和分配過程,花粉管萌發試驗中該基因增強表達株系的花粉在含毒性二肽(丙氨酰乙硫氨酸)的培養基中萌發嚴重受阻,而敲除突變體在相同培養基中花粉管生長受影響程度最低[71]。
除有機態氮素(氨基酸、寡肽、多肽等)外,無機態的硝態氮()也可以在繁殖器官積累,并影響種子發育過程。AtNPF2.12(AtNRT1.6)僅發現在繁殖器官(角果、果柄等)維管束表達,并在授粉后表達量顯著增加,該基因敲除導致種子中濃度顯著降低,形態學分析發現突變體在受精后胚胎發育的1-或2-細胞期,胚柄細胞出現過度分裂與變形萎縮,后期種子敗育率顯著增加,這一結果說明作為無機態氮源的對于擬南芥早期胚胎發育具有重要作用,AtNPF2.12參與了這一生物學過程[27]。AtNPF5.5在胚中檢測有表達,突變導致正在發育的胚中總氮含量顯著降低,影響了胚中氮素的積累,其精細表達模式以及影響胚中氮素積累的機制和對種子發育的影響有待進一步研究[43]。
1.4 參與調控植物對氮素()的響應
AtNPF6.3除了作為轉運蛋白基因吸收和轉運外,還參與調控植物對環境中的響應。首先,AtNPF6.3作為信號因子參與對的初級響應:PNR (primary nitrate response),即供處理短時間內(0.5~1 h)轉運蛋白基因及代謝相關基因迅速增強表達[51]。AtNPF6.3與細胞質膜上受誘導表達上調的離子通道基因CNGC15互作抑制其功能,的供給解離AtNPF6.3與CNGC15的互作,促進后者對Ca2+的吸收,引起細胞質中Ca2+水平顯著提高,激活蛋白激酶CPK10/30/32對信號轉導途徑中關鍵轉錄因子NLP7的磷酸化,促進NLP7的質-核穿梭進而激活下游PNR基因[52,102-103]。這一信號轉導途徑與AtNPF6.3中第101位蘇氨酸(Thr)是否磷酸化有關,當環境中濃度較低時,被誘導迅速增強表達的蛋白激酶CIPK23-CBL9復合物對AtNPF6.3中Thr101磷酸化,對具有低水平的初級響應;當環境中濃度較高時,AtNPF6.3 (NRT1.1/CHL1)中Thr101去磷酸化,引發對高水平的初級響應[51]。其次,AtNPF6.3參與對次級響應:SNR(secondary nitrate response),即PNR中相關基因的表達在長時間處理條件下的反饋抑制[104]。一方面,AtNPF6.3可以誘導轉錄因子LBD37/38/39表達負向調控PNR基因(如NRT2.1)的表達[53,105];另一方面,最近報道的PNR負向調控因子NIGTs基因位于關鍵轉錄因子NLP7下游,其中NIGT1.3和NIGT1.4對PNR基因的抑制作用依賴AtNPF6.3-CNGC15-Ca2+-CPK-NLP模塊,由于NLP對下游誘導基因的直接正向調控相比NLP-NIGTs途徑對下游基因的負向調控反應更為迅速,從而導致很多轉運蛋白基因(NPF、NRT2)和代謝相關基因(NR、NIA)表現出對的初級響應(PNR)和次級響應(SNR)[104,106-107]。再次,AtNPF6.3參與植物根系適應環境中不同濃度的“覓食”過程。當環境中濃度較低時,AtNPF6.3可以轉運生長素,減少側根原基和新生側根中生長素的積累,導致側根生長發育受阻;當環境中濃度較高時,AtNPF6.3的生長素轉運功能受到抑制,側根原基和新生側根中生長素濃度增加,促進側根發育[54]。
最近,Chen等[35]報道AtNPF4.4(NRT1.13)在調節體內的分配中具有重要功能。AtNPF4.4定位在細胞質膜,主要在葉柄和莖節部位靠近木質部的薄壁細胞中表達。atnpf4.4表現出濃度依賴的晚花、分支發生與生長缺陷,進一步研究表明,atnpf4.4中經由節向葉片、分支中的“橫向”分配減少,并且在低氮(0.2 mmol/L)條件下更為顯著。值得注意的是,由于在第10和第11跨膜結構域之間高度保守,對轉運活性具有重要作用的脯氨酸位點被絲氨酸取代(P487S),AtNPF4.4體外試驗不具有轉運的功能,但可以結合,說明AtNPF4.4可能具有感知體內水平,并通過調節在節部位的橫向分配以維持植物對低氮環境的適應性。由于AtNPF4.4不具有轉運功能,其參與體內分配的分子機制以及承擔轉運功能的組分有待進一步發掘。
