等養分投入下冬種紫云英比秸稈還田更有效抑制稻田CH4的產生和排放
2022-09-19 09:26:54陳洪儒魯艷紅廖育林萬運帆王開悅張志偉秦曉波
植物營養與肥料學報 2022年8期
關鍵詞:途徑
陳洪儒,魯艷紅,廖育林,王 斌,萬運帆,王開悅,張志偉,3,聶 軍,秦曉波*
(1 中國農業科學院農業環境與可持續發展研究所 / 中國農業科學院農業農村碳達峰碳中和研究中心 /農業農村部農業環境重點實驗室,北京 100081;2 湖南省土壤肥料研究所,湖南長沙 410125;3 南京信息工程大學,江蘇南京 210044)
溫室氣體濃度的增加已經使全球平均地表溫度比工業化前提高1.1℃[1]。甲烷(CH4)是僅次于二氧化碳(CO2)的人為溫室氣體[2],盡管在大氣中的含量遠低于二氧化碳,但CH4吸收熱紅外輻射的效率要高得多,在100年時間尺度上,其單位質量的全球變暖潛力(GWP)是CO2的28倍。水稻是全球最重要的糧食作物之一,水稻種植的淹水環境促進了CH4的大量排放。我國是世界上最大的水稻生產國,稻田CH4總排放量的估算值為7.7 Tg/a, 約占世界稻田CH4總排放量的30%[3]。同時我國還是世界上最大的氮肥生產國和消耗國,化肥的不合理施用促進了農田溫室氣體大量排放,也加劇了農業面源污染。2015年我國農業農村部明確提出“一控兩減三基本”的目標,控制農業灌溉用水,減量化肥和農藥使用,對畜禽糞便、農膜、作物秸稈進行綜合循環再利用。研究表明,化肥減量配施有機肥能夠促進水稻增產、提高氮肥利用率,不同種類有機物料對稻田甲烷排放影響的研究結果不盡相同[4-5]。水稻秸稈和冬種綠肥是我國南方稻田重要的還田有機物料,含有豐富的碳、氮元素,有利于減少化肥施用、增加土壤養分、改善生態環境,實現農田可持續利用[6-7]。關于綠肥與水稻秸稈還田對稻田CH4排放的影響已有較多研究,一般而言水稻秸稈還田會增加CH4排放[4,8],而綠肥所產生的影響還不確定。
稻田CH4排放是土壤厭氧區域產生的CH4經過氧化和傳輸的凈效應。在淹水稻田中,一系列微生物經過水解、發酵等過程將復雜有機物礦化降解成簡單小分子化合物,被產甲烷菌利用生成終端產物CH4[9-11]。CH4主要由乙酸發酵和氫氣還原二氧化碳(H2/CO2)兩種途徑產生,其中乙酸發酵途徑通常被認為占生物產CH4總量的2/3左右[12-13]。但在不同的稻田土壤中,兩種途徑對產甲烷的貢獻率并非固定不變,實際測定結果和理論值的差異主要取決于土壤微生物種群和土壤有機物料種類的差異[14]。稻田土壤中嗜甲烷菌多分布于有氧氣釋放的水稻根際、土壤-水界面和淹水溶氧層,只使用CH4作為碳源和能源,稻田土壤厭氧區產生的90%以上的CH4可被有氧區的甲烷氧化菌再氧化[12]。CH4從土壤向大氣傳輸主要包括兩種途徑:土壤—水稻植株—大氣途徑和土壤—田面水—大氣途徑,其中植株傳輸在總CH4排放中占絕大部分[15]。穩定碳同位素自然豐度法有助于了解稻田CH4排放的主要控制過程,這是由于稻田土壤產生的CH4以不同傳輸途徑向大氣排放過程中會發生明顯的碳同位素分餾,12CH4較13CH4能更快地被傳輸而釋放到大氣中,CH4產生、氧化和傳輸中CH4的δ13C值具有明顯差異。另外,利用該方法,還可通過觀測CH4和CO2的δ13C值估算乙酸發酵和H2/CO2途徑對總CH4生成的相對貢獻[15-16]。
