浙江三門灣海域主要底上動物生態位及種間聯結性*
2022-09-21 02:28:48魏永杰王曉波韓慶喜
海洋與湖沼 2022年5期
孟 周 魏永杰 王曉波 韓慶喜
浙江三門灣海域主要底上動物生態位及種間聯結性*
孟 周1魏永杰2王曉波3韓慶喜1①
(1. 寧波大學海洋學院 浙江寧波 315832; 2. 國家海洋局寧波海洋環境監測中心站 浙江寧波 315012; 3. 禹治環境科技(浙江)有限公司 浙江寧波 315199)
大型底上動物參與海洋生態系統中的物理、化學和生物過程, 具有極高的研究價值。浙江三門灣是天然的半封閉海灣, 也是海洋生物重要的棲息、覓食和育幼的場所。為探究三門灣海域物種間的資源利用情況及生態關系, 在該海域進行了底拖網生物調查。根據2017年和2018年夏季在三門灣海域進行的大型底上生物研究結果, 運用優勢度()、生態位寬度、生態位重疊、方差比率法(VR)、2檢驗、聯結系數(AC)、共同出現百分率(PC)對生態位和種間聯結性進行了研究。結果表明: 2017~2018年間三門灣海域共捕獲大型底上動物53種, 包括兩年共有種, 即主要底上動物22種。其中優勢種3種, 包括哈氏仿對蝦()、中華櫛孔蝦虎魚()和棒錐螺(), 這三個物種屬中生態位種; 三門灣主要底上動物依據生態位寬度值劃分為3組, 即廣生態位種、中生態位種和窄生態位種; 種對間生態位重疊值總體差異性較大, 其與種對的食性、棲息環境密切相關; 根據總體聯結性分析得主要底上動物總體呈顯著正關聯, 群落結構較為穩定;2檢驗、聯結系數(AC)和共同出現百分率(PC)表明種對間聯結性較弱, 趨近于相互獨立。三門灣大型底上動物的群落結構比較穩定, 但種對間的關聯性在逐漸下降。
底上動物; 優勢種; 生態位; 生態位重疊; 種間聯結性
海洋底上動物(marine epifauna)是指生活在海洋底表, 固著或附著在堅硬基質(如巖礁、動物的外殼等), 或爬行在基底之上的生物生態類群(李新正等, 2010)。底上動物種類豐富, 具有運動性較差且生活環境相對穩定的特點, 通過對它們的物種組成和群落結構動態變化進行研究, 能清楚地展現海洋環境變化和污染狀況, 因此底上動物在環境污染監測中具有重要的指示作用(張均龍等, 2016)。底上動物與底內動物、底游動物合稱為底棲動物, 共同完成沉積物和水之間的顆粒和溶解養分交換, 即海底-中上層耦合作用(Kristensen, 2014)。不僅如此, 底棲動物還在生態系統的物質循環和能量流動過程中發揮著重要作用, 如有機質分解、生物地球化學循環和食物網內營養物質的轉移等(Constable, 1999; Carvalho, 2007)。大型底上動物是大型底棲動物(分選時無法通過0.5 mm篩網的底棲生物)中的主要類群, 主要以浮游動物、藻類以及顆粒有機物為食(劉曉收等, 2020), 參與海洋生態系統中的物理、化學和生物過程, 具有極高的研究價值。自20世紀90年代以來, 生態位理論成為生態學研究領域的熱點, 是現代生態學研究中的主要內容, 其反映生物在生態學的獨特地位, 是研究物種間、物種與環境間關系的重要理論依據, 被廣泛應用于植物、動物群落的研究(李凡等, 2018; 劉坤等, 2021)。
三門灣位于我國海岸的中心, 三面環山, 岸線蜿蜒曲折, 入灣河流眾多, 是天然的半封閉海灣。灣內海域遼闊, 氣候溫和濕潤, 加之入灣河流帶來豐富的營養物質, 為灣內海洋生物提供充足的食物, 三門灣也是海洋生物重要的棲息、覓食和育幼的場所(中國海灣志編纂委員會, 1992)。另外, 三門灣土地資源豐富, 沿岸灘涂面積可達130 km2, 是浙江省經濟貝類育苗和養殖的主要基地之一。20世紀80年代以來, 隨著社會經濟迅猛發展, 核電站建設、過度捕撈和污水排放等人類擾動活動進一步增強, 對三門灣近岸海域生態環境造成巨大破壞, 使得海域內漁業資源衰退, 赤潮和水體富營養化等災害頻發(聶振林等, 2018; 俞松立等, 2021)。