大口黑鱸(Micropterus salmoides)致病性維氏氣單胞菌的分離鑒定及其特性分析*
2022-09-21 02:36:06郝貴杰黃愛霞王雨辰沈小明朱俊杰
海洋與湖沼 2022年5期
雷 寧 郝貴杰 黃愛霞 王雨辰 林 鋒① 沈小明 朱俊杰①
大口黑鱸()致病性維氏氣單胞菌的分離鑒定及其特性分析*
雷 寧1郝貴杰2黃愛霞2王雨辰2林 鋒2①沈小明3朱俊杰1①
(1. 湖州師范學院生命科學學院 浙江湖州 313000; 2. 農業農村部淡水漁業健康養殖重點實驗室 浙江省魚類健康與營養重點實驗室 浙江省淡水水產研究所 浙江湖州 313001; 3. 德清昊源水產種業有限公司 浙江湖州 313200)
大口黑鱸()是我國重要的淡水養殖經濟魚類品種, 目前包括細菌性疾病在內的諸多因素影響了其養殖業的健康發展。維氏氣單胞菌是引起淡水魚敗血癥和潰瘍綜合征的重要病原菌。從患病大口黑鱸肝臟組織中分離到1株病原菌, 通過形態學觀察、生理生化特性分析、16S rRNA序列和特定毒力基因分析對其進行鑒定, 并通過人工回歸感染試驗和藥敏試驗分析其致病性和耐藥性。結果表明該菌為致病性維氏氣單胞菌(), 含有、、、、和等毒力基因; 藥敏試驗顯示其對頭孢曲松、氯霉素、氟苯尼考、強力霉素、四環素、多粘菌素、復方新諾明、左氟沙星等8種抗生素高度敏感, 對新霉素、恩諾沙星、慶大霉素、鏈霉素、阿齊霉素、環丙沙星等6種抗生素中度敏感, 對其他8種抗生素具有一定耐藥性; 人工回歸感染實驗表明該菌具有較高的致病性, 對大口黑鱸的半致死濃度為1.4×106CFU/mL。維氏氣單胞菌是水產養殖中的條件性致病菌, 研究結果揭示了該菌株的部分生物學特性, 有利于豐富維氏氣單胞菌屬的基礎數據, 同時也為鱸魚養殖過程中的病害防控提供了一定的參考依據。
大口黑鱸; 維氏氣單胞菌; 藥敏試驗; 毒力基因
大口黑鱸()俗稱加州鱸, 隸屬于鱸形目(Perciformes)、鱸亞目(Porcoidei)、太陽魚科(Cehtrachidae)、黑鱸屬(), 是廣溫性淡水魚類, 肉質鮮嫩、營養豐富、養殖周期短、生長速度快, 是國內重要經濟性淡水養殖品種之一。據統計, 中國大口黑鱸產量占據全球總產量99.8% (Hussein, 2020), 至2019年, 資料顯示我國大口黑鱸年產量近50×104t。全國已有20多個省市養殖生產淡水鱸, 養殖面積超過1.33×104hm2(張紅燕等, 2019)。國內大口黑鱸最大產區是華南的廣東地區, 約占全國60%, 華東的浙江、江蘇地區產量約占全國30% (陳莉莉, 2018)。隨著養殖規模的不斷擴大, 區域生產不平衡、價格波動大、種質資源退化及病害等因素, 嚴重影響了大口黑鱸養殖業的健康發展(Hussein, 2020)。細菌、病毒和寄生蟲等是誘發大口黑鱸病害的重要原因, 其中細菌性疾病以柱狀黃桿菌病、氣單胞菌病、諾卡氏菌病為主, 病毒性疾病以病毒性潰瘍病、脾腎壞死病和彈狀病毒病為主, 寄生蟲病以車輪蟲、斜管蟲、指環蟲、累枝蟲、錨頭鳋等病原為主(國家特色淡水魚產業技術體系, 2021)。而細菌性疾病是魚類病害中占比極高, 具有傳染性較強、覆蓋區域廣、流行時間長的特點, 由于新毒株增加快, 導致控制難度大, 其造成的經濟損失約占據全部疾病所造成總體經濟損失的半數以上(王玉堂等, 2013)。在淡水魚類細菌性疾病中, 屬細菌性敗血病造成的損失最為嚴重, 其病原體涵括了溫和氣單胞菌、嗜水氣單胞菌以及維氏氣單胞菌(藺凌云等, 2018)。
