兩種池養模式下異育銀鯽(Carassius auratus gibelio)生物學表型對體質量影響效果的差異分析*
2022-09-21 02:15:32戎華南劉哲宇陳漢春王志錚
海洋與湖沼 2022年5期
陳 雨 戎華南, 劉哲宇 陳漢春 王志錚
兩種池養模式下異育銀鯽()生物學表型對體質量影響效果的差異分析*
陳 雨1戎華南1, 2劉哲宇1陳漢春2王志錚1①
(1. 浙江海洋大學水產學院 浙江舟山 316022; 2. 慈溪市水產技術推廣中心 浙江慈溪 315300)
水產養殖動物的增重機制既是其生存對策的重要組成部分, 也是影響其生長性能和養殖產量的重要內因。異育銀鯽()是我國的重要養殖魚類, 隨機選取池塘生態主養模式(M1)和池塘生態套養模式(M2)下經7個月養殖的異育銀鯽夏花苗種各70尾為研究對象, 以全長(1)、體高(2)、體寬(3)、頭長(4)、側線長(5)、尾柄高(6)、腹鰭間距(7)、胸鰭間距(8)為體尺性狀, 凈體質量(NW)、腸質量(1)、肝質量(2)、胃質量(3)、鰓質量(4)、心質量(5)和鰾質量(6)為稱量性狀, 采用多元分析方法分別研究了M1、M2實驗群體尺寸性狀和稱量性狀對其體質量(BW)的影響效應。結果表明: (1) 生物學測定結果顯示, CV值大于10%的均為質量性狀, 小于10%的均為體尺性狀, 除NW、1、5和6均呈M1
養殖模式; 異育銀鯽(); 體尺性狀; 稱量性狀; 體質量
鯽()隸屬于輻鰭亞綱(Actinopterygii)、鯉形目(Cypriniformes)、鯉科(Cyprinidae), 具較高的食用和藥用價值, 系廣泛分布于歐亞大陸的淡水經濟魚類。因具食性廣、生長快、耐低氧、繁殖力高、抗逆和抗病力強、可常年養殖等特點, 而備受廣大淡水養殖從業人員的推崇。據《2020漁業統計年鑒》(農業農村部漁業漁政管理局等, 2020), 全國2019年度鯽養殖產量為2.755 632×106t, 在我國七大大宗淡水魚中位列第五。近年來, 隨著環保督查力度的不斷加大, 構建并推廣銀鯽適宜的池塘生態養殖模式已日益引起業內學者和主產區漁業行政主管部門的高度關注和重視。
水產動物的體質量既是評價其階段生長的重要生物學指標, 也是開展人工選育的重要目標性狀。2002年以來, 最近
二十年間國內學者借助通徑分析方法開展水產養殖動物形態體尺性狀對體質量影響效果的研究一直非常活躍(劉小林等, 2002), 但涉及我國大宗淡水魚類的研究僅見草魚()和鯉魚() (李璽洋等, 2012; 孫俊龍等, 2015; 周洲等, 2017; 周康奇等, 2020), 至于水產養殖動物稱量性狀(組織和臟器質量)與體質量間真實關系的研究報道亦僅見中華鱉(牟恩鏇等, 2021a)。水產養殖動物的增重機制既是其生存對策的重要組成部分, 也是影響其生長性能和養殖產量的重要內因。已有研究表明, 水產養殖對象會隨養殖模式的改變而選擇相應的生存對策, 并隨著養殖時間的推移, 不僅其生長性能和決定體質量的形態體尺性狀組合均會產生較明顯的改變(楊明秋等, 2014; 袁向陽等, 2016, 2017b; 趙旺等, 2018; 原居林等, 2018), 而且其體色、體型、肉質、營養與食用風味等商品性狀也均會受到一定程度的影響(王志錚等, 2012, 2013a, 2013b; 袁向陽等, 2017a; 原居林等, 2018; 趙柳蘭等, 2021)。因此, 運用通徑分析方法從體尺性狀和稱量性狀兩個維度聚焦決定體質量的核心變量組合, 綜合剖析體尺性狀與稱量性狀在決定體質量上的相互關聯和內在邏輯, 進而揭示不同養殖模式下水產養殖動物的體質量增長對策差異, 無疑對于進一步優化各養殖模式的養殖工藝與技術參數具有重要現實意義。鑒于此, 本研究團隊分別以池塘生態主養模式和池塘生態套養模式下經7個月養殖的異育銀鯽夏花苗種為研究對象, 從體尺性狀與稱量性狀兩個維度較系統開展了生物學表型對異育銀鯽體質量影響效果的定量研究, 并以此解析上述2種養殖模式下異育銀鯽的體質量增重機制差異, 以期為指導異育銀鯽的科學養殖提供基礎資料。