生態位理論是進行資源修復、可持續利用和物種生境保護的重要依據, 被越來越多的學者應用于海洋生物群落的研究, 如劉惠等(2019)通過對杭州灣游泳動物生態位的分析, 揭示了游泳動物種間的資源利用情況及競爭關系; 徐曉群等(2013)分析了三門灣海域浮游動物優勢種后發現, 其空間生態位與物種生活類型、攝食情況密切相關; 蔣巧麗等(2021)借助生態位理論在時間、空間兩個維度上探討了該海域主要蝦蟹類的生態位寬度、生態位分化及其與環境因子之間的關系。以上雖然涵蓋不同海域的游泳動物、浮游動物及甲殼類的生態位研究, 但關于三門灣底上動物生態位和種間聯結性計算的研究未見報道。本文以2017年、2018年在三門灣海域內進行的兩次海洋底拖網數據為基礎, 探討該海域內的主要大型底上動物的生態位寬度和種間聯結性, 以期客觀展現三門灣海域大型底上動物的種間關系及其對環境資源的利用情況, 為三門灣海域環境治理與保護提供理論依據和數據支撐。
1 材料與方法
1.1 樣品的采集與處理
2017年和2018年8月(夏季), 在浙江北部三門灣海域租用漁船“浙三漁運80570”(2017年)和“浙三漁運80139” (2018年)進行大型底上動物樣品采集工作。調查區域為29°00′~29°20′N, 121°57′~121°92′E, 區域范圍內設置9個站位(圖1)。由于灣內沉積物較硬, 生物采集用網口寬為1.5 m的阿氏拖網進行, 每個站位進行一次拖網, 拖網時間為15 min, 拖速為3 kn。采集后的生物樣品經75%的酒精固定后裝入樣品瓶帶回實驗室處理。在實驗室, 進行沖洗挑揀處理, 在體視顯微鏡SZ61 (奧林巴斯, 東京)下進行分類鑒定, 主要種類應盡可能鑒定到種。鑒定計數完成后, 用精度為0.01 g的電子天平YP6002B (力辰邦西, 上海)對所有樣品進行稱重處理。樣品的采集、處理過程依據《海洋調查規范》(GB/T 12763.6- 2007)執行。

圖1 2017~2018三門灣采樣站位圖
1.2 數據處理
1.2.1 優勢度計算
=(n/)×f, (1)
式中,代表優勢度指數,為各個站位捕獲的底上動物的總個體數,n為第種生物的總個體數,f為該物種在每個調查站位出現的頻率。當物種的優勢度>0.02, 該物種即為優勢種(Mcnaughton, 1967)。
1.2.2 生態位寬度 借助Shannon指數(Shannon, 1963)完成計算:

式中,B代表該物種的生態位寬度;P為第種生物在第個站位的個體數目占第種生物全部個體數的比例;代表總站位數,B∈[0,]。
1.2.3 生態位重疊 借助Pianka指數(Pianka, 1973)完成計算:

式中,Q代表生態位重疊度;P和P為第種生物和第種生物在第個站位的個體數目占第種生物全部個體數的比例,代表總站位數。Q∈[0,1]。
1.2.4 總體聯結性 總體聯結性借助方差比率法(variance ratio, VR) (Schluter, 1984)來完成測定, 用來檢驗關聯顯著程度:


VR =2/2, (6)
= VR·, (7)
式中,為總物種數,P=n/為物種出現的頻率, 其中為總站位數,n則為物種出現的站位數。T為站位種出現的數目,代表站位中的平均物種數。當VR<1時, 物種間存在負相關; 當VR=1時, 物種間無關聯性; 反之當VR>1時, 物種間存在正相關。方差比率(VR)偏離1的顯著程度借助值進行判定: 若總體關聯性不顯著, 則2分布在90%的置信區間20.95() <<20.05(), 反之則關聯性顯著。
1.2.5 種對間聯結性 采用連續校正公式(王伯蓀等, 1985)來計算種對間聯結性, 公式如下:
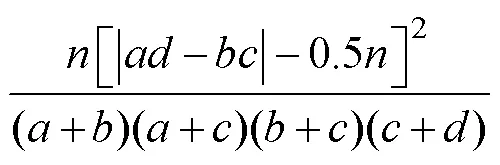
式中,為灣內的總站位數,為兩種對間同時出現的站位數;或則為出現其中一種的站位數;為兩種對都不存在的站位數。兩個物種間的正、負聯結性展現在-的正負性上。由卡方值分布表可知20.01(1)=6.6.35,20.05(1)=3.814。若2>6.635, 表現為極顯著聯結; 3.814<2<6.635, 表現為顯著聯結; 當2<3.814時, 聯結性不顯著。當其中某個物種的出現頻度為100%時, 它與其他物種組成種對后, 式中的、值將均為0, 但若將、值加權為1, 得到的結果較為客觀。
