連續施用化肥及秸稈還田對潮土酶活性、細菌群落和分子生態網絡的影響
2022-09-19 09:26:50魏建林周曉琳李子雙李國生吳小賓劉兆輝譚德水
植物營養與肥料學報 2022年8期
關鍵詞:物種
馬 壘,李 燕,魏建林,周曉琳,李子雙,李國生,吳小賓,劉兆輝,譚德水*
(1 山東省農業科學院農業資源與環境研究所 /農業農村部廢棄物基質化利用重點實驗室,山東濟南 250100;2 德州市農業科學研究院,山東德州 253015)
潮土是黃淮海平原最主要的土壤類型,其砂粒含量高、土壤結構差,在常年麥玉兩熟的高強度利用下,土壤肥力水平普遍低下[1]。為了滿足糧食需求,大量化肥被施入土壤,為作物生長補充大量養分,迅速提升作物產量。然而不合理化肥施用導致土壤出現板結、酸化,還會引發農業面源污染等環境問題[2]。秸稈還田配施化肥是一種可持續農業生產措施,可有效緩解單施化肥引起的土壤質量下降問題[3]。越來越多的研究表明,秸稈與化肥配合施用可顯著提升土壤有機質和養分含量,提高土壤酶活性,改良土壤結構,增加作物產量,同時降低秸稈焚燒引起的空氣污染[4-7]。但化肥及秸稈還田對土壤微生物群落的影響仍有待明確。
土壤微生物是農田生態系統中最為活躍的組分,其在有機質形成與降解、養分循環轉化、土壤團聚體結構形成中具有重要作用[8]。由于微生物對其生存環境變化極其敏感,可對細小的土壤環境變化迅速做出反應,因此研究微生物群落的變化可較為直觀地反映土壤生態功能的變化[9]。關于化肥和秸稈還田對微生物群落影響的研究已有大量報道,大多數研究發現,長期施用化肥引起土壤酸化,導致微生物多樣性降低[10-11];而秸稈還田配施化肥可緩解酸化問題,從而提高土壤微生物多樣性,并促進土壤中參與碳、氮、磷循環的特定微生物生長[12-14]。然而一些研究者發現長期秸稈還田后土壤微生物多樣性并未發生明顯改變[5,15]。我國安徽砂姜黑土長期定位試驗結果表明,連續30年秸稈還田配施化肥后細菌多樣性顯著降低[3,16]。這種不一樣的結論,一方面是由于試驗地土壤養分狀況和自然條件不同,另一方面可能與秸稈還田量、還田方式有關。此外,大多數研究的焦點在于微生物多樣性或群落結構,忽視了對種間關系的研究。土壤中的微生物并非單獨存在,不同物種通過頻繁的相互作用,形成復雜的生態網絡,共享土壤生態位和營養資源。微生物參與的生態功能不僅僅是不同個體的集體特征,還與它們之間的相互作用密切相關。研究物種間的網絡互作關系對于理解微生物群落構建過程,了解物種間的生態位差異或環境異質性,預測土壤生態功能具有重要意義[17]。因此從網絡共現模式角度解析長期化肥及秸稈還田對微生物群落影響的機制仍需探討。
以位于山東德州的潮土長期施肥定位試驗為研究平臺,采用16S rRNA高通量測序技術,對連續10年不施肥、單施化肥、秸稈還田配施化肥后土壤細菌群落多樣性、結構、組成展開研究;同時通過構建分子生態網絡,研究不同施肥模式對細菌種間互作關系和網絡共現模式的影響;探索細菌群落與土壤理化性質、酶活性之間的關系,以期為黃淮海潮土區土壤肥力提升和秸稈資源化利用提供理論依據。