1 材料與方法
1.1 實驗魚
本研究所測實驗魚均于2020年12月捕自慈溪市滸山華喬淡水魚養殖場, 分別為池塘生態主養模式(記為M1, 池塘四周密植傘型花科多年生挺水植物銅錢草; 異育銀鯽夏花放養時間2020年5月7日, 放養密度1.5×105ind./hm2; 鳙放養時間2020年5月28日, 規格300 ind./kg, 密度3.0×104ind./hm2)和池塘生態套養模式(記為M2, 凡納濱對蝦苗放養時間2020年5月10日, 放養密度6.0×105ind./hm2; 鳙放養規格300 ind./kg, 異育銀鯽夏花和鳙放養時間均為2020年5月19日, 放養密度分別為6.0×103ind./hm2和3.0×103ind./hm2)下經7個月常規養殖的異育銀鯽夏花苗種。實驗魚運至慈溪市水產推廣研究中心實驗基地經停食馴養1 d后, 選取其中體形完整、鱗片齊全、無病無傷、反應靈敏的健壯個體備測。
1.2 生物學測定
隨機選取M1和M2養殖模式實驗魚各70尾, 逐一編號后按性狀屬性分別用HZ1002A型電子天平(精度0.01g, 慈溪紅鉆衡器設備有限公司出品)和電子數顯游標卡尺(精度0.01 mm, 桂林廣陸數字測控股份有限公司出品)進行生物學測定。具體步驟為: (1) 冰水低溫麻醉并用紗布抹除體表水分后, 逐尾測定體質量(BW)、全長(1)、體高(2)、體寬(3)、頭長(4, 吻端至鰓蓋骨后緣的最大水平距離)、側線長(5, 側線前后端的水平距離)、尾柄高(6, 尾柄部背腹間的最小垂直距離)、腹鰭間距(7, 左右腹鰭間的最小水平距離)和胸鰭間距(8, 左右胸鰭間的最小水平距離), 上述體尺性狀的具體測量部位如圖1; (2) 按常規解剖方法, 摘取腸、肝、胃、鰓、心、鰾并剝離附于其上的脂肪和結締組織, 用吸水紙吸除各臟器表面血液、擦凈解剖后實驗魚體表和內腔后, 逐尾稱量腸質量(1)、肝質量(2)、胃質量(3)、鰓質量(4)、心質量(5)、鰾質量(6)和凈體質量(NW, 去除鰓和內臟后的體質量)。
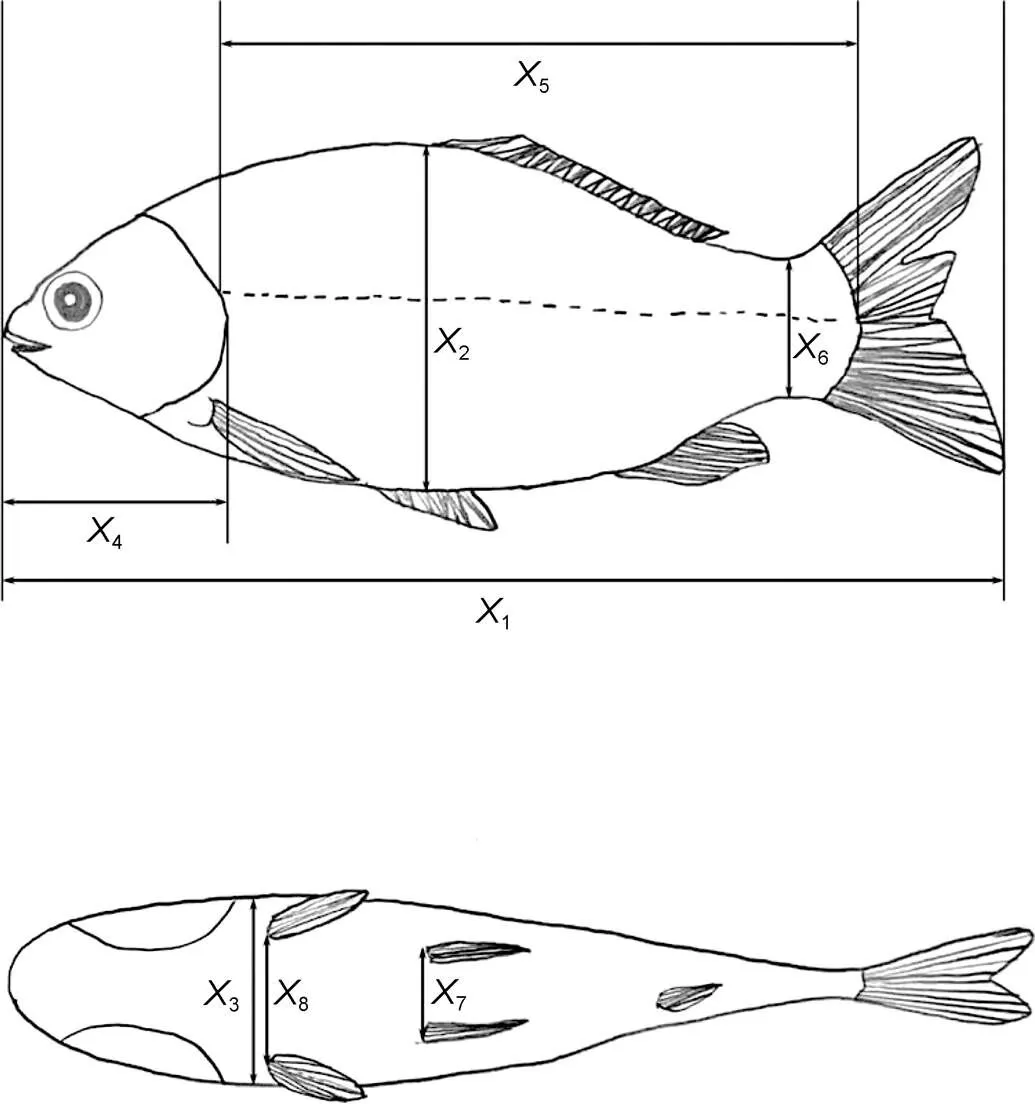
圖1 實驗魚體尺性狀的測量部位
注:1表示全長;2表示體高;3表示體寬;4表示頭長;5表示側線長;6表示尾柄高;7表示腹鰭間距;8表示胸鰭間距
1.3 數據處理
整理測定數據, 計算各項生物學性狀的均值(M)、標準差(SD)和變異系數(CV%), 并比較各臟器系數的組間差異顯著性(<0.05表示差異顯著); 參照牟恩鏇等(2021a, 2021b)的方法, 運用SPSS23.0軟件, 依次開展體尺性狀間、稱量性狀間以及各性狀與體質量間的簡單相關性分析, 體尺性狀和稱量性狀對體質量的通徑分析、復相關分析和偏回歸分析, 由此確定影響體質量的關鍵體尺性狀組合和稱量性狀組合, 并對所建用于估算體質量的多元線性回歸方程進行擬合度檢驗。
2 結果
2.1 參數統計
由表1可見, 本研究所涉實驗魚生物學表型間的異同主要表現為: (1) 從稱量性狀看, 除BW無顯著差異(>0.05), NW呈M1
表1 生物學表型的統計量描述
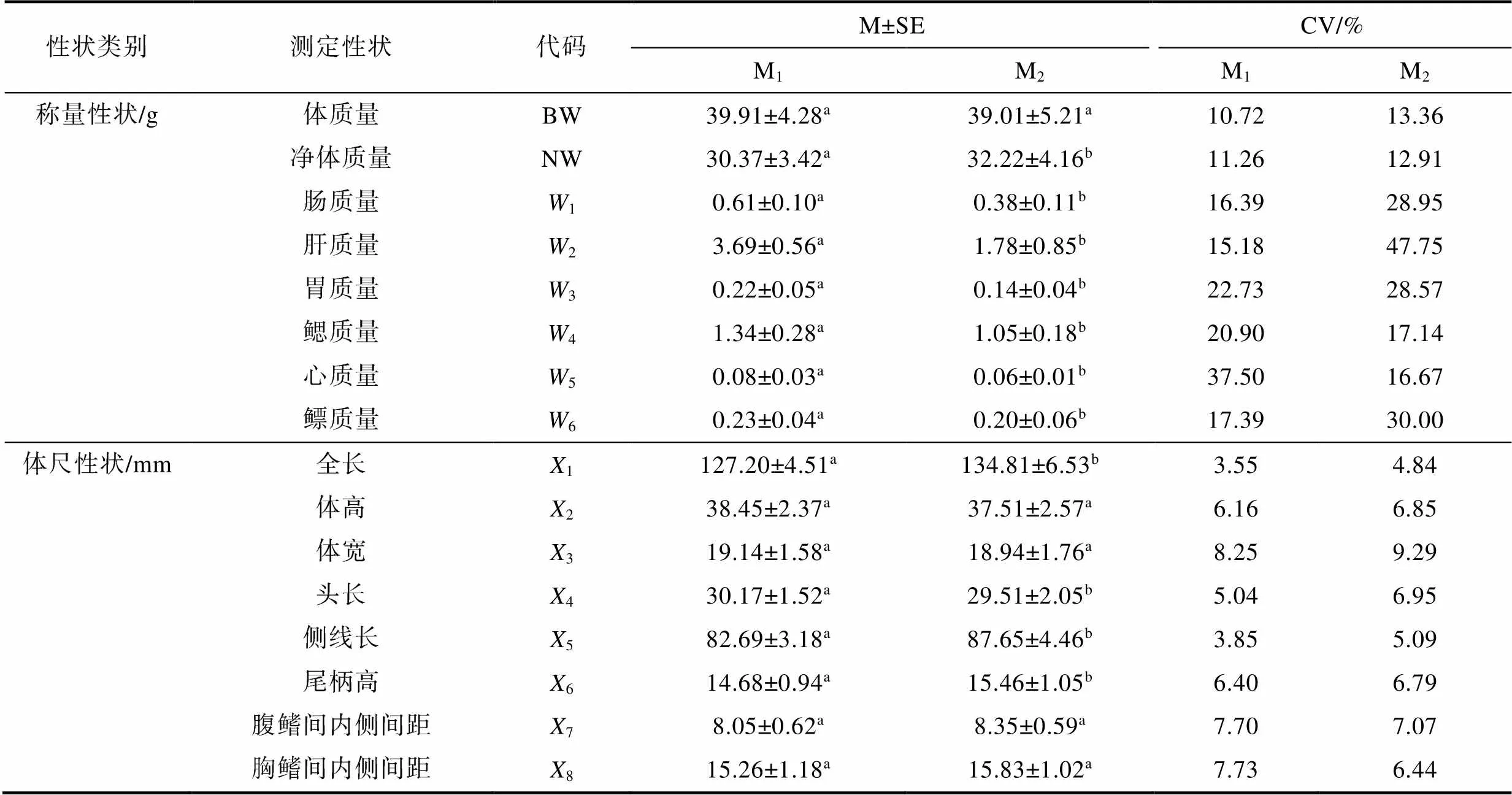
Tab.1 Description in statistics of biological phenotypes
注: 上標不同字母表示組間存在顯著差異(<0.05)。下同
2.2 相關分析