聯結系數(AC)公式(王伯蓀等, 1985)如下:
當≥時, AC=(–) / (+) (+), (9)
當>且≥, AC=(–) / (+) (+), (10)
當>且<, AC=(–) / (+) (+), (11)
式中, AC值的取值范圍為[–1, 1]。當AC值越接近1時, 表示種對間正聯結性越強; 當AC為0時, 表示種對間相互獨立; 反之當AC越接近–1時, 表示種對間負聯結性越強。
共同出現百分率(PC)公式(王伯蓀等, 1985)如下:
PC =/(++), (12)
式中, PC值的取值范圍為[0, 1]。當PC趨近于1時, 表示種間正聯結越緊密; PC等于0時說明種對間相互獨立。
1.3 數據分析
采樣站位圖借助ArcGIS Map 10.6繪制; 用R中的“spaa”、“ade4”包完成生態位寬度值、生態位重疊值以及種對間聯結的計算。底上動物生態位寬度分布圖使用GraphPad Prism 8.0繪制; 采用英國普利茅斯實驗室(Plymouth Marine Laboratory, UK)開發的PRIMER 6.0進行生態位寬度CLUSTER等級聚類圖的繪制。以上所有數據的整理和保存均在Microsoft Office Excel 2019中進行。
2 結果與分析
2.1 優勢種組成
對三門灣海域的兩次底拖網所得生物進行鑒定, 結果顯示2017、2018年共采集大型底上動物53種, 分屬6門10綱38科。在2017年出現32種生物, 由優勢種計算公式(>0.02)得出, 優勢種有5種, 包括甲殼動物中的哈氏仿對蝦()、中國毛蝦()和軟體動物的棒錐螺()、紅帶織紋螺(), 而魚類最少, 僅有中華櫛孔蝦虎魚()一種。2018年獲得底上動物48種, 明顯高于2017年, 2018年的7種優勢種中以魚類居多, 有孔蝦虎魚()、中華櫛孔蝦虎魚和龍頭魚()三種; 甲殼動物和軟體動物中各有2種, 分別是脊尾白蝦()、哈氏仿對蝦、棒錐螺以及縱肋織紋螺()其中哈氏仿對蝦、中華櫛孔蝦虎魚、棒錐螺為兩年優勢種, 優勢地位較為穩固(表1)。
表1 2017~2018年大型底上動物優勢種
2.2 生態位寬度
鑒于本次三門灣海域底上動物采樣獲得的物種較多, 本文結合大型底棲動物(彭松耀等, 2015; 湯雁濱等, 2016)和游泳動物(張苗苗等, 2018)的相關研究, 對采集到的優勢種和非優勢種進行篩選, 將兩次采樣同時出現的22種大型底上動物作為主要大型底上生物進行研究。通過生態位寬度公式計算后發現, 三門灣海域的主要大型底上動物生態位寬度值變化范圍在0.00~2.07之間, 不同物種間生態位寬度值差異明顯。根據生態位寬度值大小將之劃分成3段(圖2), 分別為a寬生態位(B≥2.0)、b中生態位(1.0≤B≤2.0)、c窄生態位(B≤1.0), 文中圖表出現的物種編號所對應的物種均與圖2相同。


圖2 主要大型底上動物生態位寬度分布

2.3 生態位重疊值
三門灣海域主要大型底上動物生態位重疊值Q如表2所示, 有231對底上動物種對, 其變化范圍在[0~0.95]之間。爪哇擬塔螺和海地瓜的生態位重疊值最高為0.95, 其次是拉氏紅狼牙蝦虎魚和龍頭魚為0.93, 說明它們利用資源的能力比較強且方式極為接近。其中Q>0.6 (生態位顯著重疊)共有58對, 占總對數的25.1%,Q>0.3 (生態位重疊有意義)共有151對, 占總對數的65.4%, 表明此次采樣的大型底上動物達到有意義的種對比例較高。生態位重疊值Q=0有16對, 占總對數的6.93%, 則說明小刀蟶和海仙人掌、毛蚶和爪哇擬塔螺等種對的資源利用能力較弱, 在本次采樣的站位分布少, 生態位重疊值低, 毛蚶與泥蚶、三疣梭子蟹、細巧仿對蝦等多個物種的生態位重疊值為0, 則表明毛蚶與它們利用環境資源的方式差異極大。