1 材料與方法
1.1 長期試驗地點概況
田間定位試驗始于2010年10月,位于山東省德州市哨馬營村 (116°17'44'' E,37°30'34'' N)。該地屬暖溫帶半濕潤季風氣候,年平均氣溫12.9℃,年降水量547.5 mm,供試土壤為非石灰性潮土。試驗地為冬小麥-夏玉米輪作,小麥品種為濟麥22,玉米品種為鄭單958。試驗開始前土壤基本理化性質如下:pH 8.3、有機質20.3 mg/kg、全氮1.2 g/kg、全磷0.83 g/kg、全鉀20.8 g/kg、有效磷37.4 mg/kg、速效鉀282 mg/kg。
1.2 土壤樣品采集與測定
2021年在該長期定位試驗選擇不施肥料對照(CK)、單施化肥(NPK)和秸稈還田配施化肥(NPKS)3個處理進行取樣分析,每個處理4個重復,共計12個小區。NPK和NPKS處理小麥季施N 300 kg/hm2、P2O5120 kg/hm2、K2O 100 kg/hm2,玉米季施N 250 kg/hm2、P2O545 kg/hm2、K2O 45 kg/hm2,NPKS 處理小麥和玉米秸稈粉碎后全量還田,于下茬作物播種前旋耕翻入土壤。供試化肥為尿素(N 46%)、過磷酸鈣 (P2O516%)和硫酸鉀 (K2O 50%)。
土壤樣品于2021年小麥灌漿期采集。截止樣品采集時,共連續種植了11季小麥、10季玉米。按照S形采樣法,使用直徑5 cm的土鉆采集0—20 cm土壤,每個小區采取7鉆混合成1個樣品重復。所有樣品置于保溫箱,采用冰袋保鮮運回實驗室。土壤樣品在去除大的石塊和植物根系后過2 mm篩。所有樣品分為3部分,一部分常溫風干后用于土壤理化性質測定,一部分4℃保鮮用于土壤酶活性測定,另一部分-80℃保存用于土壤DNA提取。
土壤理化指標測定均采用常規方法[18]。pH采用水土比(體積質量比) 5∶1提取,pH計測定;有機質(SOM)采用重鉻酸鉀容量法測定;全氮(TN)采用濃硫酸消煮,全自動凱氏定氮儀測定;全磷(TP)采用濃硫酸-高氯酸消煮,鉬銻抗比色法測定;堿解氮(AN)采用堿解擴散法測定;有效磷(AP)采用0.5 mol/L碳酸氫鈉浸提—鉬銻抗比色法測定;速效鉀(AK)采用醋酸銨浸提—火焰光度計法測定。土壤酶活性采用北京索萊寶科技有限公司試劑盒測定:β-葡糖苷酶采用對硝基酚比色法測定,以單位時間單位質量土樣催化對-硝基苯-β-D 吡喃葡萄糖苷生成的對-硝基苯酚表示;脲酶采用靛酚藍比色法,以單位時間單位質量土樣水解尿素產生的銨態氮量表示;堿性磷酸酶采用磷酸苯二鈉比色法,以單位時間單位質量土樣催化磷酸苯二鈉水解生成的苯酚量表示;脫氫酶采用氯化三苯基四氮唑還原法,以單位時間單位質量土樣還原氯化三苯基四氮唑(TTC)生成的三苯基甲臜量表示。土壤生化指標結果均以烘干土(105℃,24 h)為基準表達。