由表2、表3可見, 本研究所涉實驗群體生物學表型與BW間的相關性異同主要表現為: (1) 從稱量性狀看, M1和M2實驗群體除5與BW間的相關系數值均未達到顯著水平(>0.05)外, 其余性狀均達到極顯著水平(<0.01), 兩者與BW間的相關系數值排序分別為NW>6>4>2>1>3>5和NW>4>6>2>1>3>5; (2) 從體尺性狀看, M1和M2實驗群體各測定性狀與BW間的相關系數均達到極顯著水平(<0.01), 兩者與BW間的相關系數值排序分別呈5>1>2>4>7>6>3>8和2>3>1>6>5>8>4>7。綜上可知, 體尺性狀與BW間相關系數的組內排序受養殖模式的影響程度明顯大于稱量性狀, 其中M1和M2實驗群體稱量性狀中與BW間相關系數值最大的均為NW, 最小的均為5。
2.3 通徑分析
由表4、表5可見, 本研究所涉實驗群體間被保留性狀組合的異同主要表現為: (1) 從稱量性狀看, M1和M1被保留的性狀組合分別為NW-2-1-3和NW-2, 直接作用最大的均為NW, 間接作用最大的則分別為3和2, 其中直接作用大于間接作用的性狀均僅為NW; (2) 從體尺性狀看, M1和M2被保留的性狀組合分別為512和2315, 直接作用最大的分別為5和2, 間接作用最大的分別為2和5, 其中直接作用大于間接作用的性狀分別僅為5和2; (3) 共線性診斷顯示, M1和M2被保留的稱量性狀組合和體尺性狀組合的方差膨脹因子均遠低于經驗值(VIF<10)。綜上可知,2和5分別為體尺性狀中影響M1、M2實驗群體BW的關鍵核心變量, NW為稱量性狀中影響M1、M2實驗群體BW的公共關鍵核心變量, 其他被保留性狀則為影響BW的相對重要變量。
表2 稱量性狀與體質量間相關系數
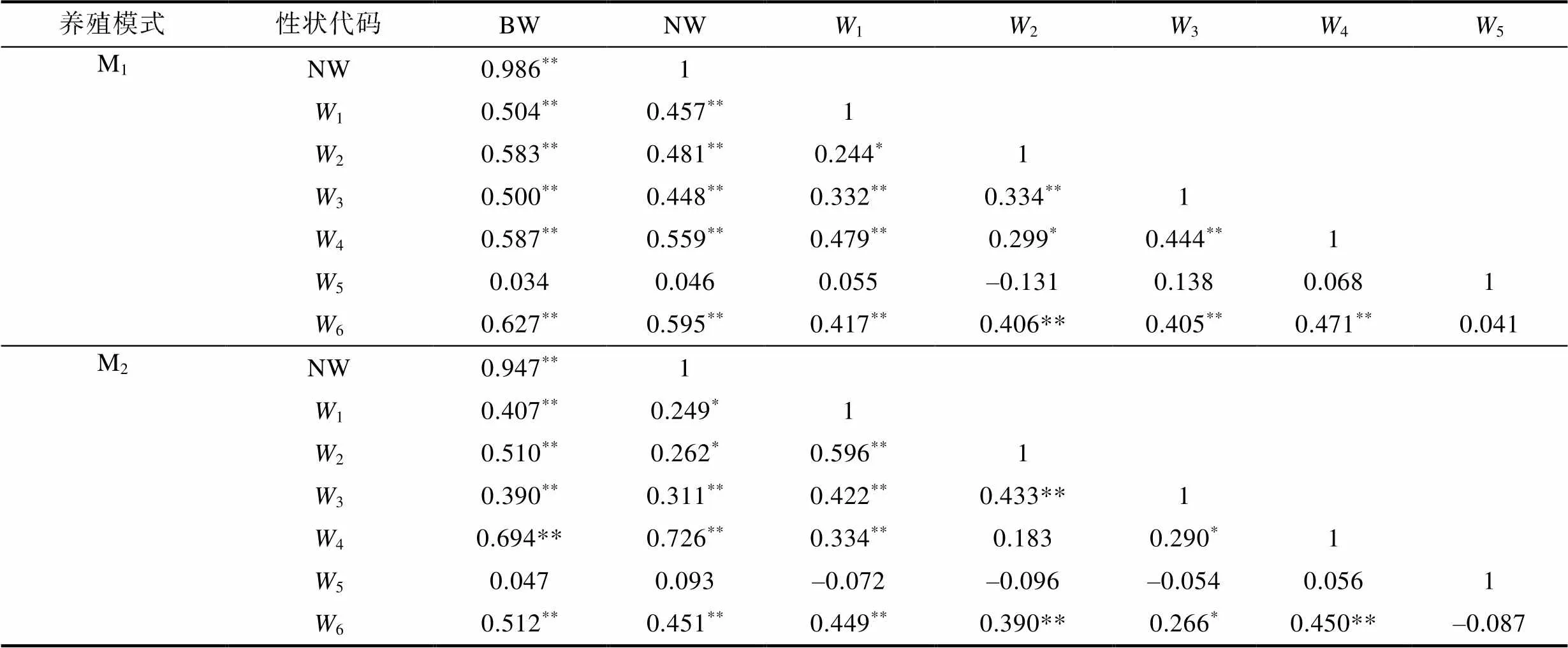
Tab.2 Correlation coefficient between quality traits and body weight
注:(0.05,68)=0.235,(0.01,68)=0.306; 上標*和**分別表示性狀間的相關系數達到顯著水平(<0.05)和極顯著水平(<0.01)。下同
表3 體尺性狀與體質量間的相關系數
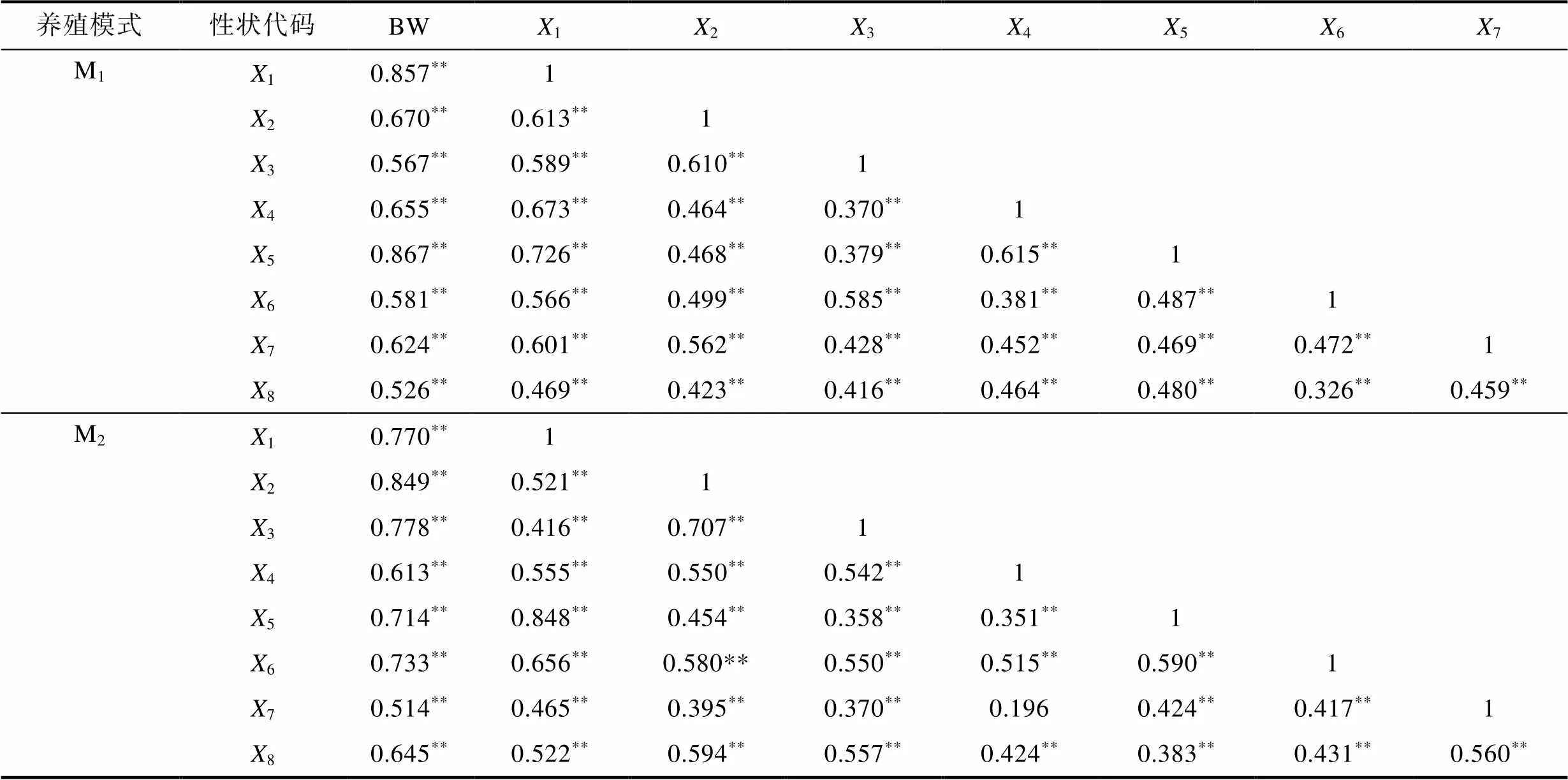
Tab.3 Correlation coefficients between body size traits and body weight
表4 稱量性狀對體質量的通徑分析

Tab.4 Path analysis of quality traits on body weight
表5 體尺性狀對體質量的通徑分析
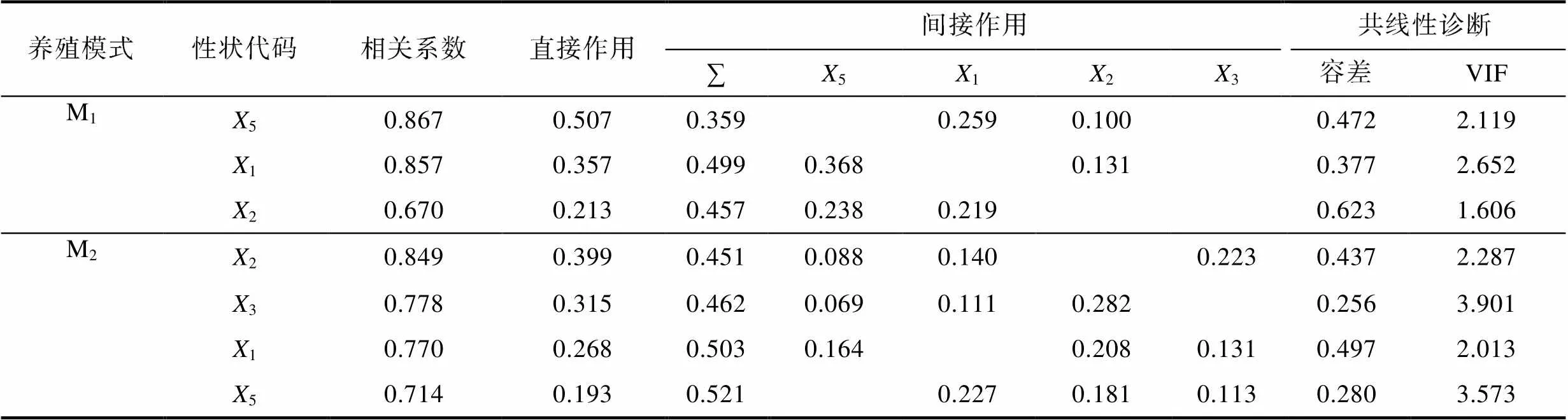
Tab.5 Path analysis of body size traits on body weight
2.4 決定程度分析與復相關分析
由表6和表7可見, M1、M2實驗群體單一稱量性狀和單一體尺性狀對BW的決定系數分別為NW>2>1>3、NW>2和5>1>2、2>1>3>5; 經計算, M1、M2被保留稱量性狀組合對BW的總決定系數分別為0.992和0.971, 體尺性狀組合對BW的總決定系數分別為0.889和0.927。復相關分析顯示(表8), M1、M2被保留性狀組合與BW的復相關系數均達到極顯著水平(<0.01), 所示復相關指數均分別與其相對應性狀組合的總決定系數完全相同。無疑, 上述結果進一步印證了表4和表5所示2和5分別為體尺性狀中影響M1、M2實驗群體BW的關鍵核心變量, NW為稱量性狀中影響M1、M2實驗群體BW的公共關鍵核心變量, 以及其他被保留性狀為影響BW相對重要變量的可靠性。
表6 稱量性狀對體質量的決定系數
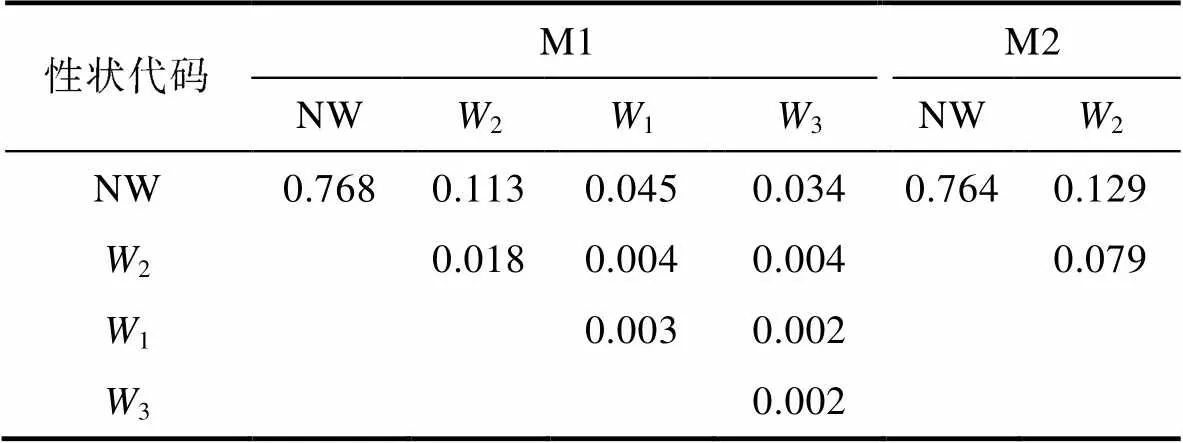
Tab.6 Determination coefficients of quality traits on body weight
表7 體尺性狀對體質量的決定系數
