基于碳氮穩定同位素的西北印度洋鳶烏賊攝食習性研究*
2022-09-21 02:26:18陸化杰歐玉哲陳新軍
海洋與湖沼 2022年5期
陳 靜 陸化杰, 2, 3, 4① 王 芮 歐玉哲 陳新軍, 2, 3, 4
基于碳氮穩定同位素的西北印度洋鳶烏賊攝食習性研究*
陳 靜1陸化杰1, 2, 3, 4①王 芮1歐玉哲1陳新軍1, 2, 3, 4
(1. 上海海洋大學海洋科學學院 上海 201306; 2. 自然資源部海洋生態監測與修復技術重點實驗室 上海 201306; 3. 國家遠洋漁業工程技術研究中心 上海海洋大學 上海 201306; 4. 農業農村部大洋漁業資源環境科學觀測實驗站 上海海洋大學 上海 201306)
內殼是頭足類重要的硬組織器官之一, 鳶烏賊()內殼的生長具有不可逆性, 是記錄鳶烏賊整個生活史信息的良好載體。根據2019年3~5月中國燈光罩網漁船在西北印度洋生產調查期間采集的鳶烏賊樣本, 利用碳氮穩定同位素技術, 研究了個體間的營養生態位關系、攝食習性等。結果表明, 雌、雄鳶烏賊個體的內殼13C不存在顯著性差異(>0.05), 而δ15N存在顯著性差異(<0.05)。營養生態位結果顯示, 雄性群體的生態位寬幅比雌性群體的要大。雌、雄個體的內殼生長對13C均沒有顯著影響(>0.05), 但雌、雄個體的內殼生長對15N均有顯著影響(<0.05),13C (1.28‰)和15N (3.46‰)值的變化表明鳶烏賊在生長過程中存在洄游現象, 營養等級發生改變, 且同時期的雌性個體較雄性個體捕食營養層級更高的食物。
鳶烏賊; 內殼; 穩定同位素; 攝食; 西北印度洋
鳶烏賊()廣泛分布在印度洋、太平洋的赤道和亞熱帶海域, 在西北印度洋分布尤其廣泛(葉旭昌等, 2004)。我國于2003年首次對西北印度洋鳶烏賊資源分布及漁場特征等進行了調查, 取得了良好效果。近幾年西北印度洋鳶烏賊已經是我國頭足類漁業重要的捕撈對象(邵鋒等, 2008; 王洪浩等, 2021)。近年來穩定同位素技術逐漸成為生態學研究中的一種重要手段, 相對于傳統胃含物分析法, 該方法局限性小, 并且可以對生物長期食性特征進行定量研究(Ibanez, 2008; 李云凱等, 2014)。生物體組織中的碳、氮穩定同位素可提供較長時間的攝食及物質能量傳遞等信息, 因此可以通過生物體硬組織連續取樣對個體或種群一定時期內的攝食習性和棲息環境進行研究(Guerra, 2010; 陸化杰等, 2014a; Liu, 2018;劉娜, 2020)。由幾丁質和蛋白質分子構成的內殼結構具有從尾椎向頭部生長的特性(Lorrain, 2011), 這一特性使其成為頭足類整個生活史信息記錄的良好載體(Cherel, 2009a), 可用于其漁業生物學的研究(Cherel, 2009b; Vander Zanden, 2010), 而且內殼結構具有形態穩定、易提取、研磨方便等優點, 越來越受到學者的關注(Lorrain, 2011; Queirós, 2019)。然而, 目前為止, 國內外尚沒有利用內殼碳氮穩定同位素開展西北印度洋鳶烏賊攝食生態的研究。鑒于此, 本研究根據2019年3~5月中國燈光罩網漁船在西北印度洋生產調查期間采集的鳶烏賊樣本, 對內殼葉軸進行連續切割, 對切割后各片段的碳、氮穩定同位素進行測定, 通過分析個體間的營養生態位關系和內殼片段上穩定同位素比值的連續序列, 探究鳶烏賊生長發育過程中攝食習性變化, 以期為后續利用內殼結構研究其攝食習性和種群特征等提供方法參考。