采用Fast DNA Spin Kit for Soil (MP Biomedicals,Santa Ana,CA,USA)試劑盒提取0.50 g鮮土DNA,具體步驟參見說明書。采用16S rRNA V4~V5區進行細菌高通量測序,使用515F/907R通用引物進行PCR擴增。每個樣品前端引物均含有不同的7 bp Barcode用于區分不同樣品。PCR擴增條件包括94℃5 min,90℃ 60 s,55℃ 60 s,72℃ 75 s,30 個循環,之后 72℃ 10 min。反應產物采用QIA quick PCR Purification kit (Qiagen)進行純化。將不同樣品的PCR擴增產物等摩爾混合后,采用Illumina公司MiSeq測序儀完成序列分析。
1.3 數據統計分析
高通量測序數據處理主要使用Usearch軟件[19]完成:1)合并雙端測試序列;2)切除引物并去除質量分數低于20的低質量序列;3)采用RDP數據庫(http://rdp.cme.msu.edu/)去除嵌合體;4)使用UNOISE算法去噪生成擴增子序列變體(amplicon sequence variant,ASV);5)與RDP數據庫進行比對進行注釋。經過質量控制后,總共得到673598條高質量序列(每個樣品48929~68929條序列),所有樣品隨機抽取48929條序列進行后續分析。測序原始數據已上傳至NCBI的SRA數據庫(SRR17968322~SRR 17968333),項目編號為PRJNA805075。
細菌alpha多樣性(包括Richness豐富度指數、Shannon多樣性指數和Peilou’s evenness均勻度指數)、基于Bray-curtis距離的主坐標分析(PCoA)、冗余分析(RDA)和置換多元方差分析(Adonis)均使用R 4.0.5的“vegan”包進行計算。多元回歸樹分析(MRT)采用“mvpart”包進行計算。不同處理ASV水平指示種分析采用“indicspecies”包進行計算,屬水平差異物種在Galaxy網站(http://huttenhower.sph.harvard.edu/galaxy/)采用線性判別分析(LEfSe)進行計算,其中線性判別分析(LDA)值大于2 的物種為顯著差異物種。采用“WGCNA”包計算ASV間Spearman相關系數,為了減少網絡復雜度,只選取|r|>0.9,P<0.001的關系用于網絡構建;采用Fast Greedy法進行網絡模塊分析,使用Cytoscape 3.8.2軟件進行網絡圖形繪制。單因素方差分析和Pearson相關性雙尾檢驗采用SPSS 25.0軟件完成,采用Fisher LSD法進行平均值多重比較。采用Microsoft Excel 2019 和Origin 2021b軟件進行數據處理和繪圖。
2 結果與分析
2.1 土壤理化性質
由表1可知,經過連續10年不同施肥處理后,土壤理化性質發生明顯改變。與CK處理相比,NPK和NPKS處理的土壤pH顯著降低了0.06個單位(P<0.05);NPK處理土壤有機質和堿解氮含量分別提升23.19%和24.22%,NPKS處理則分別提升34.82%和39.17%,較NPK處理分別提升9.20%和12.03%;與CK處理相比,NPK處理全氮、全磷、有效磷和速效鉀分別升高了26.80%、27.06%、298.39%和26.32%,NPKS處理分別升高了30.93%、17.65%、310.43%和25.74%,但NPK和NPKS處理間差異未達顯著水平(P>0.05)。