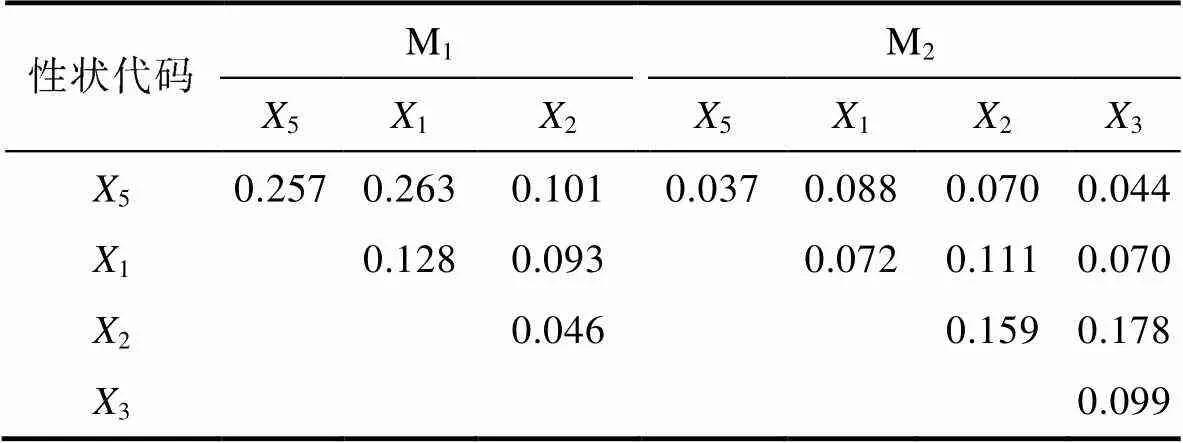
Tab.7 Determination coefficients of body size traits on body weight
表8 稱量性狀和體尺性狀對體質量的復相關系數
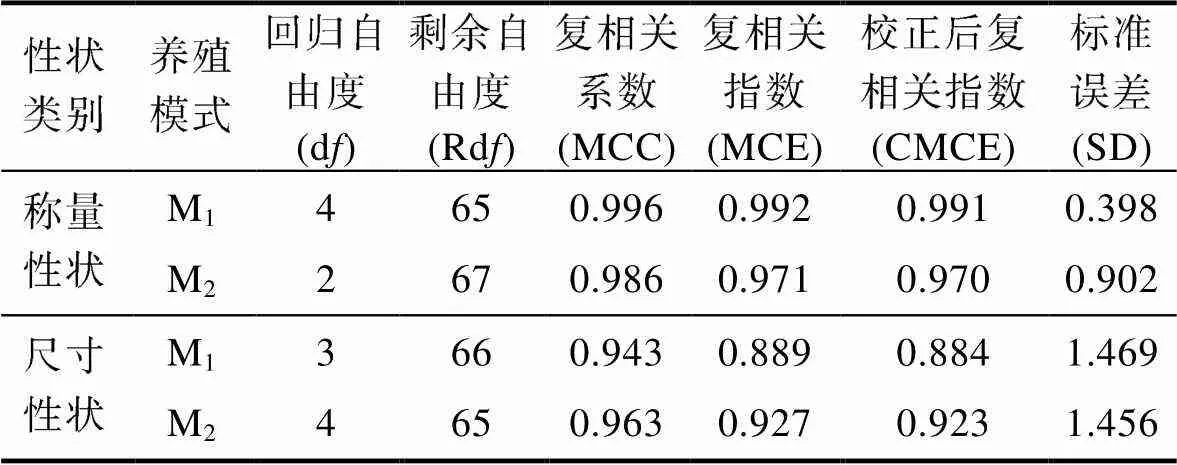
Tab.8 Multiple correlation coefficients of quality traits and body size traits on body weight
注:0.01(2, 67)=0.393;0.01 (3, 66)=0.424;0.01(4, 65)=0.427
2.5 多元線性回歸方程的建立
由表9可見, M1、M2實驗群體被保留稱量性狀組合與BW間的多元線性回歸方程分別為(W1)= 1.096NW1.0252+2.4331+3.5043+0.507和(W2)= 1.096NW+1.2872+1.044; 被保留體尺性狀組合與BW間的多元線性回歸方程分別為(L1)=0.6835+ 0.3381+0.3852-74.431和(L2)=0.8092+0.2141+ 0.9313+0.2255–57.512。方差分析結果表明, 所建立的回歸方程的回歸關系也均達到極顯著水平(<0.01) (表10)。經回歸預測, 估計值和實際測量值之間無顯著差異(>0.05), 表明所建方程能精確反映M1、M2實驗群體體尺性狀和稱量性狀與其BW之間的真實關系。
表9 生物學性狀對體質量的偏回歸分析
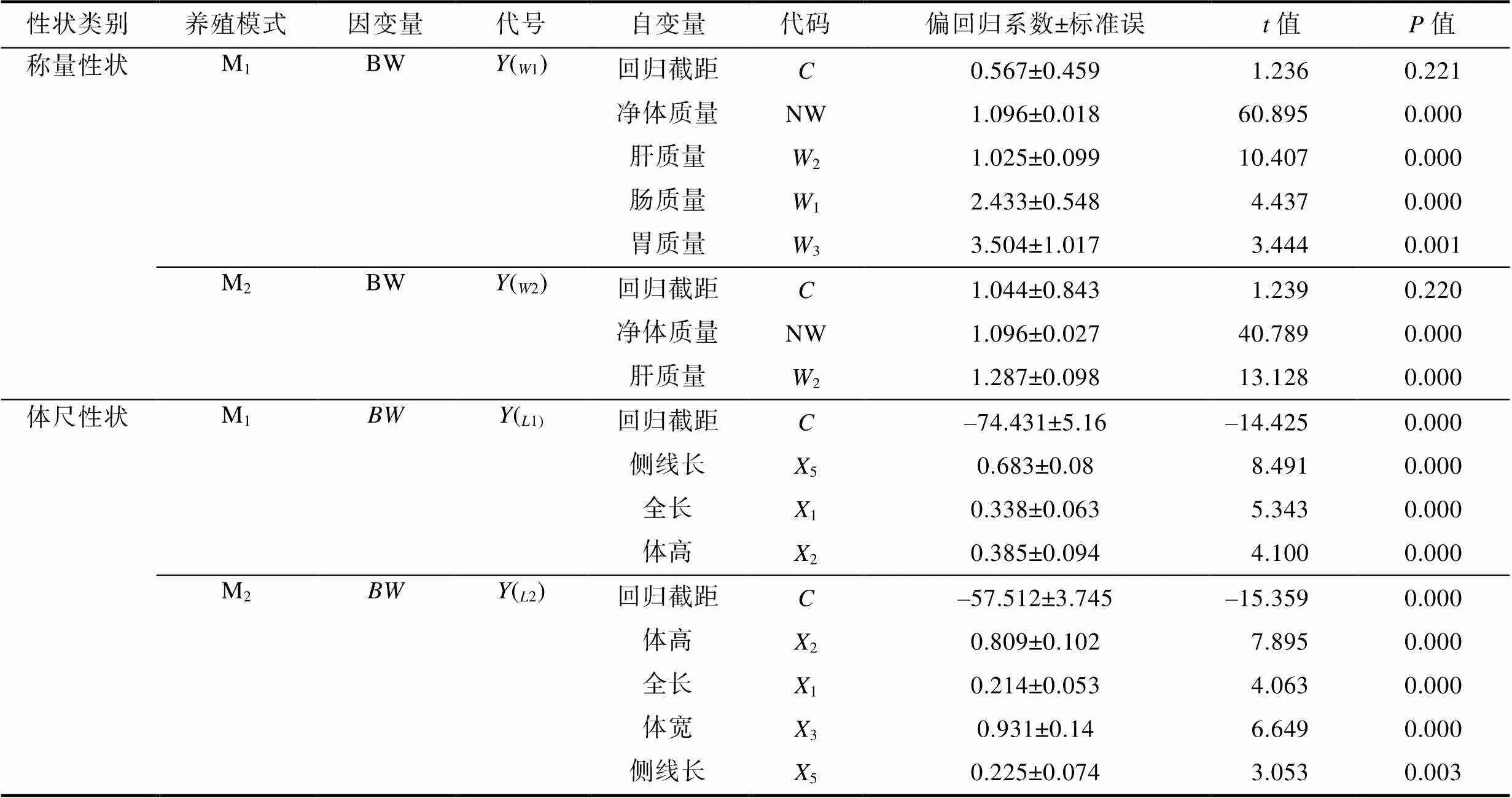
Tab.9 Partial regression analysis of biological traits on body weight
表10 生物學性狀與體質量的多元回歸方差分析
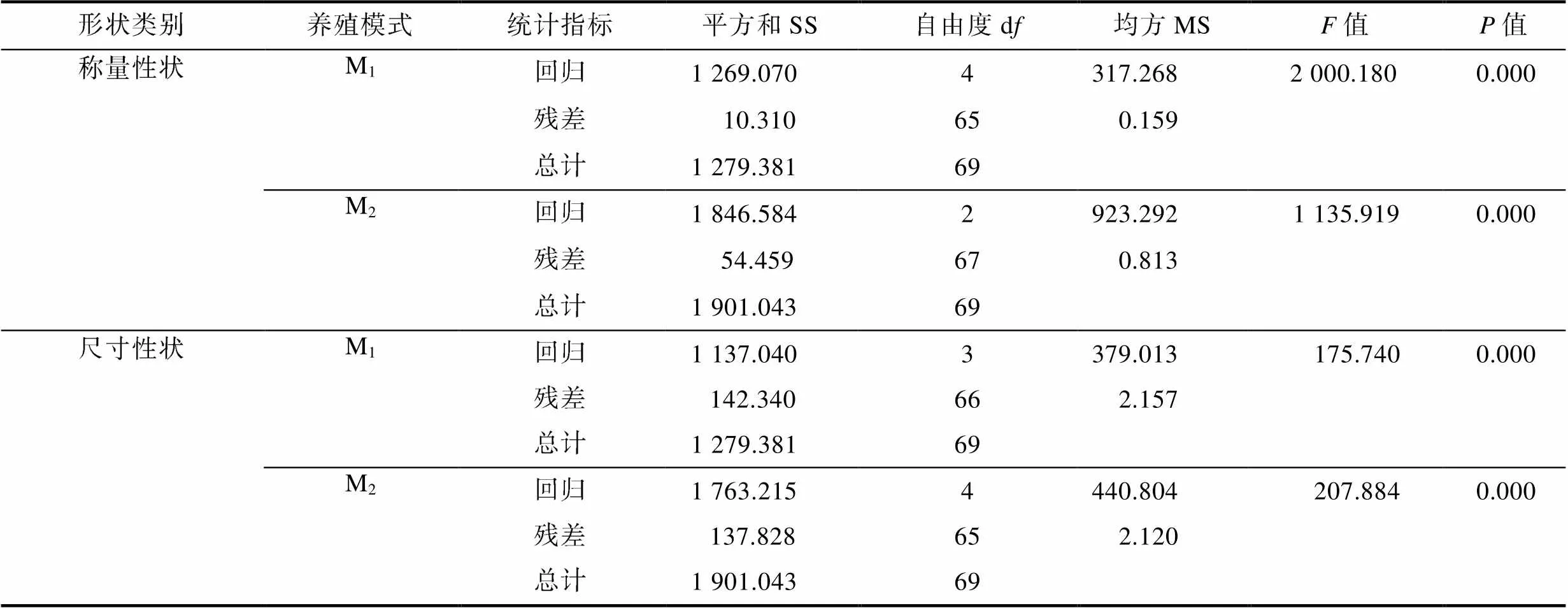
Tab.10 Multiple regression analysis of variance of biological traits and body weight
注:0.01(3,66)=4.10;0.01(4,65)=3.62;0.01(4,65)=3.62;0.01(2,67)=4.95
3 討論
3.1 M1與M2實驗群體間基于決定BW關鍵性狀組合的生存對策差異分析
由表6、表7和表8可見, NW-2-1-3、512對M1實驗群體BW的總決定系數和復相關指數均分別為0.992和0.889,2315、NW-2對M2實驗群體BW的總決定系數和復相關指數均分別為0.971和0.927, 均大于影響因變量的主要自變量組合已被找到的閾值0.85 (劉小林等, 2002), 表明上述性狀組合均為決定M1和M2實驗群體BW的關鍵性狀組合。
Wagner等(2001)認為同一物種的不同群體為長期適應不同的生活環境而形成了各自特有包括生長、發育、繁殖以及對環境因子的適應力等的生物學特性。劉建康(1999)也指出生物會基于其所處生境的變動性、可預測性和不確定性程度而在-連續譜系上作出相應的生存抉擇。據報道, 適量種植銅錢草等水生植物不僅可顯著改善水體環境并保持其相對穩定性(Pokorny, 1990; 周遺品等, 2011), 而且還可顯著提高池內水產養殖動物的存活率、生長效率并改善其養殖品質(劉鑫等, 2003; 徐增洪等, 2016)。無疑, 本研究所涉M1實驗群體較M2實驗群體具更為優越而穩定的生存環境, 且這種差異隨養殖時長的推移變得愈加顯著的情形, 勢必會導致M1與M2實驗群體在生存對策出現較明顯的分化。