圖3 2017~2018三門灣生態位寬度聚類分析
表2 主要大型底上動物生態位重疊值
注: 表內物種編號同圖2
2.4 總體關聯性
借助方差比率法計算得出=2.35 (>1, 呈正相關), 則表示三門灣海域大型底上動物總體關聯性呈正相關, 表明該群落結構處于較為穩定的階段, 各物種對資源環境利用狀態較好, 生態系統功能結構也趨于成熟。值被用來檢驗偏離1的顯著程度,=21.16 (表3)不在置信區間20.95() <<20.05()之間, 22個主要大型底上動物物種間的總體關聯性顯著。
表3 主要大型底上動物種間的總體關聯性檢驗

Tab.3 General interspecific association test of major epifauna
2.5 種間聯結性
鑒于脊尾白蝦、紅帶織紋螺和中華櫛孔蝦虎魚在9個站位均有出現, 出現頻度為100%, 因此將三者在公式中的、值加權為1后,2檢驗的結果顯示(表4), 由22種大型底上動物組成的231個種對中, 其中有229對種對的聯結性不顯著, 種對間較為獨立, 占總對數的99.1%, 表明該海域各個物種間相互依賴程度低。僅縱肋織紋螺和龍頭魚、窄體舌鰨和窩紋網蝦蛄組成的2對種對的聯結性顯著, 且呈顯著正關聯, 占0.87%。
借助公式得聯結系數(AC)后, 結果如圖4所示, 表明在231個大型底上動物種對中, 種對間表現為正聯結的有78對, 占總對數的33.8%。其中有28對正聯結性較高(AC≥0.6), 占總對數的12.1%; 聯結性程度一般(0.2≤AC<0.6)的存在50對, 占總對數的21.6%; 種對間趨近于相互獨立(–0.2≤AC<0.2)的數目最多, 出現83對, 占35.9%; 呈現負聯結的種對數目最少, 有70對, 占總對數30.3%; 有32對種對的負聯結程度較高(AC<–0.6)的, 占13.9%; 負聯結程度一般(–0.6≤AC<–0.2)的種對與負聯結性較高種對的數目相近, 有38對, 占總對數的16.5%。
表4 主要大型底上動物2檢驗表

Tab.4 Main epifauna c2 inspection table
注: 表內物種編號同圖2; *加粗數字表示顯著正關聯

圖4 聯結系數AC半矩陣圖
共同出現百分率(PC)是灣內底上動物種對間正聯結性強弱的體現, 其中有16個種對表現為相互獨立(PC=0), 占總種對數的6.9%; 正聯結性最弱(0 生態位寬度反映生物對環境的適應性, 是研究某物種在一定層次上一定范圍內生存和發展所需的資源(物質、能量、空間等)以及它在該范圍內產生的影響的重要手段之一(張光明等, 1997; 韓曉鳳等, 2020)。通常所研究的生態位, 是指在特定尺度(時間、空間、生命層次)下物種的職能地位, 生態位寬度值越大, 則表明該物種的資源競爭力越強(王鳳等, 2006; 戴冬旭等, 2019)。本研究調查的主要大型底上動物的空間生態位寬度差異較大, 分段現象明顯(圖1), 分為a組、b組和c組, 其中a組脊尾白蝦和紅帶織紋螺生態位寬度值最大, 它們既是優勢種又是廣生態位種, 在調查的9個站位中的生物量分布較均勻。脊尾白蝦是近海廣溫、廣鹽分布種, 其食性廣而雜, 這種生活習性使得它具有較強的環境適應能力, 同時這也表明物種生態位寬度會受生活習性的影響(時冬晴等, 2007; 胡成業等, 2015)。三門灣是半封閉型海灣, 受波浪影響較弱, 屬沉積型海岸, 灘涂廣闊, 而近岸海域受沿岸流、臺灣暖流和黃海水團等水系的交匯影響(蔡如星, 1991), 鹽度偏低, 非常適宜紅帶織紋螺這種低鹽廣溫性物種的生存。b組中生態位分布物種數高于其余兩組, 該段物種數目占主要底上動物總物種數的59%。中華櫛孔蝦虎魚(1.86)、哈氏仿對蝦(1.76)、縱肋織紋螺(1.69)、半褶織紋螺(1.67)等13種底上動物較為均勻的分布在中生態位, 彼此間差距較小。