1 材料與方法
1.1 材料來源和生物學測定
本研究樣品采集于西北印度洋, 采集時間為2019年3~5月, 采集海域為西北印度洋13°12′~19°42′N、61°17~67°43′E, 共采集1 009尾樣本, 樣本經冷凍保存運回實驗室。樣本解凍后, 對其胴長(mantle length, ML)、體重(body weight, BW)等基礎生物學數據進行測量, 對性別、性腺成熟度等進行目測和鑒定, 測量時胴長精確至0.1 cm, 體重精確至0.1 g。

圖1 采樣站點圖
1.2 內殼處理
考慮樣本時空分布, 個體大小和性成熟等漁業生物學特性, 在不同采樣時間和采樣地點選取不同大小、性腺成熟度和不同性別的鳶烏賊樣本各6尾作為研究對象, 分別標記為S1, S2, S3, …, S12, 解剖后取出內殼, 分別標記為G1, G2, G3, …, G12。使用超純水對內殼進行清洗, 去除內殼表面殘留的有機物質。從早期生長端沿著生長方向葉軸中線每2 cm進行標記分段(Li, 2017), 測量每段內殼距離近端的長度(以生長方向為正值), 按標記將其葉軸沿V型生長紋進行切段處理(圖2), 分段將處理的內殼片段放入冷凍干燥機, 在–50 °C條件下冷凍干燥24 h, 使用混合型球磨儀將片段磨碎成粉末(Li, 2017)。

圖2 鳶烏賊內殼結構圖
1.3 穩定同位素分析
碳穩定同位素比值(13C)和氮穩定同位素比值(15N)的測定, 具體見文獻(貢藝, 2015)。每12個樣品中放入3個標準樣以確保實驗結果的精確度, 分析精度為0.05‰ (13C)和0.06‰ (15N) (Li, 2017)。穩定同位素測定在上海海洋大學穩定同位素分析實驗室進行。
1.4 數據處理
使用RStudio軟件中的siar程序包計算13C-15N雙位圖中各凸多邊形的面積(total area of convex hull, TA)和矯正標準橢圓面積(corrected the standard ellipse area, SEAc), 用來表示鳶烏賊的生態位寬幅和核心生態位, 并繪制出營養生態位圖(汪惠瓊等, 2020), 分析鳶烏賊不同個體之間的營養生態位關系; 對內殼片段穩定同位素序列進行分析, 對比分析雌、雄鳶烏賊群體間的差異, 并討論其群體攝食情況。
2 結果與分析
2.1 樣本組成
本研究所選取的12尾鳶烏賊體重范圍為311.9~ 479.5 g, 胴長范圍為21.5~25.3 cm。具體基礎生物學數據參見表1。
表1 鳶烏賊生物學參數

Tab.1 Biological parameters of S. oualaniensis
注: M: 雄性; F: 雌性
2.2 鳶烏賊內殼穩定同位素分析結果
通過內殼片段穩定同位素分析, 結果顯示內殼葉軸切割片段的13C變化范圍為–18.70‰~ –17.42‰ (1.28‰);15N的變化范圍為7.21‰~ 10.67‰ (3.46‰), 詳見表2。雄性個體13C值范圍為–18.67‰~–17.24‰ (1.43‰), 雌性個體13C值范圍為–18.70~–17.85‰ (0.85‰); 雌雄個體15N值變化范圍相近, 雄性個體7.21‰~10.22‰ (3.01‰), 雌性個體7.37‰~10.67‰ (3.30‰), 雌、雄個體13C、15N平均值的變化趨勢情況如圖3。方差分析(Analysis of variance, ANOVA)表明, 鳶烏賊雌、雄個體內殼13C不存在顯著性差異(>0.05),15N存在顯著性差異(<0.05)。