維氏氣單胞菌()隸屬弧菌科、氣單胞菌屬, 屬于革蘭氏陰性菌, 可感染無脊椎動物、水生脊椎動物以及包括人類在內的哺乳動物(Lazado, 2018)。維氏氣單胞菌對大口黑鱸、草魚() (高彩霞等, 2018)、框鏡鯉() (胡天野等, 2012)、黃顙魚() (朱成科等, 2017)、懷頭鯰() (陸夢瑩等, 2017)、藍曼龍魚() (陳亨利等, 2016)、齊口裂腹魚() (劉港彪等, 2012)、泥鰍() (徐先棟等, 2018)、胭脂魚() (黃文明等, 2013)、鳙魚() (朱若林等, 2017)、中華鱘() (田甜等, 2017)、鯽魚() (王利等, 2013)和羅非魚(Tilapia)(黎炯等, 2011)等魚類都具有一定致病性。維氏氣單胞菌廣泛存在于自然界中, 且有著較強的適應性, 是引起淡水魚敗血癥和潰瘍綜合征的重要病原菌(Li, 2020)。水產品作為蛋白質等營養物質的重要來源之一。水產品若被維氏氣單胞菌污染, 人們食用后可能會誘發敗血癥及胃腸炎等疾病, 免疫力低下者甚至會發生死亡(鄧龍君, 2021)。因此, 本研究對發病大口黑鱸病原菌進行分離鑒定, 通過形態學觀察、生理生化特性分析、16S rDNA 序列鑒定、藥敏試驗及毒力基因檢測對分離菌株的生物學特性進行分析, 以期為大口黑鱸維氏氣單胞菌病的診斷和有效防治提供參考。
1 材料與方法
1.1 試驗材料
患病大口黑鱸[(10±1.5) cm]采自浙江省湖州市某養殖場。健康大口黑鱸[(7.00±0.65) g, (7.9±0.42) cm]購自湖州湖旺水產種業有限公司, 在50 L養殖桶內暫養7 d后用于感染試驗。
胰蛋白胨大豆肉湯(TSB)和胰蛋白胨大豆瓊脂(TSA)培養基均購于青島海博生物科技有限公司, 細菌生化微量鑒定管和藥敏試紙購自杭州濱和微生物試劑有限公司, 革蘭氏染色試劑盒購自索萊寶公司, 細菌基因組DNA提取試劑盒購自天根生化科技(北京)有限公司, OneTaq Quick-Load?2×預混液購自NEB公司, 所用引物均由上海生工生物工程有限公司合成。
1.2 細菌分離
患病大口黑鱸體表經消毒后, 無菌環境下解剖檢查魚體, 用接種針從內臟組織各部位取樣, 分別在TSA培養基上劃線培養, 28 °C恒溫培養12 h; 觀察菌落形態, 根據菌落特征的不同, 挑取優勢菌株在TSB培養基中培養, 分離純化得到的菌株命名為LB2101。
1.3 細菌形態學觀察
根據革蘭氏染色試劑盒操作說明對該分離菌進行染色。先挑取單菌落涂片固定, 滴加草酸銨結晶紫染色液初染1 min, 水洗; 滴加碘液媒染1 min, 水洗; 用95%酒精脫色約45 s, 水洗, 吸干水分; 最后滴加蕃紅復染1 min, 水洗, 自然晾干; 在100倍油鏡下觀察細菌形態以及染色特征, 并拍照保存。
將分離菌株接種于TSB培養基中, 振蕩培養過夜, 低速離心收集菌體, PBS洗滌后滅菌水重懸備用。將處理后的菌液與適量2%鋨酸小心混勻, 后把銅網覆蓋于混合溶液上, 靜置5 min左右; 滴加適量負染液磷鎢酸溶液于封口膜上, 處理好的銅網轉移至負染液磷鎢酸上靜置2 min左右, 吸去銅網上多余染液, 自然晾干, 透射電子顯微鏡下觀察并拍照保存。
1.4 生理生化特性分析
參照《常見細菌系統鑒定手冊》(東秀珠等, 2001)中細菌鑒定方法, 并采用細菌微量生化鑒定管對待測菌株各項生化指標進行測定, 每項檢測指標均重復3次并設置對照。將分離菌株LB2101接種 TSA培養基, 28 °C培養至長出單菌落。挑取新鮮單菌落分別接種于22種細菌微量生化鑒定管中, 28 °C培養24 h或48 h, 后根據測定結果分析該菌株的生理生化特性。
1.5 分子生物學鑒定