其中中華櫛孔蝦虎魚在所有站位均有出現且屬于優勢種, 但仍是中生態位種, 這也驗證了有關蝦蟹類生態位的研究(丁朋朋等, 2019; 蔣巧麗等, 2021), 優勢種不一定分布在廣生態位。窄生態位的物種數較少(7種), 其中小刀蟶(0.95)最高, 爪哇擬塔螺(0.00)最低, 生態位寬度值差距大, 分布不均勻。爪哇擬塔螺僅存在于1個站位, 數目較少且分布局限性較大, 生態位寬度值低可能與其出現率和個體數較低有關。 圖5 共同出現百分率PC半矩陣圖 三門灣海域的主要大型底上動物中, 脊尾白蝦和紅帶織紋螺生態位寬度值最高, 它們在該海域具有較強的環境適應性和資源利用能力, 在競爭中處于優勢。而毛蚶、小莢蟶等7種主要底上動物處于窄生態位, 環境適應性差, 分布局限性較大。哈氏仿對蝦與紅帶織紋螺、拉氏紅狼牙蝦虎魚、龍頭魚等多個物種重疊值較高, 存在較強的競爭關系。種間聯結性結果顯示三門灣海域的底上動物群落結構已經趨于穩定, 能較好地利用環境資源, 灣內大多數物種近于相互獨立, 并未明顯的表現出正或負的聯結性。將生態位與種間聯結性相結合后發現, 它們與群落的更替演變密切相關, 可作為三門灣海域進一步制定生態保護和資源修復策略的理論基礎。同時依據物種獨特的種間關系, 制定科學的增殖放流策略, 也可用于維持生態系統穩定和海洋資源的可持續利用。 丁朋朋, 高春霞, 彭欣, 等, 2019. 浙江南部近海主要蝦類的時空生態位及種間聯結性[J]. 應用生態學報, 30(11): 3942-3950. 王鳳, 鞠瑞亭, 李躍忠, 等, 2006. 生態位概念及其在昆蟲生態學中的應用[J]. 生態學雜志, 25(10): 1280-1284. 王慶志, 張明, 滕煒鳴, 等, 2015. 毛蚶養殖生物學研究進展[J]. 大連海洋大學學報, 30(4): 437-443. 王伯蓀, 彭少麟, 1985. 南亞熱帶常綠闊葉林種間聯結測定技術研究——I. 種間聯結測式的探討與修正[J]. 植物生態學與地植物學叢刊, 9(4): 274-285. 中國海灣志編纂委員會, 1992. 中國海灣志第五分冊(上海市和浙江省北部海灣)[M]. 北京: 海洋出版社: 234-236. 鄧小艷, 劉建國, 郭朋軍, 等, 2018. 小洋山鄰近海域主要游泳動物生態位及種間聯結性[J]. 海洋學報, 40(1): 96-105. 白歡歡, 2018. 黃山松群落主要木本植物營養生態位和種間聯結特征研究[D]. 北京: 中國林業科學研究院: 1-9. 劉坤, 俞存根, 許永久, 等, 2021. 舟山群島東側海域春秋季主要甲殼類物種的空間生態位分析[J]. 浙江大學學報(理學版), 48(4): 450-460, 480. 劉曉收, 倪大朋, 鐘鑫, 等, 2020. 黃海大型底棲動物食物網結構和營養關系研究[J]. 中國海洋大學學報(自然科學版), 50(9): 20-33. 劉惠, 俞存根, 鄭基, 等, 2019. 杭州灣灰鱉洋海域秋季主要游泳動物生態位與種間聯結[J]. 生態學報, 39(11): 3828-3836. 湯雁濱, 廖一波, 壽鹿, 等, 2016. 南麂列島潮間帶大型底棲動物群落優勢種生態位[J]. 生態學報, 36(2): 489-498. 李凡, 徐炳慶, 呂振波, 等, 2018. 萊州灣魚類群落優勢種生態位[J]. 生態學報, 38(14): 5195-5205. 李新正, 劉錄三, 李寶泉, 等, 2010. 中國海洋大型底棲生物: 研究與實踐[M]. 北京: 海洋出版社: 8-20. 時冬晴, 葉建生, 2007. 脊尾白蝦的生物學特性及健康養殖技術[J]. 漁業經濟研究(5): 40-42. 張光明, 謝壽昌, 1997. 生態位概念演變與展望[J]. 生態學雜志, 16(6): 46-51. 張均龍, 史本澤, 趙峰, 等, 2016. 中國海洋底棲生物學發展回顧與展望[J]. 海洋科學集刊(1): 194-204. 張苗苗, 王詠雪, 田闊, 等, 2018. 臺州玉環北部沿岸海域主要游泳動物生態位和種間聯結性[J]. 