表2 鳶烏賊內殼穩定同位素值
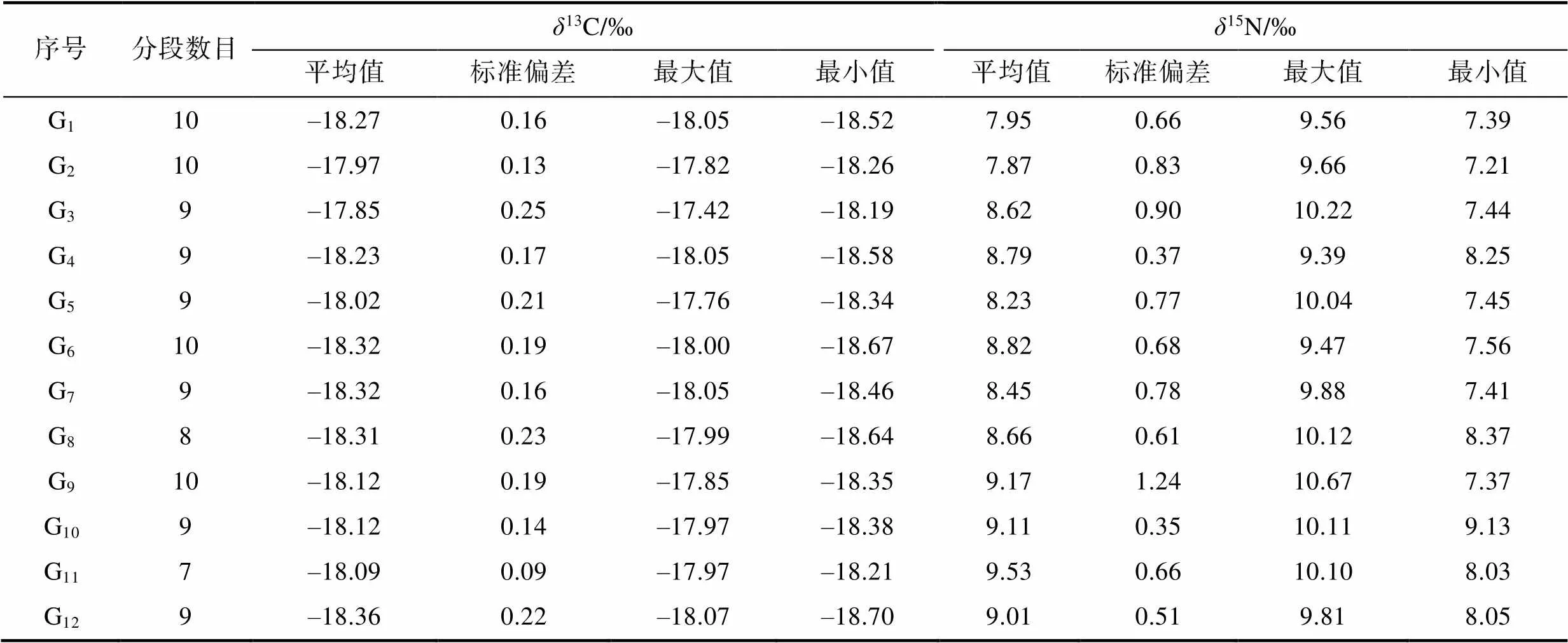
Tab.2 Isotope values of carbon and nitrogen in S. oualaniensis gladius

圖3 雌、雄個體δ13C、δ15N平均值的變化趨勢
2.3 鳶烏賊營養生態位
鳶烏賊內殼C、N穩定同位素分析結果可以反映個體間的營養生態位關系, 根據Hammerschlag-Peyer等(2011)提出的個體營養生態位變化研究框架, 以本研究中同位素測定結果數據繪制出營養生態位圖(圖4, 圖5)。

圖4 鳶烏賊個體間營養生態位的關系

圖5 鳶烏賊雌性和雄性群體營養生態位的關系
結果顯示, 12尾鳶烏賊的營養生態位均存在相互重疊的現象。雌性的生態位寬幅(TA)和核心生態位(SEAc)值分別為0.362‰和1.582‰; 雄性的TA和SEAc值分別為0.6152‰和2.352‰, 雄性群體的TA和SEAc值均大于雌性群體。
2.4 鳶烏賊生長對δ13C和δ15N的影響
ANOVA分析表明, 雌、雄個體內殼葉軸生長對13C均沒有顯著影響(>0.05), 但雌、雄個體內殼葉軸生長對15N均有影響顯著(=2.84,<0.05;=5.15,<0.05)。
13C值隨著內殼長度的增加呈現穩定趨勢, 而15N值前期上下波動較為平穩, 雌性個體在內殼長度生長到6 cm (內殼生長距近端–16 cm~–10 cm階段)后15N值呈現持續增大的趨勢, 當雄性個體內殼長度生長到8 cm (內殼生長距近端–18 cm~–10 cm階段)后15N值逐漸升高(圖6)。15N的變化表明其攝食對象和棲息環境在一定時期內發生了變化。
3 討論
3.1 鳶烏賊內殼穩定同位素分析
生物體內C、N穩定同位素比值受到生物個體攝食和生活環境等的影響。15N在不同營養級間具有富集效應, 因此可用于指示生物在食物網中的營養層次(Deniro, 1981),13C則可以反映出攝食變化與棲息環境的改變(李忠義等, 2005), 多用于追溯生物棲息地和洄游模式。Ruiz-Cooley等(2012)分析了大洋性頭足類肌肉13C值和15N值與緯度、離岸距離和胴長的關系, 結果表明13C值和15N值與個體生長和緯度變化無固定趨勢, 但均隨離岸距離增大而降低。通過對鳶烏賊內殼連續切割片段的碳、氮穩定同位素的分析, 可以了解鳶烏賊在不同生活時期的食性轉變以及棲息地環境的變化情況(Perez, 1996; Ruiz-Cooley, 2010)。鳶烏賊洄游規律雖沒有明確定論, 但相關研究表明, 在印度洋海域鳶烏賊個體大小隨著緯度變化由南向北增長趨勢顯著(Nesis, 1977; 陳新軍等, 2007)。