根據細菌基因組DNA提取試劑盒方法提取該分離菌株的DNA。采用細菌通用引物進行16S rRNA基因序列的擴增, 正向引物為27F: 5′-AGAGTTTG ATCCTGGCTCAG-3′, 反向引物為1492R: 5′-TACG GTTACCTTGTTACGAC-3′, 擴增片段大小約1 470 bp。擴增反應體系(50 μL): DNA模板2 μL, 正向和反向引物各1 μL, OneTaq Quick-Load?2×預混液25 μL, RNase-Free Water補至50 μL。PCR擴增條件: 95 °C預變性4 min; 95 °C變性30 s, 55 °C退火30 s, 72 °C延伸1 min, 35個循環后72 °C延伸10 min。PCR產物進行1% 凝膠電泳檢測。將擴增產物送至生工生物工程(上海)有限公司進行測序, 根據核苷酸測序結果, 利用NCBI中Blast檢索系統進行同源序列比對, 利用MEGA 6.0軟件構建基于細菌 16S rRNA基因的系統進化樹, 采用鄰接法(Neighbor joining method)建樹方法進行菌株LB2101的系統發育樹分析。
1.6 藥敏試驗
利用K-B紙片瓊脂擴散法進行藥敏實驗, 28 °C培養24 h后測定抑菌圈直徑, 測定3次取平均值, 根據杭州濱和微生物試劑有限公司提供的標準進行細菌敏感性判定。檢測的抗生素包含頭孢曲松、氯霉素、氟苯尼考、新霉素、恩諾沙星、強力霉素、阿米卡星、復方新諾明、四環素、諾氟沙星、乙酰螺旋霉素、羧芐西林、青霉素、頭孢噻吩、多粘菌素、痢特靈、慶大霉素、鏈霉素、利福平、阿齊霉素、環丙沙星、左氟沙星等22種。
1.7 毒力基因檢測
根據已發表文獻合成氣溶素基因、細胞毒性腸毒素基因、脂肪酶基因、膽固醇酰基轉移酶基因、脫氧核糖核酸酶基因、鞭毛蛋白結構基因及外膜蛋白等毒力相關基因的特異性引物(見表1), 以病原菌DNA為模板進行PCR檢測。擴增反應體系(25 μL): DNA模板1 μL, 正向和反向引物各0.5 μL, OneTaq Quick-Load?2×預混液12.5 μL, RNase-Free Water補至25 μL。PCR擴增條件: 95 °C預變性5 min; 95 °C變性30 s, 退火溫度30 s, 72 °C延伸1 min, 35個循環后72 °C延伸10 min。PCR產物經瓊脂糖凝膠電泳進行分析。
1.8 人工回歸感染試驗
將分離菌株接種于TSB培養基中, 28 °C振蕩培養12h, 低速離心收集菌體, 用生理鹽水將菌液濃度分別調節至1.4×105、1.4×106、1.4×107、1.4×108和1.4×109CFU/mL。健康大口黑鱸隨機分為6組, 每組30尾, 試驗組分別腹腔注射不同濃度菌液, 注射劑量為100 μL/尾, 對照組注射等量生理鹽水。試驗期間水溫控制在28~30 °C。感染后每隔24 h觀察大口黑鱸的臨床表現并統計死亡情況, 連續監測7d。從發病癥狀明顯的大口黑鱸肝臟等組織中分離病原菌, 進行分離培養及鑒定, 以確定是否是該分離菌株導致大口黑鱸死亡。
表1 毒力基因PCR擴增引物

Tab.1 Primers for virulence gene testing
2 結果與分析
2.1 細菌形態學觀察
根據觀察結果, LB2101菌落具有菌落呈圓形、表面光滑不透明、微隆起、呈現淡黃色。根據細菌革蘭氏染色結果(圖1a), 顯示該菌染色后呈紅色, 呈桿狀, 初步判定為革蘭氏陰性菌。經透射電鏡觀察發現該菌體無側鞭毛, 極端著生單鞭毛(圖1b)。
2.2 生理生化特性分析
分離菌株LB2101對精氨酸脫羧酶、葡萄糖產氣、葡萄糖、蛋白胨水、苯丙氨酸、明膠、甘露醇和蔗糖等呈陽性; 而對賴氨酸脫羧酶、鳥氨酸脫羧酶、氨基酸對照、葡磷胨水、葡萄糖酸鹽、側金盞花醇、山梨醇、棉子糖、木糖、水楊素、尿素、枸櫞酸鹽、硫化氫和半固體瓊脂等呈陰性反應(見表2)。結合細菌生化鑒定結果并根據文獻(東秀珠等, 2001)的鑒定方法, 發現分離菌株LB2101與維氏氣單胞菌的生理生化特性基本一致。
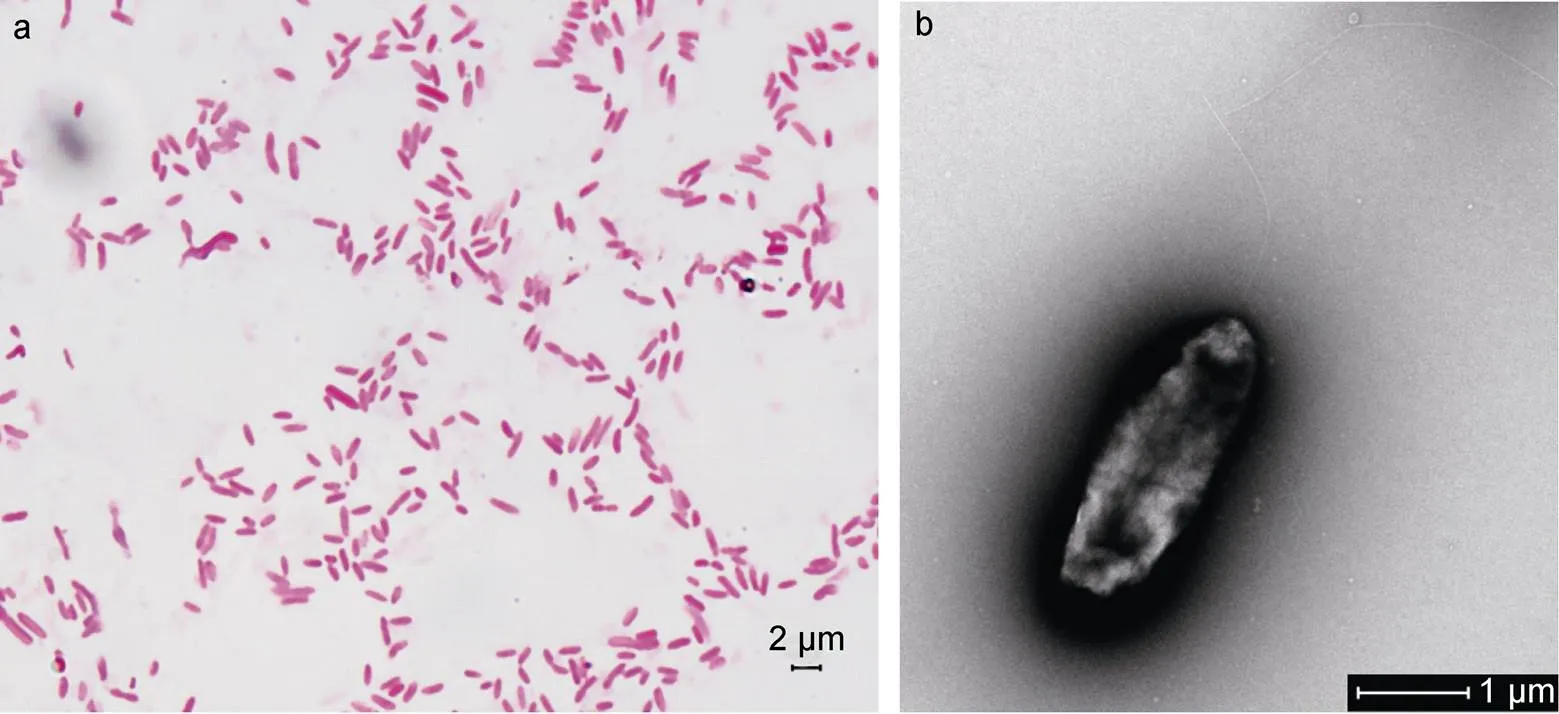
圖1 分離菌株的革蘭氏染色(a)形態及電鏡照片(b)