目前關于化肥與綠肥或水稻秸稈還田結合的不同模式對稻田CH4季節排放規律的研究較多,但對其影響CH4排放機制,尤其是在等養分投入下的節肥減排效應研究還有待加強,另外,關于稻田CH4產生、氧化、傳輸的不同特征及δ13C特征的研究也較少。本研究以超級稻為研究對象,設置等養分投入對比試驗,主要利用培養試驗、大田監測和穩定同位素等手段,研究全量還田下冬種紫云英和水稻秸稈的節肥減排效應。以厭氧培養試驗模擬稻田原位厭氧土層環境,研究綠肥與秸稈處理對稻田CH4產生潛力和氧化潛力的影響,以及兩種關鍵產CH4途徑的貢獻率,揭示冬種紫云英的節肥和CH4減排機制。
1 材料與方法
1.1 試驗點概況
試驗于2021年在湖南省農業科學院高橋試驗基地進行,該地位于季風氣候區,年均溫約16.8℃,年降水量約1400 mm,主要集中在春、夏兩季。試驗前及試驗后各處理供試土壤的基本理化性狀見表1。

表1 試驗地土壤基本性質Table 1 Basic soil properties in this experimental field
1.2 試驗設計
試驗共設6個處理:1) 不施肥(CK);2) 單施化肥(CF);3) 化肥+秸稈全量還田(S);4) 化肥+紫云英全量還田(M);5) 化肥+秸稈和紫云英全量還田(MS);6) 化肥+秸稈和紫云英全量還田+熟石灰(MSC)。供試紫云英品種為‘湘紫1號’,紫云英處理的小區于上季晚稻收獲后(2020年10月14日)播種,2021年4月中旬盛花期收割稱重測產,重復小區紫云英均量翻壓還田,紫云英鮮草還田量為22500 kg/hm2。秸稈處理的小區于上季水稻收獲后稱重測產,重復小區均量覆蓋還田,稻草還田量為2625 kg/hm2。供試紫云英和稻草的養分含量見表2。MSC處理于水稻移栽前(2021年6月6日) 重復小區撒施熟石灰3.4 kg并旋耕。除CK處理外,各施肥處理養分投入量相同,均為N 210 kg/hm2、P2O575 kg/hm2、K2O 120 kg/hm2,各處理養分投入量詳見表3。施用氮、磷、鉀肥的種類分別為尿素(含N 46%)、鈣鎂磷肥(含P2O512%)、氯化鉀(含K2O 60%)。磷肥在移栽前(2021年6月7日)作基肥施入;氮肥和鉀肥50%作基肥,50%作追肥在分蘗期施入。小區面積30 m2,3次重復,共18個小區,隨機區組排列。供試超級稻品種為秈型兩系雜交稻‘晶兩優華占’,于2021年6月9日移栽。除中期曬田外,其他時期為間歇灌溉,水稻收獲前10天自然落干。其他田間管理措施按當地常規方法進行。

表2 供試紫云英和稻草的養分含量Table 2 Nutrient contents of the tested Chinese milk vetch and rice straw

表3 不同處理氮、磷、鉀養分和熟石灰投入量(kg/hm2)Table 3 Nutrient and slaked lime input amount in different treatments
1.3 樣品采集及測定
1.3.1 稻田CH4排放通量測定 采用擁有專利權的密閉式靜態暗箱[17],采樣箱由底座和透明頂箱組成。頂箱高度為100 cm,長寬均為37 cm,側壁包有反光隔熱膜以防止采樣期間箱內溫度變化過大。各小區底座于生長季埋于田間,底座內罩1株水稻植株。取氣時間為水稻分蘗初期的2021年6月21日上午9:00—10:00,此時紫云英還田72天,秸稈還田250天,水稻生長較旺盛,CH4排放相對較高。取氣時將頂箱垂直安放在底座上部5 cm深的凹槽內并加水密封,使用溫室氣體自動采樣器分別在扣箱后第0、10、20、30 min采集氣體收集于500 mL鋁箔氣袋中,密封恒溫保存。為防止人為擾動造成誤差,在田間搭設木橋通往各小區底座。氣樣中CH4濃度和δ13C-CH4由氣體濃度/同位素分析儀(Picarro G2201-i,USA)測定。CH4排放通量由箱中濃度隨時間的變化率計算得出:

式中:F為CH4排放通量,mg/(m2·h);ρ為標準狀態下CH4的密度,取值為0.714 kg/m3;h為采樣箱高度,m;dc/dt為單位時間采樣箱內CH4濃度變化率,mL/(m3·h);T為采樣時箱內溫度,℃。
1.3.2 田面水溶存CH4采集與分析 用靜態箱取氣的同時,使用60 mL注射器在每個底座內取田面表層水(無氣泡)。于實驗室內將各注射器中水樣排出至剩余水樣30 mL,將30 mL高純氦(純度>99.999%)注入各注射器。室溫下將注射器置于振蕩器震蕩5 min,靜置10 min,使注射器內達到氣液兩相平衡。最后將注射器上部30 mL氣體轉入12 mL真空瓶內,用氣相色譜儀(Agilent 7890A,USA)測定氣樣中CH4濃度。田間操作及氣樣的測定分析詳見文獻[18-19]。
1.3.3 土壤樣品采集與分析 培養試驗土壤采集自田間取氣和田面水取樣當天(2021年6月21日)。在每個小區按“S”形隨機取0—20 cm土樣組成一個混合樣,剔除石礫和植物殘體,所取土樣隨即分成2份裝入密封袋帶回實驗室冷凍保存,一份用于測定理化性質,一份用于室內培養實驗。供試土壤有機質(SOM) (NY/T 1121.6—2006)、全氮(TN) (LY/T 1228—2015)、全碳(TC)[20]在湖南省土壤肥料研究所農化檢測中心測定,可溶性有機碳(DOC)[21]和可溶性總氮(TDN) (HJ 667—2013)在中國科學院亞熱帶農業生態研究所測定。
1.3.4 CH4產生與氧化潛力培養試驗 甲烷產生潛力厭氧培養試驗:1) 根據折合干重20 g和土壤含水量計算水土比為2∶1時的濕土重和加水體積,每個處理取3個重復小區混合濕土樣稱取相應濕土重,置入100 mL培養瓶,加入去離子水,攪拌使土樣分散混合均勻,密封瓶口,每個處理4次重復;2) 在30℃黑暗條件下進行10天預培養,預培養結束后進行反復幾次抽真空和高純氮氣吹掃循環,最后充滿氮氣至常壓,再于30℃黑暗條件下在大氣壓下緩慢平衡24 h以消耗培養瓶中的殘留氧氣;3) 在0 h取氣,取氣之前將培養瓶輕輕水平旋轉1~2 min,注射器取頂部30 mL氣樣轉入12 mL真空瓶中,取氣后補充30 mL高純氮氣至常壓,放入30℃黑暗培養箱繼續培養48 h再次取30 mL氣樣。
甲烷氧化潛力好氧培養試驗:1) 根據折合干重20 g和土壤含水量計算土壤含水量為70%時的濕土重和加水體積,每個處理取3個重復小區混合濕土樣稱取相應濕土重,置入100 mL培養瓶,加入去離子水,密封瓶口,每個處理4次重復;2) 將培養瓶放入30℃黑暗培養箱中預培養24 h;3) 用微型注射器將1 mL純CH4注入培養瓶中以形成較高的內部濃度,30℃條件下120 r/min振蕩24 h。在0、24、48、72 h取頂部30 mL氣樣轉入12 mL真空瓶中,取氣之前將培養瓶輕輕水平旋轉1~2 min,每次取完氣補充30 mL新鮮空氣。
取樣完成后用氣相色譜儀測定CH4氣體濃度,計算甲烷產生/氧化潛力:

式中:P為 CH4產生/氧化潛力 [μmol/(mol·d)];dc/dt為培養瓶上空CH4濃度單位時間變化率[μmol/(mol·d)];V為培養瓶內氣體體積(L);MV為標準狀態下1 mol氣體的體積(22.4 L);MW為CH4的分子量(g);W為干土重(g);T為培養溫度(K)。
1.3.5 δ13C-CH4與關鍵產CH4途徑 用氣體濃度/同位素分析儀(Picarro G2201-i,USA)測定田間排放、田面水溶存和厭氧培養試驗產生氣體的δ13C-CH4和δ13C-CO2,根據CH4和CO2同位素標氣擬合曲線計算樣品氣體中δ13C的實測值。穩定碳同位素組成(‰)利用公式(3)計算:
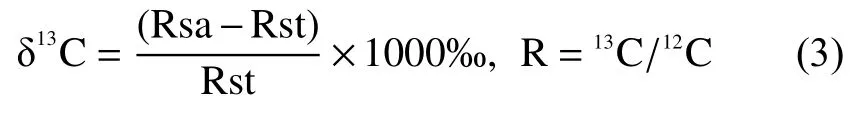
式中:Rsa和Rst分別表示待測樣品和標準樣的同位素比值;13C/12C對應于國際標準ViennaPDB,分析精度為1‰。
采用簡單混合模型(公式4、5)計算乙酸發酵和二氧化碳還原兩種關鍵產CH4途徑占全部CH4的比例[22-23]:

式中:δ13C-CH4是測定的全部產生途徑的CH4碳同位素值;δ13C-CH4(CO2)為CO2還原途徑產生的CH4碳同位素值(利用公式4計算),CO2分餾系數 αCO2一般采用某一固定值,本研究使用1.079來計算[22],該系數在一個純培養研究中確定,其中CO2還原是天然濕地中CH4產生的唯一途徑[24],已應用于確定湖泊沉積物中的CH4產生途徑[22,25];fCO2和face分別為二氧化碳還原和乙酸發酵2個途徑占全部CH4的比例 (假設 fCO2+face=1);δ13C-CH4(ace)為乙酸發酵途徑產生的CH4碳同位素值,在新鮮有機質充足的假設下,其值為-43‰,而在新鮮有機質極度消耗的假設下其值為-27‰[25],本研究使用二者平均值-35‰來計算。
1.4 數據處理
使用微軟Excel (v.2010, Microsoft)整理并初步計算數據均值和標準誤差;用SPSS (v.23.0, IBM)進行統計分析,通過單因素方差分析(ANOVA)識別不同施肥處理間稻田CH4排放通量、田面水溶存CH4濃度、稻田土壤CH4產生潛力和氧化潛力的差異,用最小顯著性檢驗(LSD)法比較不同處理間的差異顯著性(P<0.05),并對各觀測變量做Spearman相關分析,在0.05水平和0.01水平檢驗相關系數顯著性;用GraphPad Prism (v.7, GraphPad Software)和R程序[26]繪圖。
2 結果與分析
2.1 稻田土壤培養過程中CH4濃度變化
在厭氧培養的48 h內,各處理CH4濃度均一致呈現升高的趨勢(圖1A)。S處理的CH4濃度及提升幅度最大,遠高于其他處理。M處理的CH4濃度和提升幅度均最小,甚至低于CK。MS和CF的CH4濃度和提升幅度十分接近,而MSC的提升幅度大于CK。
相反,在好氧培養的72 h內,各處理CH4濃度均隨培養時間延長而下降(圖1B)。CK處理的CH4濃度下降幅度最大,其他處理的CH4濃度下降幅度非常接近。

圖1 厭氧培養(A)和好氧培養(B)過程中各處理CH4濃度變化Fig.1 Variation of CH4 concentration during anaerobic incubation (A) and aerobic incubation (B)of different treatments
2.2 稻田土壤CH4產生潛力和氧化潛力
厭氧條件下,不同處理稻田土壤CH4產生潛力范圍為 0.0039~0.0427 μmol/(mol·d),無統計學上顯著差異(圖2A)。S、MS和MSC處理的CH4產生潛力均高于單施化肥(CF)和不施肥(CK)處理。M處理的CH4產生潛力最低,比其它5個處理平均低61.04%,比S處理低83.16%。
好氧條件下,不同處理稻田土壤CH4氧化潛力范圍為 2.75~3.85 μmol/(mol·d),CK 處理的 CH4氧化潛力顯著高于所有施肥處理(P<0.05) (圖2B)。CK、CF、MS、MSC處理的CH4氧化潛力大小順序與CH4產生潛力相反。M處理的CH4氧化潛力最低,比其它5個處理平均低7.56%,比S處理低5.36%。