在本研究中西北印度洋鳶烏賊在內殼生長長度<8 cm (內殼生長距近端–18 cm~–10 cm階段)的范圍內13C平均值變化范圍介于–18.28‰~ –18.03‰ (0.25‰) (圖3), 且不同個體13C值的分散程度逐漸增大, 這表明鳶烏賊幼體在產卵場附近水域攝食, 隨著個體的生長, 鳶烏賊行動能力增強, 其不同個體間活動和攝食方式有不同的選擇; 隨著個體繼續生長,13C值也逐漸增加, 表明此時期個體開始趨向于近岸水域攝食, 隨著個體增長鳶烏賊攝食對象發生了改變。與本研究結果相似, 在Merten 等(2017)對翼柄柔魚的研究中所有個體的13C值都大幅波動, 并在內殼長度達到20 cm后顯著增加。操亮亮等(2022)對秘魯公海莖柔魚的研究中13C值也發現相似的變化趨勢, 肌肉的13C值在胴長200~270 mm范圍內顯著減小, 胴長>270 mm后13C值升高, 表明其開始轉向近岸海域攝食, 與本研究中鳶烏賊內殼13C值發生變化的時期有所不同, 這可能是由頭足類不同群體間的生長差異造成的。
內殼片段中15N變化產生的原因主要是鳶烏賊食性發生改變, 另一原因是不同海域碳氮穩定同位素基線對鳶烏賊15N產生了影響。西北印度洋鳶烏賊內殼15N平均值在內殼長度<8 cm (內殼生長距近端–18 cm~–10 cm階段)范圍內變化很小, 表明早期幼體活動能力受限, 攝食對象結構單一; 隨著內殼生長,15N平均值的變化范圍介于8.10‰~9.88‰ (提升1.78‰), 鳶烏賊攝食能力隨著個體的生長逐漸提高, 攝食更高營養層級的食物。有研究認為, 鳶烏賊生活前期多以甲殼類生物為食, 隨著個體的生長而逐漸轉向攝食營養級較高的魚類和頭足類(Rodhouse, 2005)。這一研究結果與鳶烏賊的生存策略相符, 即攝食方式由早期階段隨水流進行被動隨機攝食逐漸轉化為主動選擇性的攝食(Stephens, 1986; Sajikumar, 2018), 這與謝嘉儀等(2021)對南沙海域鳶烏賊的營養級關系研究結論一致, 即隨著個體的生長營養級大小出現明顯增長的趨勢。隨著內殼長度的增加,15N值并沒有趨于平穩, 仍有增長的趨勢, 這可能與采樣環境中營養物質有關, 也可能是鳶烏賊個體尚未達到最高的攝食水平, 鳶烏賊的垂直移動也可能對15N值產生影響。有研究提出, 在鳶烏賊胃組織中微塑料含量與年齡有顯著的線性相關關系(陸化杰等, 2021a, 2022), 微塑料污染也可能對同位素測定產生影響, 后續可深入研究。

圖6 鳶烏賊個體碳、氮穩定同位素值隨內殼長度的變化
3.2 鳶烏賊營養生態位差異
自然存在的碳、氮穩定同位素可以反映食物來源和食物網中的營養水平信息(Hobson, 1992; Peterson, 1999), 利用內殼各片段15N和13C值繪制出鳶烏賊的營養生態位圖可以很直觀地反映個體之間生態位關系(Hammerschlag-Peyer, 2011)。Fang等(2015)比較北太平洋東、西部的柔魚群體角質顎中15N值, 發現兩個群體的攝食生態位分散, 重疊很小。而在本研究中12尾鳶烏賊的生態位相互有不同程度的重疊, 即可能在其生活史過程中, 個體的食物和生活環境相似。對雌、雄個體生態位分析結果表明雌雄生態位發生重疊, 說明雌雄個體在不同階段攝食對象的營養等級存在差異, 同時也可能存在群體間攝食競爭關系, 本研究與張宇美(2014)對南海鳶烏賊攝食習性的研究結果一致。