由表4可見, 稱量性狀中決定M1、M2實驗群體BW的關鍵核心變量均為NW, 結合表1所示BW、NW分別呈M1≈M2(>0.05)和M1 動物的組織和臟器按其生理代謝功能可分為活性和惰性兩部分。就本研究所涉稱量性狀而言, 除NW屬惰性部分外, 其余稱量性狀均屬活性部分。無疑, 本研究所涉M1與M2實驗群體NW為稱量性狀中影響兩實驗群體BW的公共核心關鍵變量的結果(表 4), 既與“器官代謝活性假說”所述動物組織和臟器質量與體質量間呈異速生長, 活性部分和惰性部分與體質量間的比例隨個體生長分別呈下降和上升趨勢的論斷相符(Itazawa, 1983; Oikawa, 1992), 也與牟恩鏇等(2021a)所述動物體質量的增長既依賴活性臟器代謝能耗的持續增加, 更表露為惰性部位對代謝產物的不斷積累的觀點相吻。 就本研究所涉實驗群體所處環境嚴峻度而言, 因缺乏水草對養殖環境的調控, M2實驗群體的抗逆壓力明顯大于M1。故本研究所涉實驗群體的活性臟器質量均呈M1>M2(<0.05) 結果(表 1), 既反映了M2實驗群體因機體代謝能耗明顯大于M1致使活性臟器質量增速明顯受抑的客觀事實, 也映射了M2實驗群體為增強體質通過加速NW積累以響應高嚴峻度生存環境的適應對策, 更揭示了M2實驗群體為促進NW積累通過顯著抑制活性臟器質量增速以補償機體代謝能耗的增重對策。“器官代謝活性假說”認為, 活性器官的臟器系數與其代謝水平呈正相關(Itazawa, 1983; Oikawa, 1992)。馬細蘭等(2009)也指出, 胃、肝、腸作為動物重要的消化器官, 是各種營養物質消化吸收與代謝的主要場所, 其發達程度直接影響動物的生長速度。故, 決定M1、M2實驗群體BW的關鍵稱量性狀組合中均包含2的原因, 應與表 1所示兩實驗群體的消化臟器質量均呈2>1>3(<0.05) 的結果密切相關, 即肝臟是異育銀鯽最大的消化器官。至于,1、3未被選入決定M2實驗群體BW關鍵性狀組合的原因, 可能與腸、胃質量遠小于肝臟且為補償機體代謝能耗致使其增速明顯受抑, 從而導致1、3未能真實反映其消化代謝水平有關。 M1實驗群體的養殖密度為M2的25倍, 故致其種內食物競爭壓力遠高于M2。無疑, 如何切實提高種內食物競爭能力是M1實驗群體BW增長必須面對的生存抉擇。梁旭方(1998)指出, 魚類的測線感受系統具識別和定位獵物的作用。故,5取代1成為決定M1實驗群體BW的關鍵核心變量的原因, 應與M1實驗群體通過提高攝食靈敏度和成功率以提升種內食物競爭能力密切相關。 異育銀鯽與凡納濱對蝦同屬雜食性底棲種類。就BW增長而言, M2實驗群體與同池高密度養殖的凡納濱對蝦勢必在食物和棲境爭奪上均存在較激烈的種間競爭。由此, 表1所示M2實驗群體4、4和6均顯著小于M1(<0.05)的結果, 也印證M2實驗群體通過顯著降低呼吸與運動強度以更專注于持續占據池底食場生境的生態適應性。故,3被增選進入決定M2實驗群體BW關鍵性狀組合的原因, 應與M2實驗群體通過增大魚體三維幾何空間以更利于擠占食場生境有關。至于,1、2和5對M1和M2實驗群體BW的通徑系數排序上互為倒序的原因, 既反映了M1和M2實驗群體的生存對策在-連續譜系上已出現顯著偏離的客觀事實, 也揭示了M1實驗群體更注重種內食物競爭而M2實驗群體則更關注于種間食物和棲境競爭的體質量增長對策。 (1)M1實驗群體更注重臟器質量的積累和頭部的延長, 以利增強機體功能朝個體大型化方向發展的趨勢, 而M2實驗群體則更注重凈體質量的積累、全長和測線長的增長以及尾柄部的增高, 以利快速儲備能量并朝個體小型化方向發展的趨勢; 盡管養殖模式可影響稱量性狀和體尺性狀CV值的組內排序, 卻改變不了稱量性狀較體尺性狀更具可塑性, 以及兩者可塑性均分別以NW和1為最小的共性。 (2)M1和M2實驗群體除5與BW間的相關系數均未達到顯著水平(>0.05)外, 其余性狀均達到極顯著水平(<0.01); 體尺性狀與BW間相關系數的組內排序受養殖模式的影響程度明顯大于稱量性狀, 其中M1和M2實驗群體稱量性狀中與BW間相關系數值最大的均為NW, 最小的均為5。 (3)影響M1、M2實驗群體BW的關鍵稱量性狀均為NW, 關鍵體尺性狀分別為2和5; 決定M1、M2的稱量性狀組合分別為NW-2-1-3和NW-2, 體尺性狀組合分別為512和2315。可用于估算M1、M2實驗群體BW的最優多元線性回歸方程組分別為(L1)=0.6835+0.3381+0.3852–74.431、(W1)=1.096NW+1.0252+2.4331+3.5043+0.507和(L2)=0.8092+0.2141+0.9313+0.2255–57.512、(W2)=1.096NW1.2872+1.044。 (4)M2實驗群體為響應高嚴峻度生存環境更關注種間食物和棲境競爭, 在稱量性狀上采取了通過顯著抑制活性臟器質量增速以補償機體代謝能耗, 以及加速促進NW積累以增強體質并提高抗逆能力的體質量增重對策, 在體尺性狀上采取了通過加速增大魚體三維幾何空間以利驅逐池底層與其共同競爭食物和棲息空間的凡納濱對蝦的體質量增重對策, 其生存對策在-連續譜系上較更注重于種內食物競爭的M1實驗群體已明顯表露出朝選擇偏移的傾向。 馬細蘭, 張勇, 周立斌, 等, 2009. 脊椎動物雌雄生長差異的研究進展[J]. 動物學雜志, 44(2): 141-146. 王志錚, 付英杰, 楊磊, 等, 2013a. 三種養殖模式下日本鰻鱺()養成品體色和肌肉品質的差異[J]. 海洋與湖沼, 44(4): 1042-1049. 王志錚, 楊磊, 朱衛東, 2012. 三種養殖模式下日本鰻鱺養成品的形質差異[J]. 應用生態學報, 23(5): 1385-1392. 王志錚, 吳一挺, 楊磊, 等, 2011. 日本沼蝦()形態性狀對體重的影響效應[J]. 海洋與湖沼, 42(4): 612-618. 王志錚, 趙晶, 楊磊, 等, 2013b. 三種養殖模式下日本鰻鱺()養成品血清生化指標和臟器消化酶、抗氧化酶活力的差異[J]. 海洋與湖沼, 44(2): 403-408. 劉小林, 常亞青, 相建海, 等, 2002. 櫛孔扇貝殼尺寸性狀對活體重的影響效果分析[J]. 海洋與湖沼, 33(6): 673-678. 劉建康, 1999. 高級水生生物學[M]. 北京: 科學出版社: 71-72. 劉鑫, 王哲, 張一, 等, 2003. 兩種沉水植物對黃顙魚()夏花培育水體主要水質因子的影響[J]. 湖泊科學, 15(2): 184-188. 農業農村部漁業漁政管理局, 全國水產技術推廣總站, 中國水產學會, 2020. 2020中國漁業統計年鑒[M]. 北京: 中國農業出版社: 25. 孫俊龍, 沈玉幫, 傅建軍, 等, 2015. 草魚一齡前不同月齡主要形態性狀對體重影響效果的分析[J]. 上海海洋大學學報, 24(3): 341-349. 牟恩鏇, 申屠琰, 葉文婷, 等, 2021a. 池塘專養模式下中華鱉養成品組織和臟器質量對體質量的影響效果分析[J]. 浙江海洋大學學報(自然科學版), 40(6): 498-504. 牟恩鏇, 申屠琰, 熊玉鳳, 等, 2021b. 池塘專養模式下中華鱉養成品形態性狀對體質量和凈體質量的影響效應[J]. 浙江海洋大學學報(自然科學版), 40(1): 22-28, 79. 李璽洋, 白俊杰, 樊佳佳, 等, 2012. 二齡草魚形態性狀對體質量影響效果的分析[J]. 上海海洋大學學報, 21(4): 535-541. 楊明秋, 劉金葉, 王永波, 等, 2014. 不同養殖模式對駝背鱸生長的影響[J]. 熱帶生物學報, 5(2): 132-135. 周洲, 李建光, 王艷艷, 等, 2017. 清水江鯉外部形態性狀對體質量的影響[J]. 河南農業科學, 46(12): 134-138. 周康奇, 潘賢輝, 林勇, 等, 2020. 