圖2 各處理CH4產生潛力(A)和氧化潛力(B)Fig.2 CH4 production potential (A) and oxidation potential (B) of different treatments
2.3 不同處理稻田土壤CH4關鍵產生途徑
CH4產生潛力厭氧培養期間首末兩次取的CH4均以H2/CO2還原產生途徑為主導(圖3)。培養0 h各處理乙酸發酵產CH4途徑所占比例范圍為9.93%~32.56%,培養48 h后各處理乙酸發酵產CH4途徑所占比例范圍為22.31%~50.46%,因此相較于0 h,培養48 h后各處理CO2還原產CH4途徑所占比例均下降,乙酸發酵產CH4途徑所占比例均增加。此外,培養期間各處理乙酸發酵產CH4途徑比例平均值大小順序為 MS>S>CK>CF>MSC>M,M 處理乙酸發酵產CH4途徑比例最低,比其他5個處理平均低52.52%,分別比S、MS、MSC處理低53.49%、61.17%、45.65%。

圖3 各處理乙酸發酵途徑和CO2還原途徑產CH4比例Fig.3 The proportion of CH4 produced by AM and HM of different treatments
2.4 稻田CH4排放通量和田面水溶存CH4濃度
不同處理間CH4排放通量范圍為5.70 ~ 26.65 mg/(m2·h),各處理差異不顯著,所有施肥處理相比不施肥(CK)均增加了CH4排放(圖4)。含秸稈還田的3個有機處理的CH4排放通量均高于CF,而沒有秸稈還田的M處理比CF降低了5.91%,相比S、MS、MSC分別低12.50%、15.30%和44.40%。
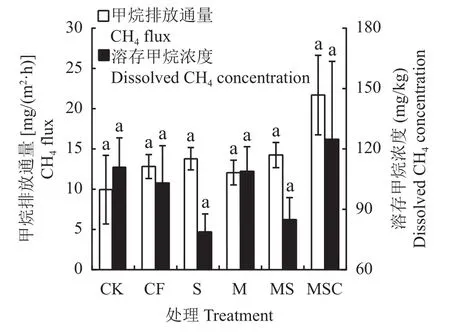
圖4 不同處理稻田CH4排放通量和田面水溶存CH4濃度Fig.4 CH4 flux from rice field and dissolved CH4 concentration from surface water in different treatments
不同處理間田面水溶存CH4濃度范圍為70.02~163.58 mg/kg,各處理差異不顯著(圖4)。MSC處理的田面水溶存CH4濃度最高,比其他處理平均提高30.68%。除MSC處理外,其他施肥處理的溶存CH4濃度均低于CK,相比S和MS處理,M處理的田面水溶存CH4濃度較高。
2.5 δ13C-CH4分布特征
各處理CH4產生、排放和田面水傳輸過程均呈現出一致的δ13C分餾變化趨勢:厭氧培養產生<整體排放<田面水溶存。厭氧培養期為CH4的產生過程,初始期(0 h)各處理δ13C-CH4為-102.73‰~-84.00‰,48 h后為-94.65‰~-72.53‰,δ13C-CH4呈富集趨勢,這可能是由于發生了一定厭氧氧化,消耗了不穩定的12C,使δ13C相對富集(圖5A)。田面水溶存δ13C-CH4值相對產生過程較富集,分布范圍也較大(-57.66‰~-20.24‰) (圖5B),這說明經過土壤氧化及田面水中的好氧氧化,發生了同位素分餾和δ13C-CH4富集。田間整體排放的δ13C-CH4值(-66.38‰~-61.83‰)同樣相對產生過程較富集(圖5B),說明土壤產生的CH4經過氧化和傳輸過程的同位素分餾,最終排放出的12C-CH4較產生過程呈貧化的趨勢。

圖5 不同處理CH4產生(A)、排放和田面水溶存(B)過程的δ13C-CH4特征Fig.5 Characteristics of δ13C-CH4 of production (A), emission and soluble state (B) of different treatments