表2 分離菌LB2101的理化特性

Tab.2 The physiological and biochemical characteristics of LB2101
注: +表示陽性, –表示陰性
2.3 16S rRNA序列測定和系統發育學分析
利用引物27F和1492R進行PCR擴增, 其1%瓊脂糖凝膠電泳結果(如圖2)所示, 產物大小約1 470 bp。將該分離菌的核酸序列在NCBI庫上進行比對, 比對結果表明該菌株與維氏氣單胞菌HC050630A-1 (EF669480.1)同源性較高, 兩者同源性達99.24%; 該菌株16S rRNA序列構建系統發育樹的結果(見圖3), 顯示LB2101與維氏氣單胞菌HC050630A-1聚為一個分支。綜合菌株LB2101的表型、生理生化特征及16S rRNA測序及系統發育樹分析結果, 可以判定菌株LB2101屬于維氏氣單胞菌()。

圖2 菌株LB2101 16S rRNA PCR電泳結果
注: M: DL2000 DNA Marker; 1: LB2101

圖3 菌株LB2101 16S rRNA基因序列系統發育樹
2.4 藥敏試驗
利用紙片擴散法進行藥敏實驗。結果顯示, 該分離菌株LB2101對頭孢曲松、氯霉素、氟苯尼考、強力霉素、四環素、多粘菌素、復方新諾明、左氟沙星等8種抗生素高度敏感, 對新霉素、恩諾沙星、慶大霉素、鏈霉素、阿齊霉素、環丙沙星等6種抗生素中度敏感; 對阿米卡星、諾氟沙星、乙酰螺旋霉素、羧芐西林、青霉素、頭孢噻吩、痢特靈、利福平等8種抗生素具有耐藥性。
表3 菌株LB2101的藥物敏感性試驗結果

Tab.3 The antimicrobial susceptibility test of LB2101
注: S表示高度敏感, I表示中度敏感, R表示耐藥
2.5 毒力基因檢測
根據瓊脂糖凝膠電泳檢測結果(圖4), 檢測表明菌株LB2101含有氣溶素基因、細胞毒性腸毒素基因、脂肪酶基因、膽固醇酰基轉移酶基因、鞭毛蛋白結構基因和外膜蛋白基因, 但不具有脫氧核糖核酸酶基因。
2.6 人工回歸感染試驗
根據人工回歸感染試驗結果, 試驗期間對照組大口黑鱸未見異常, 試驗組出現不同程度死亡情況(詳見表4)。大口黑鱸在感染后第2天開始出現死亡, 1.4×108CFU/mL濃度組48h內死亡率達到100%; 1.4×106CFU/mL濃度組第3天開始出現死亡, 死亡率為50%; 而低濃度組1.4×104CFU/mL在第5天出現發病癥狀, 但該濃度組卻并未發生大量死亡。感染后發病的大口黑鱸出現游泳不正常、鱗片脫落、肛門紅腫、腹部膨大等臨床表現(圖5), 解剖發現腹腔有積液、肝臟腫大出血, 與自然發病大口黑鱸的癥狀相似, 且從發病魚體中可再次分離到該菌, 這充分表明LB2101分離株對大口黑鱸具有較強的致病性。

圖4 分離菌株毒力基因檢測
注: M: DL2000 DNA Marker; 1:; 2:; 3:; 4:; 5:; 6:; 7:
表4 分離菌LB2101的回歸感染試驗
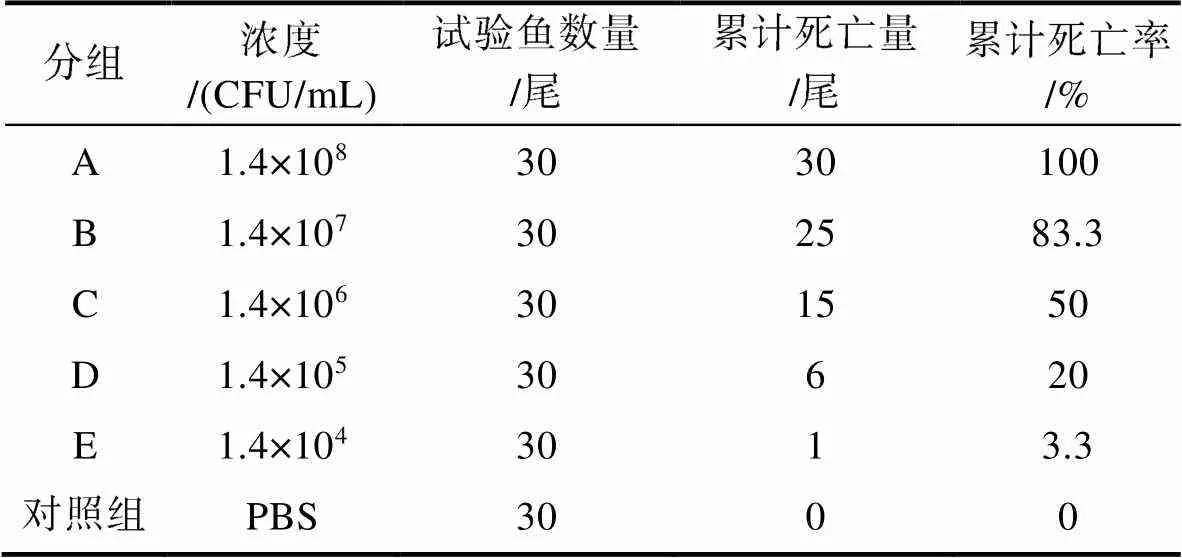
Tab.4 Pathogenic test of the isolated bacteria LB2101
3 討論
3.1 維氏氣單胞菌的分離鑒定