廣西禾花鯉形態性狀與體質量的關系分析[J]. 河南農業科學, 49(9): 159-165. 周遺品, 劉雯, 雷澤湘, 等, 2011. 銅錢草對城市生活污水的凈化效果[J]. 仲愷農業工程學院學報, 24(2): 9-12. 趙旺, 江淼, 陳明強, 等, 2018. 離岸養殖與陸基養殖尖吻鱸形態性狀與體質量的相關性研究[J]. 海洋學報, 40(8): 53-62. 趙柳蘭, 龍亞男, 羅杰, 等, 2021. 池塘和稻田兩種養殖模式下建鯉腸道菌群、免疫酶活性及肌肉氨基酸比較分析[J]. 中國水產科學, 28(1): 48-56. 袁向陽, 王志錚, 楊程, 等, 2017a. 兩種養殖模式下泥鰍養成品體色、質構及其臟器消化酶、抗氧化酶和ATP酶活力的差異[J]. 漁業科學進展, 38(2): 121-127. 袁向陽, 李鴻鵬, 楊程, 等, 2016. 泥鰍養成品形態性狀與體質量和凈體質量的相互關系[J]. 南方農業學報, 47(7): 1234-1240. 袁向陽, 富裕, 楊程, 等, 2017b. 稻田養殖泥鰍形態性狀與體質量、凈體質量關聯分析的研究[J]. 海洋湖沼通報, 39(1): 88-95. 原居林, 劉梅, 倪蒙, 等, 2018. 不同養殖模式對大口黑鱸生長性能、形體指標和肌肉營養成分影響研究[J]. 江西農業大學學報, 40(6): 1276-1285. 徐增洪, 劉國鋒, 水燕, 等, 2016. 人工栽培水草對池塘養殖生態環境和蝦蟹生長的影響[J]. 江蘇農業科學, 44(6): 328-331. 梁旭方, 1998. 魚類側線機械感覺獵物識別和定位的研究進展[J]. 動物學雜志, 33(2): 50-53. ITAZAWA Y, OIKAWA S, 1983. Metabolic rates in excised tissues of carp [J]. Experientia, 39(2): 160-161. OIKAWA S, TAKEMORI M, ITAZAWA Y, 1992. Relative growth of organs and parts of a marine teleost, the porgy,, with special reference to metabolism-size relationships [J]. Japanese Journal of Ichthyology, 39(3): 243-249. POKORNY J, KVET J, ONDOK J P, 1990. Functioning of the plant component in densely stoked fishponds [J]. Bulletin of Ecology, 21(3): 44-48. WAGNER E J, ARNDT R E, BROUGH M, 2001. Comparative tolerance of four stocks of cutthroat trout to extremes in temperature, salinity, and hypoxia [J]. Western North American Naturalist, 61(4): 434-444. DIFFERENTIAL ANALYSIS ON THE EFFECT OF BIOLOGICAL PHENOTYPES ON BODY WEIGHT OFIN TWO POND AQUACULTURE MODES CHEN Yu1, RONG Hua-Nan1, 2, LIU Zhe-Yu1, CHEN Han-Chun2, WANG Zhi-Zheng1 (1. College of Fisheries, Zhejiang Ocean University, Zhoushan 316022, China; 