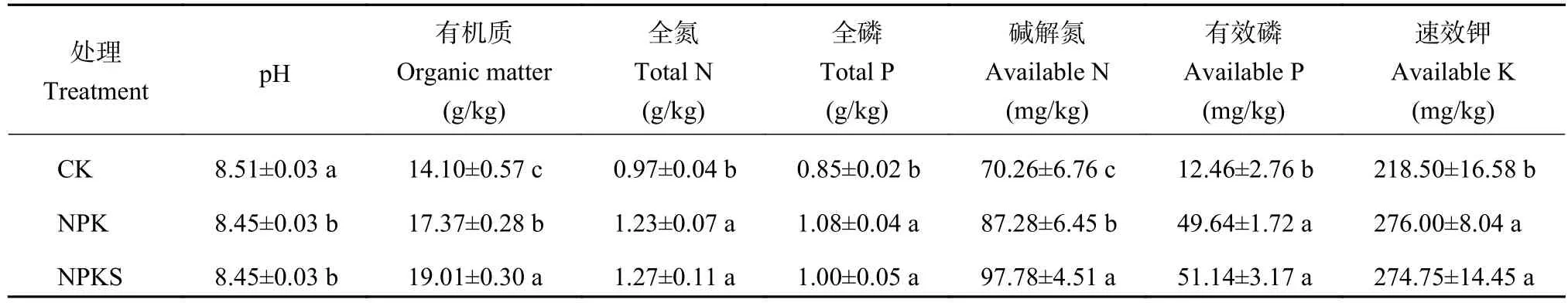
表1 長期不同施肥處理下土壤理化性質Table 1 Physio-chemical properties of soil under different long-term fertilizations
2.2 土壤酶活性
通過測定β-葡糖苷酶(β-GC,降解纖維素)、脲酶(UE,水解尿素)、堿性磷酸酶(ALP,水解有機磷)和脫氫酶(DHA,表征微生物活性)等4種土壤胞外酶活性,表征長期施用化肥及秸稈還田對土壤微生物功能和活性的影響(圖1)。β-GC活性高低順序為NPKS>NPK>CK,NPKS處理β-GC活性較CK和NPK分別增加了78.31%和17.09%,NPK處理較CK處理升高47.91%。與CK處理相比,NPK和NPKS處理ALP活性分別顯著增加了50.35%和46.53%;DHA在NPKS處理中最高,比CK和NPK處理分別提高50.91%和37.21%。UE活性3個處理間差異不顯著(P>0.05)。
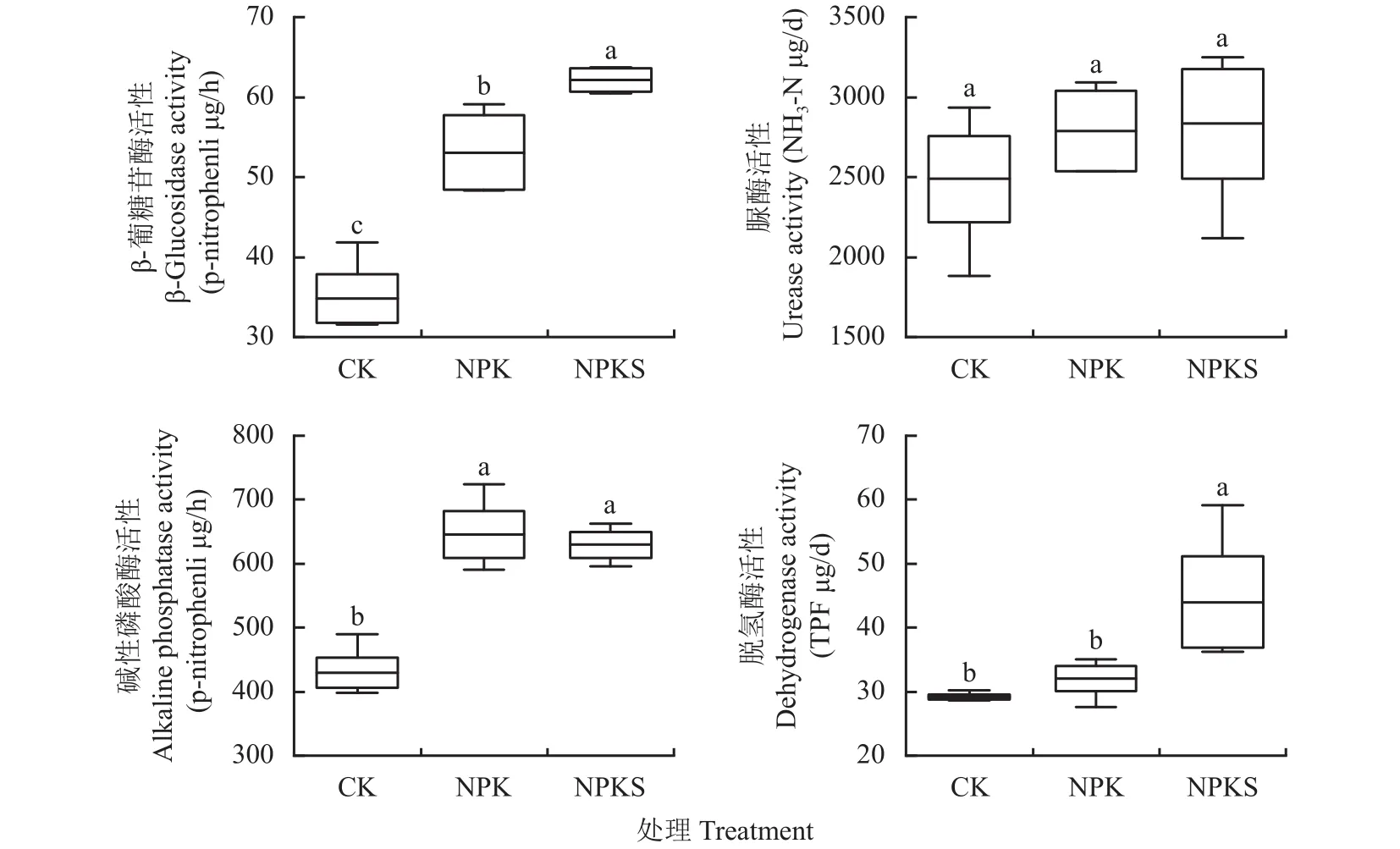
圖1 長期不同施肥處理下土壤酶活性Fig.1 Soil enzyme activities under different long-term fertilizations
2.3 土壤細菌Alpha 多樣性
通過16S rRNA高通量測序總共得到673598條高質量序列,可劃分為5993個ASV (每個樣品4860~5088條ASV),經過與RDP數據庫進行比對,這些ASV歸屬于23個門,50個綱,69個目,145個科和323個屬。將所有樣品均一化為48929條序列后,不同施肥處理細菌Alpha多樣性如圖2a所示。細菌多樣性(Diversity)和均勻度(Evenness)在CK處理中最高(11.11和0.90),而在NPK處理(11.02和0.89)和NPKS處理(11.01和0.89)中均顯著降低;豐富度(Richness)處理間差異未達顯著水平。
相關性分析結果(圖2b)表明,細菌多樣性與土壤有機質 (r=0.76,P<0.01)、全氮 (r=0.59,P<0.05)、堿解氮 (r=0.59,P<0.05)、有效磷 (r=0.81,P<0.01)、速效鉀 (r=0.60,P<0.05)、β-葡糖苷酶(r=0.62,P<0.05)、堿性磷酸酶 (r=0.60,P<0.05)等生化指標呈顯著負相關,與pH (r=0.74,P<0.05)呈顯著正相關;細菌均勻度與土壤有機質 (r=0.81,P<0.01)、全氮 (r=0.59,P<0.05)、堿解氮 (r=0.62,P<0.05)、有效磷 (r=0.81,P<0.01)、速效鉀 (r=0.62,P<0.05)、β-葡糖苷酶 (r=0.68,P<0.05)、堿性磷酸酶(r=0.59,P<0.05)等生化指標呈顯著負相關,與pH(r=0.75,P<0.05)呈顯著正相關;細菌豐富度則與土壤生化指標相關性未達顯著水平。以上結果說明長期施用化肥和秸稈還田后,土壤養分含量的升高反而導致細菌Alpha多樣性顯著降低。