細菌性病原菌的鑒定方法有多種, 包括了血清分型、形態學觀察、生理生化鑒定、遺傳學分析等方法(宋路萍等, 2020)。其中, 16S rRNA基因檢測被許多學者認為是細菌鑒定的有效方法(Kita-Tsukamoto, 1993)。16S rDNA序列同時包含了高度保守序列和高度可變序列, 可用于研究各類生物間的遺傳進化關系(宋路萍等, 2020)。孟令緣等(2021)利用16S rDNA序列測定方法對18株金黃色葡萄球菌進行鑒定, 結果表明可將其中15株菌鑒定到種水平。16S rDNA序列分析法可將菌株鑒定到種水平, 表明其鑒定結果準確可靠。本研究結合利用形態觀察、生理生化測定及16S rRNA分子鑒定法, 對從大口黑鱸肝臟中分離出的菌株LB2101進行鑒定。本研究通過革蘭氏染色發現該分離菌LB2101是革蘭氏陰性菌, 呈桿狀, 具極端單鞭毛; 生理生化結果可初步判定LB2101為維氏氣單胞菌; 利用16S rRNA分子鑒定法進一步證實菌株LB2101是維氏氣單胞菌()。

圖5 患病大口黑鱸的癥狀
注: a、b和c為自然患病, d、e和f為人工感染。a: 鱗片脫落; b: 腹部膨大, 肛門紅腫; c: 腹腔積液; d、e: 鱗片脫落, 肛門紅腫; f: 肝臟有出血點
3.2 維氏氣單胞菌的毒力與致病性分析
維氏氣單胞菌可以感染大口黑鱸(楊超等, 2021)、草魚(高彩霞等, 2018)、鳙魚(朱若林等, 2017)、藍曼龍魚(陳亨利等, 2016)、鯽魚(王利等, 2013)、羅非魚(黎炯等, 2011)和胭脂魚(黃文明等, 2013)等多種水生魚類, 感染后臨床癥狀存在一定差異, 但大致都出現鱗片脫落、肛門紅腫及腹部有積液等臨床表現。該菌屬的致病性由多重因素決定, 涉及水生態環境以及毒力因子的分泌表達等。已知維氏氣單胞菌具有多種毒力因子, 如氣溶素、腸毒素、外膜蛋白、鞭毛、溶血素、細胞外蛋白酶、脂多糖和S層等(宋明芳等, 2018)。氣溶素通過與糖基磷脂酰肌醇(GPI)錨定蛋白相結合, 與宿主細胞發生相互作用在質膜上形成一個通道, 從而引發小離子的滲透、激活G蛋白和某些細胞類型的凋亡(Abrami, 2003)。而細胞毒性腸毒素, 作為維氏氣單胞菌重要毒力因子之一, 它具有使腸道絨毛和小腸隱窩退化的作用, 可刺激腸道促進其分泌作用從而導致小腸上皮細胞廣泛性損失(熊廣澤等, 2021)。外膜蛋白則是革蘭氏陰性菌的重要結構特征, 它能夠促進細菌黏附作用和增強致病性(宋明芳等, 2018)。有研究表明基因缺失可導致維氏氣單胞菌的毒力和粘附力顯著降低, 而基因編碼蛋白屬于外膜蛋白(Yang, 2019), 這間接說明外膜蛋白與維氏氣單胞菌的毒力或致病性密切相關。此外, 鞭毛在氣單胞菌侵染魚類細胞系的過程中也起著關鍵作用, 它的存在可作為菌株具有致病性的特征之一, 可增強病原細菌與真核細胞表面間的相互作用(Merino, 1997; Gavín, 2003)。
其中, 氣溶素()是維氏氣單胞菌感染魚類過程中最重要、最豐富的毒力因子之一(Foysal, 2019)。Li等(2020)從多種魚類體中分離出87株維氏氣單胞菌, 毒力基因檢測結果表明氣溶素()檢出率為88.51%。氣溶素的存在可作為判定菌株是否具有毒力的標志(趙武義, 2018)。本研究檢測了維氏氣單胞菌7種毒力相關基因, 結果表明該分離菌株LB2101同時含有氣溶素()、細胞毒性腸毒素()、脂肪酶()、膽固醇酰基轉移酶()、鞭毛()和外膜蛋白()等多種致病性毒力因子, 但不具有脫氧核糖核酸酶()毒力基因。綜合人工回歸感染試驗結果, 病魚出現鱗片脫落、肛門紅腫及腹腔積液等明顯癥狀, 充分表明本分離菌株具有顯著的致病性。
3.3 維氏氣單胞菌的藥敏性分析
不同養殖環境中分離出的維氏氣單胞菌, 其對抗生素的耐藥性會存在一定差異。如鄧龍君和楊超等分別從四川和佛山某養殖場分離得到維氏氣單胞菌, 藥敏試驗結果表明兩菌株對氟苯尼考和大觀霉素都敏感, 均對復方新諾明和阿莫西林產生耐藥性, 但在四環素、卡那霉素和多西環素上存在不同耐藥性(鄧龍君, 2021; 楊超等, 2021)。而本研究發現該分離菌株LB2101對氟苯尼考、四環素、復方新諾明、新霉素、多粘菌素B和強力霉素等敏感, 對青霉素、乙酰螺旋霉素、羧芐西林和利福平等具有耐藥性, 這與鄧龍君(2021)和楊超等(2021)的試驗結果都有差異, 說明了不同菌株間存在藥物敏感性差異。造成菌株間耐藥性差異的原因, 這可能是與養殖地區和抗生素種類等因素相關, 不同養殖地區其抗菌藥物的使用類型、用藥劑量和用藥方式往往不同, 可能會形成不同的藥物環境或養殖環境。有研究證明, 病原細菌耐藥性與苗種來源、養殖水平和模式、菌株來源、分離年份及各地用藥習慣有關(胡大勝, 2010; 丁正峰等, 2011; 鄧玉婷等, 2019)。