2. Cixi Fisheries Technology Promotion Center, Cixi 315300, China) Seventy allogynogenetic crucian carps were cultured for 7 months in a pond ecological dominant culture mode (M1) and a pond ecological polyculture mode (M2), and then randomly selected for the research. The total length (1), body height (2), body width (3), head length (4), lateral line length (5), tail handle height (6), ventral fin spacing (7), and pectoral fin spacing (8) were used as dimensional traits, while the net body mass (NW), intestinal mass (1), liver mass (2), stomach mass (3), gill mass (4), heart mass (5) and swim bladder mass (6) were used as weighing characters. And the multivariate analysis method was used to study the effects of weighing and size traits on body mass (BW) of gibel carp in the two breeding modes. Results show the following. (1) The CV (coefficient of variation ) values greater than 10% were from weighing traits, and those less than 10% from body size traits. There was no significant difference traits (>0.05) except for NW,1,5and6whose M1 culture mode;; body size traits; weighing traits; body mass Q953; S965 10.11693/hyhz20220100022 *國家星火計劃項目, 2013GA701022號; 寧波市重大科技攻關項目, 2012C10032號; 慈溪市農業科技重點項目, CN201901號。陳 雨, 碩士研究生, E-mail: chen_yuyuer@163.com; 同等貢獻第一作者: 戎華南, E-mail: 13706742262@139.com 王志錚, 教授, E-mail: wzz_1225@163.com 2022-01-26, 2022-03-143.2 M1與M2實驗群體間基于稱量性狀的BW增長對策差異分析
3.3 M1和M2實驗群體基于體尺性狀的BW增長對策差異
4 結論
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
中國生殖健康(2019年2期)2019-08-23 08:12:08
產品可靠性報告(2017年7期)2017-09-05 09:49:12
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
汽車觀察(2016年3期)2016-02-28 13:16:26