通常生態位寬度越大表明其對空間食物資源的利用和對環境的適應能力越強, 本研究中雄性群體的TA和SEAc值均大于雌性群體, 表明雄性群體營養生態位寬幅較雌性群體范圍更廣, 陸化杰等(2014b)的研究中發現, 雄性樣本明顯少于雌性, 且雄性鳶烏賊個體較雌性個體偏小, 這些因素讓雄性鳶烏賊在種內競爭中處于劣勢, 因此雄性鳶烏賊需要拓寬生態位寬度以滿足生存需求, 這一結果與謝嘉儀等(2021)對南沙海域鳶烏賊的研究結果一致。而Kato等(2016)在分析莖柔魚穩定同位素時發現, 雌、雄莖柔魚群體在攝食、洄游過程存在差異, 雄性個體在較狹窄的緯度范圍內覓食, 與本研究中雄性群體占有更大的生態位恰好相反, 這可能與頭足類不同種類間攝食習性差異有關, 也可能與采樣季節和個體大小等有關, 后續還需要開展深入研究。
3.3 鳶烏賊生長對δ13C和δ15N的影響
鳶烏賊攝取食物體內的營養物質, 其中部分蛋白質分子和幾丁質構成了內殼的穩定結構(Perez, 1996; Ruiz-Cooley, 2010; 李云凱等, 2014)。有研究發現, 在海洋生態系統中海水溫度、光照強度和海水中CO2濃度等差異會導致基線生物的13C和15N發生時空變化(Germain, 2013; Ruiz-Cooley, 2013), 營養轉移對15N值的影響較為顯著(Mariotti, 1983), 因此洄游與攝食以及生物自身的生長代謝等過程也會對生物體內C、N穩定同位素比值產生不同影響(Deniro, 1981)。本研究中13C值在雌、雄群體間無顯著性差異, 可能是由于厄爾尼諾的影響, 特殊的氣候條件不僅會對西北印度洋鳶烏賊生長產生影響(陸化杰等, 2020; 汪惠瓊等, 2020; 陸化杰等, 2021b), 而且海水溫度等條件對鳶烏賊活動和攝食空間范圍也有限制, 使得鳶烏賊群體活動受限, 導致13C值在雌、雄群體間的差異不明顯。內殼生長對15N影響顯著(<0.05), 可能由于頭足類隨著胴長增加, 食性發生改變, 營養級提高導致的。雌性群體當內殼長度>6 cm (內殼生長距近端–16 cm~–10 cm階段)時15N值逐漸增加, 表明此時攝食水平開始提高, 對食物的選擇逐漸轉向營養等級較高的物種, 這一結果與張宇美等(2013)的研究相同。15N值在雌性個體中明顯高于雄性個體, 這表明雌、雄群體在對食物營養級的選擇上有所不同, 雌性群體攝食等級高于雄性群體, 這可能是因為雌性個體在生長階段和性腺發育過程中需要更多的能量累積。陸化杰等(2021b)的研究結果顯示不同年間鳶烏賊胃飽滿度雌性均高于雄性, 更多的營養獲取利于雌性個體生長。張宇美等(2013)對南海鳶烏賊的研究結果也表明, 雌性個體攝食等級以3、4級為主, 雄性則以2、3級為主, 雌性群體攝食等級高于雄性群體, 且雌、雄個體攝食等級均隨胴長的增加而增加, 這與本研究結果相同。結合何靜茹等(2021)對西北印度洋海域鳶烏賊角質顎形態研究, 西北印度洋鳶烏賊雌性個體的角質顎結構明顯大于雄性個體, 本研究中雌性群體內殼15平均值明顯高于雄性個體, 也進一步說明了鳶烏賊雌性個體在攝食能力和攝食水平上高于雄性個體。攝食營養層級的不同也可能與雌雄個體采樣時間和采樣位置不同有關, 在后續研究中將會深入探討。
4 結語