3.4 大口黑鱸維氏氣單胞菌病的防治技術
鑒于維氏氣單胞菌其致病機理尚未清楚, 在養殖生產中主要以預防為主, 這其中防治的有效防治手段包括疫苗和免疫增強劑。水產上針對維氏氣單胞菌的疫苗已有研究, 如滅活疫苗(任燕等, 2019)、減毒疫苗(徐一軻等, 2019)、菌影疫苗(徐展等, 2014)、菌蛻疫苗(閆兵兵等, 2017)等。研究表明五味子、雞血藤、苦諫樹皮、丹參、烏梅、蘇木、五倍子、丁香、艾葉、石榴皮和地榆等中草藥, 對于維氏氣單胞菌有較好的抑制作用, 可用于預防維氏氣單胞菌感染(馬秀慧等, 2018; 朱成科等, 2018; 王寶屯等, 2021)。此外, 還可以通過調節魚體自身免疫力進行防控, 現已發現在飼料中添加益生菌(如干酪乳桿菌、屎腸球菌)和免疫調節劑(如百泰A、酵母細胞壁、牛磺酸)等可顯著增強魚體自身的非特異性免疫力, 有效提高抗維氏氣單胞菌感染的能力(郁歡歡等, 2014; 張海朋等, 2017; 鞠安琪等, 2018)。除此之外, 部分抗生素對維氏氣單胞菌也有著較強的抑菌效果。但抗生素濫用會導致水產養殖對象產生耐藥性, 而且抗生素殘留會引發水產品質量安全問題, 間接性危害人類健康, 目前很多抗生素已經列入禁用或限用范疇。因此, 在實際生產中對于維氏氣單胞菌的防治, 需要根據國家漁業用藥標準合理用藥, 以選用適宜的中草藥、益生菌等為宜, 實現綠色養殖, 做到減量用藥。
4 結論
本實驗從患病大口黑鱸肝臟組織中分離到1株病原菌, 通過形態學觀察、生理生化特性分析、16S rRNA序列和特定毒力基因分析對其進行鑒定, 并通過人工回歸感染試驗和藥敏試驗分析其致病性和耐藥性, 結果表明分離獲得的維氏氣單胞菌是一種導致鱸魚發病的條件性致病菌。研究結果揭示了維氏氣單胞菌菌株的部分生物學特性, 有利于豐富該菌屬的基礎數據, 為鱸魚養殖過程中的病害防控提供了有效的參考依據。
丁正峰, 薛暉, 王曉豐, 等, 2011. 江蘇主要水產病原菌耐藥譜系監測[J]. 江蘇農業科學, 39(2): 344-347.
馬秀慧, 姜海波, 羅學評, 等, 2018. 中草藥對羅非魚源維氏氣單胞菌體外抑菌效果研究[J]. 黑龍江畜牧獸醫(16): 174-176, 179.
王玉堂, 陳昌福, 呂永輝, 2013. 水產養殖動物致病菌耐藥性檢測數據實際作用[J]. 中國水產(10): 57-61.
王利, 魏勇, 2013. 鯽魚維氏氣單胞菌的分離鑒定及耐藥表型檢測[J]. 黑龍江畜牧獸醫(3): 101-103.
王寶屯, 顏遠義, 毛燦, 等, 2021. 82種中草藥及其復方對花鱸源維氏氣單胞菌的體外抑菌效果[J]. 華南師范大學學報(自然科學版), 53(3): 64-72.
王惠, 2014. 不同動物源性維氏氣單胞菌的生物特性及四種毒力基因比較研究[D]. 長春: 吉林農業大學: 19-23.
鄧玉婷, 譚愛萍, 張瑞泉, 等, 2019. 廣東主要水產養殖地區氣單胞菌耐藥狀況的調查分析[J]. 南方農業學報, 50(11): 2375-2383.
鄧龍君, 2021. 大口黑鱸維氏氣單胞菌的分離鑒定及其感染的病理損傷[J]. 河南農業科學, 50(1): 164-171.
東秀珠, 蔡妙英, 2001. 常見細菌系統鑒定手冊[M]. 北京: 科學出版社: 106-120.