2 糧食作物中NPF基因在氮素吸收和利用中的生物學功能
2.1 水稻
目前,糧食作物中有關NPF基因的研究主要集中在水稻中(表1、圖1)。已報道OsNPF2.4、OsNPF5.16、OsNPF6.1、OsNPF6.3、OsNPF6.5、OsNPF7.1、OsNPF7.2、OsNPF7.4和OsNPF7.7參與吸收過程[76,79-82,85-86,90],OsNPF4.5在根系通過叢枝菌根共生途徑獲取中具有重要功能[78],進一步拓寬了人們對植物獲取途徑的認知,這些基因在參與吸收過程中是否存在互作關系(協同、冗余、拮抗等)有待深入研究。其中OsNPF2.4和OsNPF6.5與另外一個家族成員OsNPF2.2在根維管組織(木質部)中表達量較高,與擬南芥中AtNPF2.3、AtNPF6.3和AtNPF7.3功能相似,三者亦參與由根向地上部的長距離運輸過程[75-76,82];OsNPF5.16在根、莖基部和葉鞘中表達水平較高,亦參與由根向葉鞘的分配過程[79];OsNPF7.2主要在根伸長區和成熟區厚壁細胞、皮層與中柱表達,除了參與吸收,還與在根中不同部位的分配有關[86];OsNPF7.7存在兩種可變剪接OsNPF7.7-1(編碼較長產物)和OsNPF7.7-2(編碼較短產物),分別定位在細胞膜(OsNPF7.7-1)和液泡膜(OsNPF7.7-2),提高其表達水平則分別促進了對和的吸收[90]。值得注意的是,OsNPF5.16與已報道NFP7亞家族成員OsNPF7.1、OsNPF7.2、OsNPF7.3、OsNPF7.4和OsNPF7.7均參與水稻蘗芽的發育,并最終影響水稻分蘗。其中OsNPF7.1和OsNPF7.4在蘗芽中對不同供氮水平具有相反的表達模式,對蘗芽的生長發育分別具有促進和抑制作用[85]。分析表明,OsNPF5.16和OsNPF7.2表達變化影響了莖蘗基部細胞分裂素(cytokinins, CKs)水平,并且后者同時影響了獨腳金內酯(strigolactones, SLs)信號途徑相關基因的表達,說明CKs信號途徑參與了OsNPF5.16依賴的蘗芽生長發育[79],CKs和SLs信號途徑協同參與了OsNPF7.2依賴的蘗芽生長發育[87],對于其它NPF基因(OsNPF7.1、OsNPF7.3、OsNPF7.4和OsNPF7.7)參與蘗芽生長發育是否涉及CKs、SLs或其它激素(如生長素),有待進一步研究。
OsNPF6.5(OsNRT1.1B)是AtNPF6.3(NRT1.1/CHL1)在水稻中的功能性同源基因,該基因定位在細胞膜上,受誘導表達顯著增強[82]。與AtNPF6.3類似,OsNPF6.5不僅參與吸收和由根向地上部的長距離運輸,還參與調控水稻對初級響應[81-82]。不同于擬南芥PNR反應中依賴AtNPF6.3、由第二信使Ca2+和磷酸化修飾介導信號關鍵轉錄因子AtNLP7的質-核穿梭[104],在OsNPF6.5調控的PNR反應中,磷信號途徑關鍵抑制因子OsSPX4可以與信號關鍵轉錄因子OsNLP3互作抑制其質-核穿梭,可以促進OsNPF6.5與OsSPX4結合,并招募OsNPF6.5互作蛋白OsNBIP1 (OsNRT1.1B Interacting Protein 1)介導OsSPX4經泛素化途徑降解,增強OsNLP3的質-核穿梭引發PNR反應,同時促進了磷信號途徑相關基因的表達[83]。因此,水稻中OsNPF6.5作為關鍵因子整合了信號轉導途徑(PNR反應)和受調節的磷信號轉導途徑(即促進對磷的吸收利用)。進一步研究表明,OsNPF6.5還可以調節水稻根際氮代謝功能相關微生物區系組成,影響根際對氮素的吸收[84]。Wang等[81]報道了AtNPF6.3在水稻中的另外一個同源基因OsNPF6.3(OsNRT1.1A)。不同于OsNPF6.5定位在細胞膜,OsNPF6.3定位在液泡膜,并且受另外一種無機態氮素銨誘導表達顯著增強。與OsNPF6.5基因功能相比,OsNPF6.3不僅能促進對的吸收,還促進了對的吸收,說明OsNPF6.3在對不同形態氮素的吸收和利用中可能起著更為基礎的作用[81]。