2.6 環境要素對CH4產生和排放的影響
各處理甲烷產生、排放與各環境要素間的相關性分析表明,CH4排放通量與CH4產生潛力呈正相關,與CH4氧化潛力呈負相關,CH4產生與排放受到土壤理化因子的綜合影響(圖6)。CH4排放通量、CH4產生潛力與土壤TDN含量呈顯著正相關(P<0.05),各處理TDN含量大小順序為MSC>S>MS>CF>M>CK (表1),所有施肥處理TDN含量高于CK,與CH4排放通量規律一致。CH4產生潛力與土壤DOC含量呈顯著正相關(P<0.05),其中M處理的DOC含量分別比S、MS、MSC處理低25.99%、1.54%、21.18%,與CH4產生潛力變化趨勢一致。另外,土壤C/N與CH4排放通量、CH4產生潛力均呈負相關性,各處理土壤C/N大小順序為M>CK>MS>S>MSC>CF (表1)。綜上可見,土壤TDN、DOC和C/N與CH4產生和排放過程密切相關。

圖6 各觀測變量相關矩陣Fig.6 Correlation among different measurement indexes
3 討論
3.1 冬種紫云英減施化肥與CH4減排效應
綠肥是農作物生產的重要物質基礎,是綠色農業的有效技術支撐,數據表明,綠肥能有效促進作物養分供給、保障作物穩產高產、全面改善土壤物理、化學、生物性狀以及減少溫室氣體排放[27]。本研究結果表明,冬種紫云英具有較好的氮肥減施和甲烷減排效應。相比單施化肥(CF)處理,化肥+紫云英全量還田(M)減施化肥N 57.01 kg/hm2,節省氮肥27.15% (圖7),而相比化肥+秸稈全量還田(S)處理,也能減施13.78%的N,折合化肥N 24.46 kg/hm2。另外,M處理的CH4排放通量低于CF和其他幾種組合處理,尤其比S處理低12.50%,表明冬種紫云英可在替代氮肥的同時減少超級稻田CH4排放。田卡等[8]對華南雙季稻田的研究表明,等化肥投入下秸稈還田相比單施化肥提高了水稻幼穗分化前稻田CH4排放通量,而冬種綠肥影響不大。張麗[28]在廣州雙季稻田的研究發現,等化肥投入下秸稈還田比冬種紫云英提高了早稻和晚稻CH4排放通量,但相比單施化肥,紫云英、秸稈還田及二者協同還田均顯著增加了雙季稻CH4排放。另外,劉威[29]研究了秸稈不同還田方式對江漢平原單季中稻CH4排放通量的影響,發現化肥+秸稈覆蓋還田、化肥+紫云英還田處理的CH4排放通量高于不施肥、單施化肥和化肥+秸稈焚燒還田處理。這些研究結果的差異可能是由于研究區域氣候、土壤質地、施氮量、研究年限等不同導致的,另外,以往研究多是基于等氮肥基礎上添加不同有機物料,這與本研究等養分投入的設定有差異。
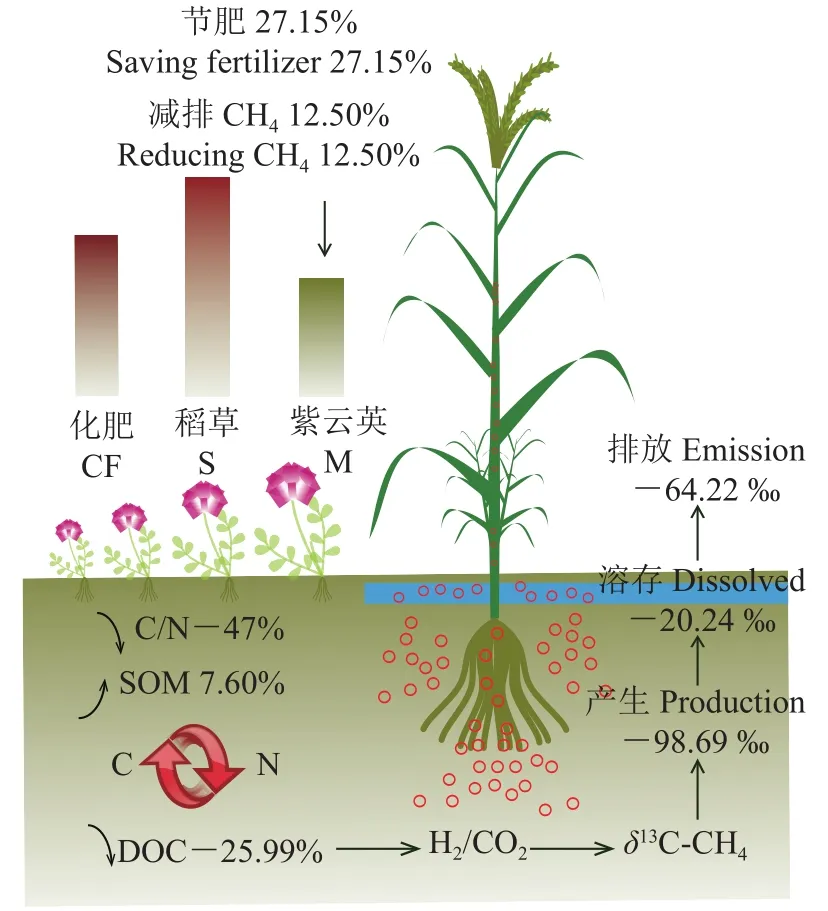
圖7 冬種紫云英節肥和CH4減排效應示意圖Fig.7 Schematic diagram of the effects of winter-planting Chinese milk vetch on nitrogen input reduction and CH4 mitigation