關于鳶烏賊生物學的研究多基于耳石、角質顎等硬組織結構展開, 到目前為止基于鳶烏賊內殼的研究少有報告。本研究通過對西北印度洋鳶烏賊內殼連續切段, 進行碳氮穩定同位素測定, 以定量的方法對鳶烏賊生長過程中食性轉換及洄游展開初步研究, 彌補了傳統胃含物研究方法的局限, 并進一步說明該方法在頭足類攝食及洄游方向研究的可行性。本研究受采樣時空跨度影響及缺少年齡數據, 僅反映出了鳶烏賊生活史過程中攝食和洄游的變化的大致時期, 下一步研究中可以對西北印度洋不同海域和不同季節鳶烏賊, 結合年齡數據對鳶烏賊攝食變化與洄游路徑等信息進一步探索。
王洪浩, 何濤, 陸化杰, 等, 2021. 性成熟和個體大小對西北印度洋鳶烏賊耳石形態的影響[J]. 水產學報, 45(8): 1350-1360.
葉旭昌, 陳新軍, 2004. 印度洋西北海域鳶烏賊生物學特性初步研究[J]. 上海水產大學學報, 13(4): 316-322.
劉娜, 2020. 赤道莖柔魚不同組織穩定同位素分析[D]. 上海: 上海海洋大學: 16-18.
貢藝, 2015. 基于內殼穩定同位素信息的秘魯外海莖柔魚攝食與洄游研究[D]. 上海: 上海海洋大學: 25-28.
李云凱, 貢藝, 陳新軍, 2014. 穩定同位素技術在頭足類攝食生態學研究中的應用[J]. 應用生態學報, 25(5): 1541-1546.
李忠義, 金顯仕, 莊志猛, 等, 2005. 穩定同位素技術在水域生態系統研究中的應用[J]. 生態學報, 25(11): 3052-3060.
何靜茹, 陸化杰, 陳炫妤, 等, 2021. 西北印度洋鳶烏賊角質顎外形變化的影響因素[J]. 應用生態學報, 32(5): 1881-1889.
汪惠瓊, 陳潔南, 李云凱, 等, 2020. 厄爾尼諾對柔魚亞科近緣種莖柔魚與鳶烏賊營養生態位的影響[J]. 海洋漁業, 42(5): 524-532.
張宇美, 2014. 基于碳氮穩定同位素的南海鳶烏賊攝食生態與營養級研究[D]. 湛江: 廣東海洋大學: 23-29.
張宇美, 顏云榕, 盧伙勝, 等, 2013. 西沙群島海域鳶烏賊攝食與繁殖生物學初步研究[J]. 廣東海洋大學學報, 33(3): 56-64.
陸化杰, 王從軍, 陳新軍, 2014b. 4-6月東太平洋赤道公海鳶烏賊生物學特性初步研究[J]. 上海海洋大學學報, 23(3): 441-447.
陸化杰, 王洪浩, 劉凱, 等, 2020. 厄爾尼諾發生期冬春季西北印度洋鳶烏賊耳石外形生長特性[J]. 生態學雜志, 39(11): 3694-3703.
陸化杰, 寧欣, 劉維, 等, 2021b. 不同氣候條件下南海西沙海域鳶烏賊()漁業生物學比較研究[J]. 海洋與湖沼, 52(4): 1029-1038.
陸化杰, 劉凱, 陳子越, 等, 2022. 南海西沙群島海域鳶烏賊胃組織微塑料沉積特性研究[J]. 海洋與湖沼, 53(1): 187-194.
陸化杰, 劉凱, 歐玉哲, 等, 2021a. 微塑料染污及其對不同棲息地、不同食性海洋魚類影響的研究進展[J]. 水產學報, 45(12): 2099-2111.
陸化杰, 陳新軍, 馬金, 2014a. 西北太平洋柔魚耳石微量元素[J]. 應用生態學報, 25(8): 2411-2417.
陳新軍, 劉金立, 2007. 利用形態學方法分析印度洋西北部海域鳶烏賊種群結構[J]. 上海水產大學學報, 16(2): 174-179.