2.2 玉米
目前,糧食作物玉米中已報道4個NPF基因的生物學功能(表1、圖1)。ZmNPF7.9在胚乳轉移細胞特異性表達,該基因突變導致籽粒中濃度顯著降低,并引發嚴重的籽粒發育障礙(胚乳發育遲緩、淀粉沉積異常、粒重顯著降低等),說明無機態的對于玉米籽粒(胚乳)發育具有重要作用,ZmNPF7.9參與了這一生物學過程[96]。ZmNPF8.8(ZmPTR1)主要在種子萌發過程中盾片上皮細胞中表達,擬南芥中異源表達ZmNPF8.8在二肽(Ala-Ala)作為唯一氮源的培養基中,種子吸脹后萌發的存活率顯著高于對照材料,說明ZmNPF8.8可能參與萌發籽粒中有機態氮素(二肽等)向胚的運輸過程[97]。
借助定點誘變和電生理技術,Wen等[95]對玉米中AtNPF6.3兩個同源基因ZmNPF6.4 (ZmNRT1.1A)和ZmNPF6.6 (ZmNRT1.1B)的吸收特性進行了深入研究,發現二者在功能上出現分化,分別對Cl-和具有高親和力(high-affinity)吸收特性。ZmNPF6.6中含有與結合的關鍵組氨酸位點(His-362, 擬南芥AtNPF6.3中該位點為His-356[99-100]),而在ZmNPF6.4中該位點被酪氨酸(Tyr-370)取代。將ZmNPF6.6中該關鍵組氨酸位點突變成酪氨酸(His-362-Tyr),導致ZmNPF6.6喪失了對的轉運功能;在ZmNPF6.4中引入該關鍵組氨酸位點(Tyr-370-His),則使ZmNPF6.4獲得了對的高親和力吸收特性[95]。通過改變NPF基因編碼蛋白中特定氨基酸位點,為提高作物氮素吸收利用(包括對Cl-的耐受性)提供了新的視角。
2.3 小麥
糧食作物小麥是異源六倍體(基因組類型為AABBDD),基因組龐大并且十分復雜[108]。目前,小麥中有關NPF基因在氮素吸收利用中生物學功能的研究尚未見報道,但已有證據表明NPF基因參與小麥對氮素的吸收利用過程[109-111]。最近,Wang等[15]和Li等[16]分別對小麥中NPF基因家族進行了系統鑒定與分析,發現小麥基因組中含有高達331個NPF基因,其數量遠高于水稻(93個)和玉米(79個)。小麥中家族成員眾多的NPF基因在氮素吸收利用中的生物學功能有待發掘與解析。
3 結論與展望
如前所述,目前糧食作物玉米(含79個NPF基因)中僅有4個NPF基因的生物學功能被報道,在小麥(含331個NPF基因)中尚未有相關報道,未來對玉米和小麥中NPF基因的發掘與功能研究,將為改良作物氮素利用效率提供新的基因資源。此外,現有文獻報道中對NPF基因功能的研究多采用水培與盆栽試驗,試驗條件為單一氮水平(高氮或低氮)處理。在自然環境中,作物生長在多重氮水平條件下,并且在不同生長發育階段對氮素的需求各異,因此,實踐中需要考慮綜合運用多種策略(調節氮素吸收、轉運、分配/再分配、代謝及其調控基因的綜合表達)探索提高作物的氮素利用效率。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
現代畜牧科技(2021年4期)2021-07-21 06:13:00
福建基礎教育研究(2019年1期)2019-09-10 07:22:44
福建基礎教育研究(2019年1期)2019-05-28 08:39:49
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
醫學研究雜志(2015年11期)2015-06-10 06:44:03