圖2 長期不同施肥處理下土壤細菌Alpha 多樣性Fig.2 Soil bacteria alpha-diversity under different long-term fertilizations
2.4 土壤細菌群落結構和組成
長期施用化肥及秸稈還田下土壤細菌群落結構和組成發生明顯改變。變形菌門(Proteobacteria)是所測潮土中的優勢菌門(圖3a),其相對豐度在12個樣品中介于40.40%~49.20%,其余豐度大于1%的物種包括酸桿菌門(Acidobacteria,相對豐度16.10%~21.30%)、放線菌門(Actinobacteria,相對豐度8.01%~11.80%)、浮霉菌門(Planctomycetes,相對豐度3.54%~5.20%)、擬桿菌門(Bacteroidetes,相對豐度3.07%~7.01%)、綠彎菌門(Chloroflexi,相對豐度1.96%~3.55%)、WPS-1 (相對豐度0.89%~1.41%)和厚壁菌門(Firmicutes,相對豐度0.67%~1.24%)。基于Braycurtis距離對細菌群落結構進行主坐標分析(PCoA)(圖3b),結果發現主坐標的前兩軸總共解釋了34.06%的群落差異(第一軸和第二軸分別解釋了21.31%和12.75%的群落差異):CK處理聚集在主坐標第一軸的左側,NPK處理聚集在主坐標第一軸的右側,而NPKS處理介于兩者之間。置換多元方差分析(Adonis)結果表明,NPKS處理與CK (R2=0.27,P=0.03)和NPK處理(R2=0.20,P=0.03)細菌群落結構均具有顯著差異。
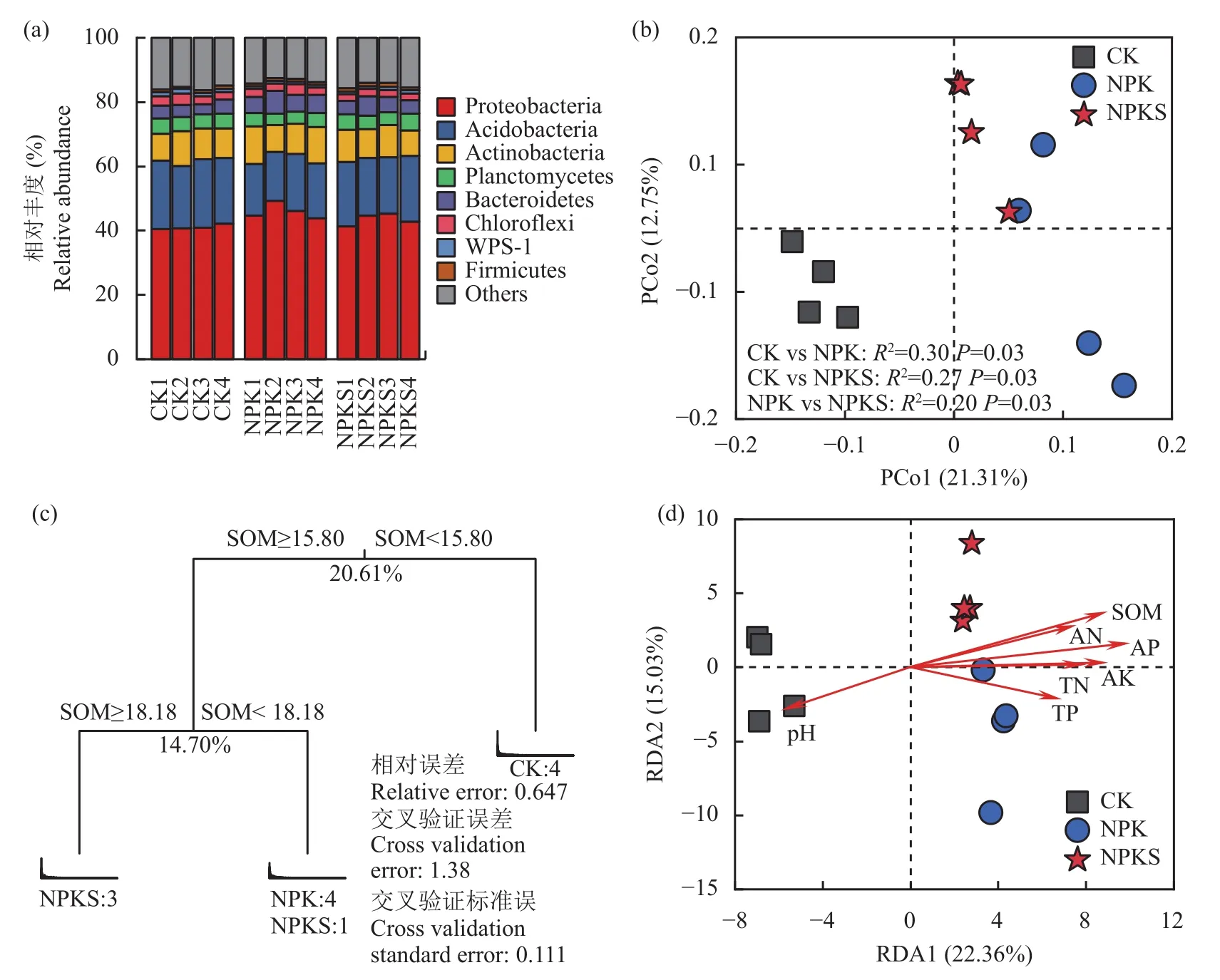
圖3 長期不同施肥處理下土壤的細菌群落結構Fig.3 Soil bacteria community structures under different long-term fertilizations
采用多元回歸樹分析(MRT)和冗余分析(RDA)檢驗細菌群落結構與環境因子間的關系(圖3c, d)。結果發現MRT模型總共解釋了35.31%的群落變異,基本將3個處理分開(除1個NPKS處理歸屬于第二片葉子)。細菌群落首先由土壤有機質(15.80 g/kg)分為不施肥處理(CK)和施用化肥處理(NPK和NPKS)兩大分支,然后由土壤有機質 (18.18 g/kg)又分為單施化肥(NPK)和秸稈還田配施化肥(NPKS)兩大分支。與MRT結果相似,RDA分析(圖3d)結果同樣發現土壤有機質是導致NPK和NPKS處理細菌群落變異的最主要因子。以上結果表明長期施用化肥及秸稈可明顯改變細菌群落結構,而這主要是由于土壤有機質含量的升高造成的。