本研究發現,化肥+紫云英全量還田(M)處理的CH4產生潛力最低,比其他5個處理平均低61.04%,分別比CF和S兩個處理的CH4產生潛力低了62.29%和83.16%,表明了冬種紫云英有較強的CH4產生抑制效應。紫云英還田也抑制了部分CH4氧化能力,相比CF和S處理,CH4氧化潛力分別降低了4.53%和5.36%但差異不顯著(P>0.05),這說明單獨紫云英還田對土壤微生物群落特別是產甲烷菌和嗜甲烷菌分別具有抑制和激發效應,但對CH4產生的抑制明顯強于對CH4氧化過程的激發,因此最終顯示出對稻田甲烷整體排放的抑制效應。Zhou等[30]的培養試驗得出相似的結論,發現添加秸稈殘茬的處理CH4產生潛力和氧化潛力均高于添加綠肥殘茬處理。實際上,綠肥與稻秸投入直接改變了土壤養分狀況,間接改變了產甲烷菌和嗜甲烷菌群落結構和活性,進而改變了土壤CH4產生和氧化潛力,最終導致CH4排放差異[30-31]。本研究相關分析表明,CH4排放通量與CH4產生潛力呈較明顯正相關性,但與CH4氧化潛力負相關性不明顯。這與前人研究結論相似,如Zhou等[30]研究表明,CH4產生潛力而非CH4氧化潛力對CH4排放具有很強的直接影響。
3.2 冬種紫云英CH4減排機制
由于紫云英植株C/N值顯著低于水稻植株,在本研究中,紫云英全量還田處理(M)顯著增加了土壤有機質和總碳含量,比秸稈全量還田(S)處理分別提高7.60%和7.55% (表1),但該處理顯著降低了土壤活性有機碳(DOC)的含量,相比CF和S處理分別降低了17.45%和25.99%,這就從碳源上抑制了產CH4過程,而更多的碳則儲存于土壤中。本研究的相關分析表明,各處理CH4產生潛力與土壤DOC含量呈顯著正相關,進一步證實了紫云英對于土壤碳周轉和CH4產生與排放的影響。多位學者研究證明CH4排放與土壤DOC呈正相關,DOC增加促進CH4排放,原因在于淹水條件下壯大的厭氧及兼性厭氧微生物群落能夠對土壤DOC進行無氧發酵,從而增加土壤CH4產生與排放。另外,研究表明,隨有機物料殘茬C/N增加,土壤CH4排放隨之增大[32-35],因此本研究中不同C/N的紫云英和秸稈因自身分解差異導致土壤DOC含量差異,進而造成紫云英比秸稈還田顯著抑制稻田CH4產生與排放,原因可能在于本試驗中紫云英于水稻移栽前兩個月翻壓還田,經過充分的好氧預分解過程促進了土壤活性有機碳徹底分解為CO2,導致在水稻移栽之后的淹水土壤中紫云英處理的土壤可利用碳較低,CH4產生被強烈抑制;相比水稻秸稈,紫云英C/N明顯更低(低47.04%,表2),水稻秸稈含有木質素等更多難以降解的物質,分解更緩慢且不完全,在試驗淹水期間土壤DOC含量高于其他處理,刺激了CH4產生。
此外,本研究還發現,CH4排放通量、CH4產生潛力也與土壤可溶性總氮(TDN)含量呈顯著正相關,所有施肥處理CH4排放通量均高于不施肥處理(CK),而且各處理相比CK均提高了土壤TDN含量,Yu等[36]研究也發現有機物料連續施用10年的稻田土壤TDN顯著增加。相比化肥+秸稈全量還田處理(S),化肥+紫云英全量還田處理(M)的總氮含量增加了1.82%,而TDN含量下降了19.95% (表1),可見冬種并翻壓紫云英降低了水稻生育早期土壤有效態氮,可能減少了產甲烷菌氮源進而抑制其生長和活性,從而抑制了CH4產生與排放。
本研究發現,土壤C/N與CH4通量和CH4產生潛力均呈負相關性,化肥+紫云英還田提高了土壤C/N (促進提高了碳固存),抑制了CH4產生與排放,反之,化肥+秸稈還田降低了土壤C/N,促進了CH4產生與排放。這可能是由于低C/N的紫云英還田后易于被微生物消耗分解,短期內有多余的氮進行氮礦化,有利于土壤中可利用氮的增加,可直接為植物所吸收,土壤C/N相對提高。高C/N的秸稈含氮量低,等氮條件下比紫云英還田需配施更多氮肥,氮輸入能降低土壤C/N,促進土壤有機質分解,增加土壤活性碳含量,促進CH4產生。