邵鋒, 陳新軍, 2008. 印度洋西北海域鳶烏賊漁場分布與海面高度的關系[J]. 海洋科學, 32(11): 88-92.
謝嘉儀, 張麗姿, 吳文秀, 等, 2021. 南沙群島海域鳶烏賊攝食習性與營養生態位[J]. 水產學報, 45(12): 1993-2002.
操亮亮, 劉必林, 李建華, 2022. 基于穩定同位素技術的東南太平洋公海莖柔魚攝食生態分析[J]. 大連海洋大學學報, 37(1): 120-128.
CHEREL Y, FONTAINE C, JACKSON G D,, 2009a.Tissue, ontogenic and sex-related differences in13C and15N values of the oceanic squid(Cephalopoda: Ommastrephidae) [J]. Marine Biology, 156(4): 699-708.
CHEREL Y, KERNALéGUEN L, RICHARD P,, 2009b. Whisker isotopic signature depicts migration patterns and multi-year intra- and inter-individual foraging strategies in fur seals [J]. Biology Letters, 5(6): 830-832.
DENIRO M J, EPSTEIN S, 1981. Influence of diet on the distribution of nitrogen isotopes in animals [J]. Geochimica et Cosmochimica Acta, 45(3): 341-351.
FANG Z, XU L L, CHEN X J,, 2015. Beak growth pattern of purpleback flying squidin the eastern tropical Pacific equatorial waters [J]. Fisheries Science, 81(3): 443-452.
GERMAIN L R, KOCH P L, HARVEY J,, 2013. Nitrogen isotope fractionation in amino acids from harbor seals: implications for compound-specific trophic position calculations [J]. Marine Ecology Progress Series, 482: 265-277.
GUERRA á, Rodríguez-NAVARRO A B, GONZáLEZ á F,, 2010. Life-history traits of the giant squidrevealed from stable isotope signatures recorded in beaks [J]. ICES Journal of Marine Science, 67(7): 1425-1431.
HAMMERSCHLAG-PEYER C M, YEAGER L A, ARAúJO M S,, 2011. A hypothesis-testing framework for studies investigating ontogenetic niche shifts using stable isotope ratios [J]. PLoS One, 6(11): e27104.
HOBSON K A, WELCH H E, 1992. Determination of trophic relationships within a high arctic marine food web using13C and15N analysis [J]. Marine Ecology Progress Series, 84: 9-18.
IBANEZ C M, ARANCIBIA H, CUBILLOS L A, 2008. Biases in determining the diet of jumbo squid(D’Orbigny 1835) (Cephalopoda: Ommastrephidae) off southern-central Chile (34°S-40°S) [J]. Helgoland Marine Research, 62: 331.
KATO Y, SAKAI M, NISHIKAWA H,, 2016. Stable isotope analysis of the gladius to investigate migration and trophic patterns of the neon flying squid () [J]. Fisheries Research, 173: 169-174.
LI Y K, GONG Y, ZHANG Y Y,, 2017. Inter-annual variability in trophic patterns of jumbo squid () off the exclusive economic zone of Peru, implications from stable isotope values in gladius [J]. Fisheries Research, 187: 22-30.
LIU B L, LIN J Y, CHEN X J,, 2018. Inter- and intra-regional patterns of stable isotopes inbeaks: biological, geographical and environmental effects [J]. Marine and Freshwater Research, 69(3): 464-472.
LORRAIN A, ARGüELLES J, ALEGRE A,, 2011. Sequential isotopic signature along gladius highlights contrasted individual foraging strategies of jumbo squid () [J]. PLoS One, 6(7): e22194.
MARIOTTI A, 1983. Atmospheric nitrogen is a reliable standard for natural15N abundance measurements [J]. Nature, 303(5919): 685-687.
MERTEN V, CHRISTIANSEN B, JAVIDPOUR J,, 2017. Diet and stable isotope analyses reveal the feeding ecology of the orangeback squid(Steenstrup 1855) (,) in the eastern tropical Atlantic[J]. PLoS One, 12(12): e0189691.
NESIS K N, 1977. Population structure of the squid(Lesson, 1830) in the tropical West Pacific [J]. Trudy IOAN SSSR, 107: 15-29.