在ASV水平上對細菌群落進行指示種分析(圖4a和圖4b),結果發現在CK、NPK和NPKS處理中分別有141、49和38個指示種,其中CK處理指示種中酸桿菌門、浮霉菌門相對豐度在3個處理中最高;NPK處理中變形菌門、放線菌門和擬桿菌門相對豐度在3個處理中最高,而NPKS處理中厚壁菌門相對豐度在3個處理中最高。屬水平上的線性判別分析(LEfSe)發現(圖4c),CK處理中相對豐度顯著升高的物種主要歸屬于酸桿菌門的Acidobacteria_Gp3綱,歸屬于變形菌門的生絲微菌科(Hyphomicrobiaceae)和伯克氏菌科(Burkholderiaceae);NPK處理中相對豐度顯著升高的物種主要歸屬于變形菌門的草酸桿菌科(Oxalobacteraceae)、紅丹桿菌科(Rhodanobacteraceae)、黃單胞菌科(Xanthomonadaceae);NPKS處理中顯著升高的物種主要歸屬于厚壁菌門的芽胞桿菌科(Bacillaceae)。
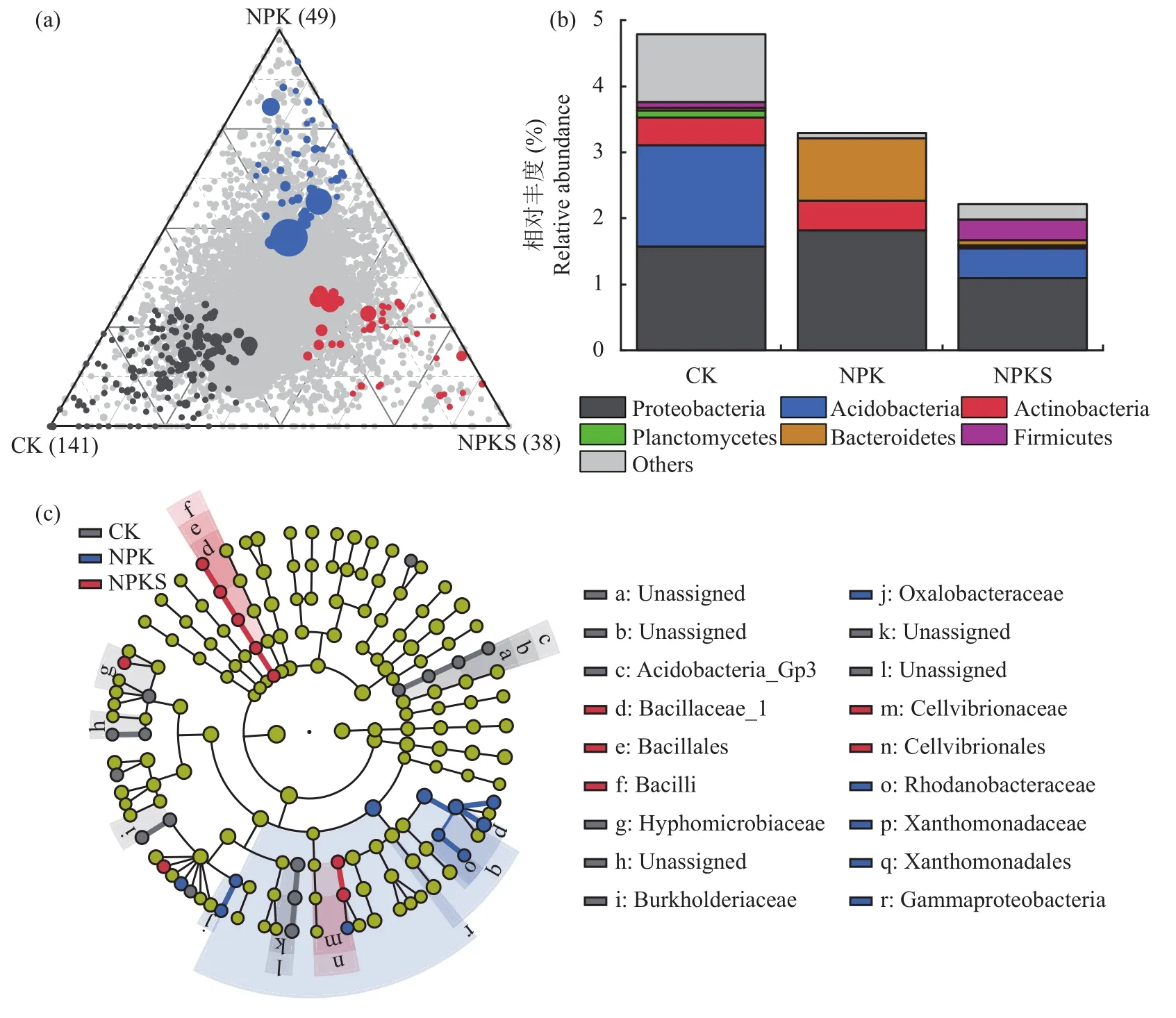
圖4 長期不同施肥處理下土壤細菌的群落組成Fig.4 Soil bacteria community composition under different long-term fertilizations
2.5 土壤細菌分子生態網絡
通過構建細菌分子生態網絡,研究不同施肥模式下土壤細菌物種間互作關系,結果如圖5a所示。圖中每一個節點代表一個ASV,各處理指示種采用不同顏色節點標識,兩個節點間的連線表示存在顯著相關關系。通過對網絡進行模塊(相同模塊內物種高度相連,與其它模塊物種關系較少)分析,發現整個網絡由4個主要模塊組成:CK處理中的指示種主要分布在模塊1和模塊2中;NPK處理中的指示種主要聚集在模塊3;而NPKS處理中的指示種集中于模塊4。對4個模塊內的物種累計豐度進行統計(圖5b),結果發現模塊1和模塊2內物種累計豐度在CK處理中最高,較NPK處理分別升高136.61%和69.70%,較NPKS處理分別升高76.95%和103.74%;模塊3內物種累計豐度在NPK和NPKS處理中分別是CK處理的3.57和2.57倍;模塊4內物種累計豐度按照CK<NPK<NPKS的順序升高,其中NPKS處理內物種累計豐度分別較NPK和CK處理提升52.69%和269.57%。
相關性分析結果(圖5c)表明,模塊1內物種累計豐度與土壤有機質、全氮、全磷、堿解氮、有效磷、速效鉀、β-葡糖苷酶、堿性磷酸酶等生化指標顯著負相關,與pH呈顯著正相關;模塊2內物種累計豐度與土壤有機質、全氮、堿解氮、有效磷、速效鉀、β-葡糖苷酶、堿性磷酸酶和脫氫酶等生化指標顯著負相關;模塊3內物種累計豐度與土壤有機質、全氮、全磷、速效磷、速效鉀和堿性磷酸酶等指標呈顯著正相關,與pH呈顯著負相關;模塊4內物種累計豐度與土壤有機質、全氮、堿解氮、速效磷、速效鉀、β-葡糖苷酶、堿性磷酸酶和脫氫酶等指標呈顯著正相關,與pH呈顯著負相關。以上結果說明不同施肥模式可通過改變土壤養分含量,調節細菌物種網絡互作關系,促進特定微生物集群的生長,從而改變土壤碳、磷循環轉化胞外酶活性。