田甜, 張建明, 杜合軍, 2017. 中華鱘源維氏氣單胞菌的分離鑒定及其藥敏特性[J]. 安徽農業大學學報, 44(6): 1011-1017.
邊宇, 錢宏偉, 孟慶峰, 等, 2013. 維氏氣單胞菌RCR快速檢測方法的建立及初步應用[J]. 中國獸藥雜志, 47(1): 1-3.
朱成科, 王建, 周燕, 等, 2018. 150種中草藥體外抑殺維氏氣單胞菌的藥效研究[J]. 淡水漁業, 48(1): 80-85, 96.
朱成科, 劉桂嘉, 張爭世, 等, 2017. 黃顙魚源維氏氣單胞菌Aha和gyrB基因雙重PCR檢測方法的建立[J]. 中國預防獸醫學報, 39(10): 810-814.
朱若林, 馬騰, 陳露, 等, 2017. 鳙魚維氏氣單胞菌的分離鑒定及其毒力基因檢測[J]. 安徽農業大學學報, 44(2): 229-233.
任燕, 時云朵, 曾偉偉, 等, 2019. 維氏氣單胞菌滅活疫苗對草魚免疫相關基因表達的影響及其保護效果[J]. 中國生物制品學雜志, 32(7): 726-731.
劉港彪, 王利, 王吉坤, 等, 2012. 齊口裂腹魚維氏氣單胞菌16S rDNA序列分析[J]. 水生態學雜志, 33(1): 92-96.
閆兵兵, 姜娜, 羅琳, 等, 2017. 維氏氣單胞菌菌蛻疫苗免疫后鯉免疫應答與基因表達特征[J]. 微生物學通報, 44(6): 1395-1404.
楊超, 董浚鍵, 劉志剛, 等, 2021. 大口黑鱸源維氏氣單胞菌的分離鑒定[J]. 南方水產科學, 17(3): 54-61.
宋明芳, 張冬星, 張海鵬, 等, 2018. 維氏氣單胞菌毒力因子的研究進展[J]. 中國獸醫科學, 48(8): 1038-1042.
宋路萍, 楊振蘋, 王斌, 等, 2020. 基于16S rDNA快速鑒定細菌的PCR測序方法的建立及其進化關系分析[J]. 中國醫藥科學, 10(17): 86-91.
張紅燕, 袁永明, 賀艷輝, 等, 2019. 中國淡水鱸養殖生產現狀分析[J]. 湖南農業科學(4): 112-114.
張海月, 康元環, 陳龍, 等, 2016. 不同動物源性維氏氣單胞菌OMPAⅠ基因的克隆及比較研究[J]. 中國獸醫科學, 46(5): 607-610.
張海朋, 谷巍, 鞠安琪, 等, 2017. 飼料中添加干酪乳桿菌對鯉魚非特異性免疫力和抗病力的影響[J]. 中國獸醫雜志, 53(7): 85-88.
陸夢瑩, 胡秀彩, 呂愛軍, 等, 2017. 鯰源維氏氣單胞菌的分離鑒定及藥敏特性[J]. 大連海洋大學學報, 32(5): 563-567.
陳亨利, 莫金龍, 康元環, 等, 2016. 藍曼龍魚致病性維氏氣單胞菌的分離鑒定與系統進化分析[J]. 中國獸醫雜志, 52(6): 115-118.
陳莉莉, 2018. 加州鱸飼料市場最全調查, 一文看懂如今產業現狀、痛點和契機[J]. 當代水產, 43(2): 80-81, 83.
郁歡歡, 薛敏, 韓芳, 等, 2014. 幾種免疫調節劑對花鱸生長性能、免疫力以及細菌感染后存活率影響的比較研究[J]. 動物營養學報, 26(8): 2386-2396.
國家特色淡水魚產業技術體系, 2021. 中國淡水鱸產業發展報告[J]. 中國水產(3): 40-48.
孟令緣, 牛沁雅, 廉魯昕, 等, 2021. 基于16S rDNA序列、MALDI-TOF-MS和VITEK的沙門氏菌和金黃色葡萄球菌的鑒定[J]. 中國食品學報, 21(10): 197-205.
趙武義, 2018. 加州鱸源維氏氣單胞菌的鑒定與分析[D]. 成都: 四川農業大學: 4-5.
胡大勝, 2010. 廣西水產養殖主要細菌病的監測與耐藥性調查[J]. 中國科技成果(13): 49-51.
胡天野, 吳同壘, 孟慶峰, 等, 2012. 框鏡鯉維氏氣單胞菌的生物學特性[J]. 中國獸醫雜志, 48(8): 18-21.
徐一軻, 孫愉宸, 盛強龍, 等, 2019. 維氏氣單胞菌的、及敲除菌株減毒活疫苗篩選[J]. 山東農業大學學報(自然科學版), 50(2): 186-190.
徐先棟, 李燕華, 王海華, 等, 2018. 臺灣泥鰍幼魚紅點病病原的分離鑒定及藥敏分析[J]. 水產科學, 37(4): 522-526.
徐展, 孟慶峰, 王偉利, 等, 2014. 維氏氣單胞菌菌影疫苗的構建及其溶菌動力學研究[J]. 中國預防獸醫學報, 36(12): 982-984.
高彩霞, 任燕, 王慶, 等, 2018. 草魚源致病性維氏氣單胞菌的分離鑒定及藥物敏感性分析[J]. 安徽農業大學學報, 45(3): 409-415.