PEREZ J A A, O’DOR R K, BECK P,, 1996. Evaluation of gladius dorsal surface structure for age and growth studies of the short-finned squid,(Teuthoidea: Ommastrephidae) [J]. Canadian Journal of Fisheries and Aquatic Sciences, 53(12): 2837-2846.
PETERSON B J, 1999. Stable isotopes as tracers of organic matter input and transfer in benthic food webs: a review [J]. Acta Oecologica, 20(4): 479-487.
QUEIRóS J P, PHILLIPS R A, BAETA A,, 2019. Habitat, trophic levels and migration patterns of the short-finned squidfrom stable isotope analysis of beak regions [J]. Polar Biology, 42(12): 2299-2304.
RODHOUSE P G, 2005. C2. World squid resources [J]. Review of the State of World Marine Fisheries, 5: 175-187.
RUIZ-COOLEY R I, BALLANCE L T, MCCARTHY M D, 2013. Range expansion of the jumbo squid in the NE Pacific:15N decrypts multiple origins, migration and habitat use [J]. PLoS One, 8(3): e59651.
RUIZ-COOLEY R I, GERRODETTE T, 2012. Tracking large-scale latitudinal patterns of13C and15N along the E Pacific using epi-mesopelagic squid as indicators [J]. Ecosphere, 3(7): 1-17.
RUIZ-COOLEYR I, VILLA E C, GOULD W R, 2010. Ontogenetic variation of13C and15N recorded in the gladius of the jumbo squid: Geographic differences [J]. Marine Ecology Progress Series, 399: 187-198.
SAJIKUMAR K K, RAGESH N, VENKATESAN V,, 2018. Morphological development and distribution of paralarvae and juveniles of purpleback flying squid(Ommastrephidae), in the Southeastern Arabian Sea [J]. Vie et Milieu - Life and Environment, 68(2/3): 75-86.
STEPHENS D W, KREBS J R, 1986. Foraging Theory [M]. Princeton: Princeton University Press: 22-24.
VANDER ZANDEN H B, BJORNDAL K A, REICH K J,, 2010.Individual specialists in a generalist population: Results from a long-term stable isotope series [J]. Biology Letters, 6(5): 711-714.
FEEDING ECOLOGY OFBASED ON CARBON AND NITROGEN STABLE ISOTOPE ANALYSIS IN THE NORTHWEST INDIAN OCEAN
CHEN Jing1, LU Hua-Jie1, 2, 3, 4, WANG Rui1, OU Yu-Zhe1, CHEN Xin-Jun1, 2, 3, 4
(1. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China; 2. Key Laboratory of Marine Ecological Monitoring and Restoration Technologies, Ministry of Natural Resources, Shanghai 201306, China; 3. National Distant-water Fisheries Engineering Research Center, Shanghai Ocean University, Shanghai 201306, China; 4. Scientific Observing and Experimental Station of Oceanic Fishery Resources, Ministry of Agriculture and Rural Affairs, Shanghai Ocean University, Shanghai 201306, China)
Gladius is one of the most important hard tissues of cephalopods. The gladius is a good carrier for recording the whole life history of.samples were collected by Chinese light falling-net fishery in March to May 2019 in the northwest Indian Ocean, from which the trophic niche and feeding ecology ofwere studied using stable isotope technology. Results showed that no significant difference in13C between female and male individuals with gladius length (>0.05), but15N of male individuals had significant differences with gladius length (<0.05). The gladius growth of both males and females had no significant relation with13C content (>0.05), but a significant relation with15N (<0.05). The changes in13C (1.28‰) and15N (3.46‰) indicate a migratory phenomenon that the migration and trophic level ofwere improved as it grew. Furthermore, the females were in a higher trophic level than males.
; gladius; stable isotope; feeding habits; Northwest Indian Ocean
S931; Q958; X174
10.11693/hyhz20220200044
*國家重點研發計劃項目, 2019YFD090402號; 國家自然科學青年基金項目, NSFC 4150618號。陳 靜, 碩士研究生, E-mail: chenjing9802@163.com
陸化杰, 碩士生導師, 副教授, E-mail: hjlu@shou.edu.cn
2022-02-28,
2022-05-12
猜你喜歡
今日農業(2021年11期)2021-08-13 08:53:34
保健醫苑(2021年7期)2021-08-13 08:48:02
小讀者(2021年2期)2021-03-29 05:03:48
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