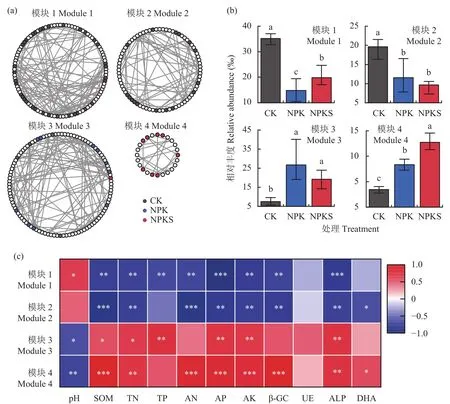
圖5 長期不同施肥處理下土壤細菌的分子生態網絡Fig.5 Soil bacterial co-occurrence patterns under different long-term fertilizations
3 討論
3.1 連續施用化肥及秸稈還田對土壤細菌群落的影響
微生物多樣性是土壤質量和肥力評價的重要依據[3]。然而,在本研究中,長期施用化肥(NPK)和化肥配合秸稈處理(NPKS)雖然提高了土壤有機質含量和全量及速效氮磷鉀含量,但細菌多樣性分別降低0.90%和0.91%,均勻度均降低1.11%。根據微生物對碳底物利用能力和生存策略的不同,可分為富營養型物種(R-策略,Copiotrophs)和寡營養型物種(K-策略,oligotrophs)。富營養型物種適宜在養分含量和有效性高的土壤中生長,優先利用簡單有機碳,表現出相對較快的生長速度;相反地,寡營養型物種則偏向于養分含量和有效性低的土壤,可分解利用頑固有機碳,具有較慢的生長速率[20-21]。因此施用化肥后土壤有機質和有效磷等養分的升高可能會淘汰環境中的K-策略物種,導致細菌多樣性降低。Sun等[16]同樣發現砂姜黑土中土壤肥力與細菌群落變化呈相反趨勢,并認為微生物多樣性的變化與土壤肥力的變化并不一致。我們的研究發現細菌群落多樣性與多個土壤養分指標呈顯著負相關關系,表明土壤中養分的富集是細菌多樣性降低的重要因素。由于大多數微生物在土壤中占據特有的生態位并執行不同生態功能,因此微生物多樣性的降低通常意味著功能的損失[22]。然而本研究發現細菌多樣性與土壤β-葡糖苷酶和堿性磷酸酶等胞外酶呈顯著負相關關系,說明即便是在生物多樣性較低(相對于自然生態系統)的農田生態系統中,土壤微生物仍存在較強功能冗余現象。土壤生態系統中每種代謝功能,可由多個分類地位不同但可以共存的物種執行,因此物種多樣性的降低并不會影響微生物群落執行特定的生態功能。
長期不同施肥處理導致細菌群落組成發生明顯改變,CK處理土壤富集的群落大多歸屬于酸桿菌門(Acidobacteria_Gp3綱)、浮霉菌門、生絲微菌科和伯克氏菌科。其中酸桿菌門是典型的寡營養型物種,廣泛分布在養分貧瘠和退化土壤中,其相對豐度與土壤有機質含量呈顯著負相關[23]。浮霉菌門則被認為是一種溫和的寡營養型物種(moderate oligotrophs),對不同的環境具有廣泛的適應性[24],且在秸稈還田處理后豐度顯著降低[13]。一個有趣的現象是歸屬于變形菌門(通常被認為是富營養型物種)的生絲微菌科和伯克氏菌科同樣在CK處理中廣泛富集,研究表明生絲微菌科和伯克氏菌科均具有較強的固氮功能,可吸收利用大氣中N2,這說明固氮微生物可能在全氮和堿解氮含量較低的CK處理中具有較強的競爭力[25]。NPK處理中變形菌門(草酸桿菌科、紅丹桿菌科和黃單胞菌科)、放線菌門和擬桿菌門相對豐度顯著升高。