另外,本研究發現化肥+秸稈和紫云英全量還田+熟石灰處理(MSC)的稻田CH4排放通量和田面水溶存CH4濃度最高(圖4),且CH4產生潛力僅低于S處理。研究表明,土壤pH可能通過影響有機質分解、土壤C/N和CH4產生過程間接影響CH4排放[37-38]。一方面MSC處理的土壤DOC含量較高,土壤C/N較低(表1),可能因此促進了稻田土壤CH4產生與排放(圖6);另一方面土壤pH的輕微變化會影響土壤CH4排放,尤其是厭氧條件下CH4產生對土壤pH值變化非常敏感,中性或微堿性土壤最有利于CH4產生[38],因此添加熟石灰可能因土壤pH升高而促進了產CH4菌活性。
本研究中,培養試驗土壤取自水稻分蘗初期,研究發現各處理CH4產生過程以H2/CO2還原為主。一些研究也發現冬閑期和水稻生長季大部分時期H2/CO2還原產生途徑非常重要(約50%),僅在7、8月水稻生長旺盛時期,乙酸發酵途徑占主導地位(約70%)[39-40]。我們也發現,隨培養時間延長,乙酸發酵過程的貢獻占比增加(圖3)。本研究中,乙酸發酵產CH4途徑比例平均值[為減少時間誤差,本研究中乙酸發酵產CH4途徑貢獻率采用厭氧培養0和48 h的平均值(圖3)來計算]與相應δ13C-CH4平均值(圖5A)呈極顯著正相關(r=0.943,P<0.01),這可能是由于利于乙酸發酵產CH4的環境中12CH4較快氧化,導致δ13C-CH4值偏正,反之厭氧培養的δ13CCH4值偏負,從而說明此環境利于H2/CO2還原產CH4。另有研究認為,曬田會導致土壤氧化還原電位偏高,乙酸型產甲烷作用在很大程度上優于H2/CO2還原型產甲烷作用[41],這說明氧化作用會使δ13C-CH4值偏正,環境中乙酸型產CH4途徑比例增加,氫營養型產CH4途徑比例下降,這與本研究的結果一致。此外,本研究發現乙酸發酵產CH4途徑比例平均值(圖3)與CH4產生潛力(圖2A)呈極顯著正相關(r=0.628,P<0.01),表明乙酸發酵途徑對CH4產生總量的重要影響,厭氧培養期間紫云英全量還田處理乙酸發酵產CH4途徑貢獻率最低,平均只有16.11%,而秸稈全量還田處理占到34.65%,表明紫云英單獨還田抑制了乙酸發酵產CH4途徑(相比秸稈單獨還田降低53.49%),從而抑制了CH4產生,而水稻秸稈還田可明顯促進乙酸發酵過程,這可能是由于介導兩種途徑的功能微生物類群的差異。
4 結論
甲烷排放通量和產生潛力與土壤可溶性有機碳、可溶性總氮含量呈正相關,與土壤碳氮比呈負相關。在等氮磷鉀投入條件下,冬種并翻壓紫云英減少了化學氮肥投入,降低了土壤可溶性有機碳含量,提高了土壤碳氮比,因而有效抑制了稻田土壤甲烷的產生和排放。秸稈的作用正好相反,因此,秸稈還田需配合紫云英翻壓來抵消甲烷的產生和排放。
厭氧條件下,乙酸發酵產甲烷途徑的比例隨培養時間延長而升高。厭氧培養的乙酸發酵產甲烷途徑比例平均值與甲烷產生潛力呈正相關,紫云英全量還田處理主要抑制了乙酸發酵這一關鍵產甲烷過程,從而減少了甲烷產生,其乙酸發酵產甲烷途徑比例和甲烷產生潛力比其他5個處理平均降低52.52%、61.04%,其中比秸稈單獨還田處理降低53.49%、83.16%。
猜你喜歡
語數外學習·高中版中旬(2023年2期)2023-05-10 13:26:53
語數外學習·高中版中旬(2022年5期)2022-07-13 20:47:51
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學生百科·大語文(2017年10期)2017-11-04 06:56:38
中國衛生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國衛生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03