黃文明, 王利, 冀國楨, 等, 2013. 胭脂魚維氏氣單胞菌和中間氣單胞菌的鑒定及藥物敏感性[J]. 水產科學, 32(4): 210-214.
藺凌云, 潘曉藝, 袁雪梅, 等, 2018. 淡水魚蝦細菌病診斷與防控技術研究進展[J]. 中國動物檢疫, 35(6): 83-88.
熊廣澤, 張焱, 2021. 魚源維氏氣單胞菌的研究進展[J]. 陜西農業科學, 67(9): 76-80.
黎炯, 葉星, 盧邁新, 等, 2011. 羅非魚維氏氣單胞菌的分離鑒定和藥敏試驗[J]. 水生態學雜志, 32(3): 132-136.
鞠安琪, 谷巍, 張海朋, 等, 2018. 屎腸球菌對鯉魚非特異性免疫酶活性和抗病力的影響[J]. 中國獸醫科學, 48(2): 228-233.
ABRAMI L, FIVAZ M, GLAUSER P E,, 2003. Sensitivity of polarized epithelial cells to the pore-forming toxin aerolysin [J]. Infection and Immunity, 71(2): 739-746.
FOYSAL J, MOMTAZ F, ALI H,, 2019. Molecular characterization and interactome analysis of aerolysin () gene from fish pathogen: The pathogenicity inferred from sequence divergence and linked to histidine kinase () [J]. Journal of Fish Diseases, 42(4): 465-475.
GAVíN R, MERINO S, ALTARRIBA M,, 2003. Lateral flagella are required for increased cell adherence, invasion and biofilm formation byspp [J]. FEMS Microbiology Letter, 224(1): 77-83.
HUSSEIN G H G, CHEN M, QI P P,, 2020. Aquaculture industry development, annual price analysis and out-of-season spawning in largemouth bass[J]. Aquaculture, 519: 734901.
KITA-TSUKAMOTO K, OYAIZU H, NANBA K,, 1993. Phylogenetic relationships of marine bacteria, mainly members of the family, determined on the basis of 16S rRNA sequences [J]. International Journal of Systematic and Evolutionary Microbiology, 43(1): 8-19.
LAZADO C C, ZILBERG D, 2018. Pathogenic characteristics ofisolated from the liver of a diseased guppy () [J]. Letters in Applied Microbiology, 67(5): 476-483.
LI T, RAZA S H A, YANG B T,, 2020.infection in commercial freshwater fish: a potential threat to public health [J]. Animals (Basel), 10(4): 608.
MERINO S, RUBIRES X, AGUILAR A,, 1997. The role of flagella and motility in the adherence and invasion to fish cell lines byserogroup O: 34 strains [J]. FEMS Microbiology Letters, 151(2): 213-217.
NAWAZ M, KHAN S A, KHAN A A,, 2010. Detection and characterization of virulence genes and integrons inisolated from catfish [J]. Food Microbiology, 27(3): 327-331.
YANG B T, ZHANG D X, WU T L,, 2019. Maltoporin (LamB protein) contributes to the virulence and adhesion ofTH0426 [J].Journal of Fish Diseases, 42(3): 379-389.
ISOLATION AND IDENTIFICATION OF PATHOGENICIN
LEI Ning1, HAO Gui-Jie2, HUANG Ai-Xia2, WANG Yu-Chen2, LIN Feng2, SHEN Xiao-Ming3, ZHU Jun-Jie1
(1. College of Life Science, Huzhou University, Huzhou 313000, China; 2. Key Laboratory of Healthy Freshwater Aquaculture, Ministry of Agriculture and Rural Affairs,Key Laboratory of Fish Health and Nutrition of Zhejiang Province, Institute of Freshwater Fisheries, Huzhou 313001, China; 3. Deqing Haoyuan Aquatic Seed Industry Co., Ltd., Huzhou 313200, China)
In China,is an important economic freshwater farmed fish species. At present, the healthy development ofaquaculture industry has been affected by many factors, including bacterial diseases.is an important pathogen causing septicemia and ulcer syndrome in freshwater fish. A bacterial strain was isolated and identified from liver of diseasedby morphological, physiological, and biochemical approaches, and 16S rRNA sequence and specific virulence gene analysis. The pathogenicity and drug resistance were analyzed by artificial regression infection test and drug sensitivity test. Results show that the strain was pathogenic, and it contained virulence genes such as,,,,, and. The strain was highly sensitive to 8 antibiotics (ceftriaxone, chloramphenicol, florfenicol, doxycycline, tetracycline, polymyxin, cotrimoxazole, and levofloxacin), moderately sensitive to 6 antibiotics (neomycin, enoxacin, gentamicin, streptomycin, azithromycin, and ciprofloxacin), and was resistance to other 8 antibiotics, as revealed in our drug sensitivity test. The bacterium was pathogenic to, and the median lethal dose was 1.4×106CFU/mL as shown in our artificial regression infection experiment.is a conditional pathogen in aquaculture, and some biological characteristics of this strain were documented, which is beneficial to enrich the basic data of, and provides some scientific basis and reference for the prevention and control of diseases inculture.
;; drug sensitivity test; virulence gene
S917.1
10.11693/hyhz20220200045
*浙江省重點研發計劃項目, 2019C02082號; 浙江省科技專項, 2021YSZX007號, 2021C02069-2-02號。雷 寧, 碩士研究生, E-mail: yleining@163.com
朱俊杰, 副教授, E-mail: zhjj@zjhu.edu.cn; 林 鋒, 副研究員, E-mail: wwlinfeng@163.com
2022-02-28,
2022-04-07