其中草酸桿菌科是重要的幾丁質溶解細菌[26];紅丹桿菌科和黃單胞菌科均屬于黃單胞菌目,可以分解木質素作為碳源生長[27];而放線菌門和擬桿菌門均具有較強的分解土壤中復雜大分子有機質的能力[28]。與前人研究結果[29]相同,長期秸稈還田處理中富集大量厚壁菌門(芽胞桿菌科),芽胞桿菌科中的許多物種可以分泌纖維素酶,參與秸稈中纖維素的降解。由以上分析可以看出土壤有機質主導了潮土細菌群落結構和組成,這與位于河南鄭州[12]和封丘[30]的潮土施肥定位試驗結果相同。長期不施肥處理中富集了大量養分需求低的寡營養型細菌和固氮菌;單施化肥處理中由于C∶N較低,為了獲得碳源,一些利用頑固碳組分的細菌類群大量富集;而常年秸稈還田配施化肥為土壤提供充足易利用碳源,激發了土壤中具有纖維素降解功能的芽胞桿菌科相關物種。
3.2 連續施用化肥及秸稈還田對土壤細菌分子生態網絡的影響
Li等[12]通過網絡分析發現,長期不同施肥后土壤中富集的物種間存在強烈的合作關系,且與土壤有機質含量呈顯著相關關系,并認為有機無機肥配施可通過調節微生物種間關系,影響土壤有機碳累積。Fan等[31]發現長期施肥抑制了固氮微生物集群(模塊)的生長,導致土壤固氮功能的降低,說明施肥對土壤生態功能的調控可能是通過影響特定微生物模塊的生長。本研究中,我們將3種施肥方式下所有細菌物種進行網絡構建,并對不同施肥模式下的指示物種在網絡中的分布模式進行研究。發現施肥方式不同,網絡中的指示種通過強烈的共生關系聚集在不同的模塊中。模塊1和模塊2主要包含CK處理的優勢菌群,且均與土壤養分指標、β-葡糖苷酶和堿性磷酸酶呈顯著負相關,說明長期施用化肥及秸稈還田降低了這兩個模塊內的群落豐度,淘汰了那些占據低養分含量生態位,且碳、磷降解能力差的群落。模塊3主要包含NPK施肥模式的指示種,與多個土壤養分指標正相關,但與β-葡糖苷酶相關性不高,說明該模塊內的物種可能降解纖維素的能力不強。模塊4內聚集了大量NPKS處理的指示種,且與土壤有機質、全氮、堿解氮、有效磷、速效鉀、β-葡糖苷酶、堿性磷酸酶和脫氫酶等指標呈顯著正相關,說明長期秸稈還田配施化肥刺激該模塊內物種生長,這些微生物通過強烈的種間共生關系,形成碳、磷循環功能微生物集群。前人研究結果表明,秸稈還田后土壤微生物豐度的變化導致群落生態功能發生改變[12,32],而本研究結果則進一步說明,長期不同施肥模式下土壤生態功能的改變,不僅與物種個體豐度變化有關,還與微生物種間共生關系密切相關。秸稈還田配施化肥可促進特定網絡模塊內物種的生長,形成強烈的共生關系,從而提高細菌功能和活性。
4 結論
化肥配合秸稈還田不僅提升了土壤肥力,還改變了細菌群落結構和組成,雖然細菌的多樣性較不施肥有顯著降低,但激活了土壤中芽胞桿菌等纖維素降解群落的活性,通過強烈共生關系形成碳、磷循環功能微生物集群,優化了細菌群落結構和組成,提升了土壤碳、磷的循環轉化功能。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