應用生態學報, 29(11): 3867-3875. 張琳琳, 周永東, 蔣日進, 等, 2020. 浙江中南部近岸海域春季主要魚類空間生態位[J]. 應用生態學報, 31(2): 659-666. 胡成業, 水玉躍, 杜肖, 等, 2015. 浙江蒼南南部沿岸海域主要游泳動物生態位及種間聯結性[J]. 應用生態學報, 26(10): 3195-3201. 俞松立, 王詠雪, 韓曉鳳, 等, 2021. 三門灣海域漁業生物群落結構及營養級變化特征[J]. 大連海洋大學學報, 36(3): 478-487. 聶振林, 王詠雪, 胡成業, 等, 2018. 三門灣春秋季魚類群落特征及其與環境因子的關系[J]. 水產學報, 42(9): 1390-1398. 柴雪良, 張永普, 宣瑛, 等, 2008. 若干環境因子對成體小莢蟶存活的影響[J]. 海洋學研究, 26(1): 45-51. 徐曉群, 曾江寧, 陳全震, 等, 2013. 浙江三門灣浮游動物優勢種空間生態位[J]. 應用生態學報, 24(3): 818-824. 徐滿厚, 劉敏, 翟大彤, 等, 2016. 植物種間聯結研究內容與方法評述[J]. 生態學報, 36(24): 8224-8233. 梁靜香, 周永東, 王忠明, 等, 2020. 三門灣大型底棲動物群落結構及其與環境因子的關系[J]. 應用生態學報, 31(9): 3187-3193. 彭松耀, 李新正, 王洪法, 等, 2015. 南黃海春季大型底棲動物優勢種生態位[J]. 生態學報, 35(6): 1917-1928. 董靜瑞, 水柏年, 胡成業, 等, 2017. 溫州南部沿岸海域主要魚類的生態位及種間聯結性[J]. 應用生態學報, 28(5): 1699-1706. 蔣巧麗, 許永久, 鄭基, 等, 2021. 浙江披山海域主要蝦蟹類時空生態位及種間聯結性[J]. 應用生態學報, 32(7): 2604-2614. 韓曉鳳, 王詠雪, 求錦津, 等, 2020. 臺州南部近岸海域春秋季主要魚類生態位及其種間聯結性[J]. 水產學報, 44(4): 621-631. 蔡如星, 1991. 浙江動物志(軟體動物)[M]. 杭州: 浙江科學技術出版社: 1-8. 戴冬旭, 俞存根, 劉惠, 等, 2019. 南麂列島東側海域主要游泳動物生態位及種間聯結性[J]. 應用生態學報, 30(12): 4249-4258. CARVALHO S, BARATA M, GASPAR M B,, 2007. Enrichment of aquaculture earthen ponds with: Consequences for benthic dynamics and natural productivity [J]. Aquaculture, 262(2/3/4): 227-236. CONSTABLE A J, 1999. Ecology of benthic macro-invertebrates in soft-sediment environments: A review of progress towards quantitative models and predictions [J]. Australian Journal of Ecology, 24(4): 452-476. KRISTENSEN E, DELEFOSSE M, QUINTANA C O,, 2014. Influence of benthic macrofauna community shifts on ecosystem functioning in shallow estuaries [J]. Frontiers in Marine Science, 1: 41. MCNAUGHTON S J, 1967. Relationships among functional properties of Californian grassland [J]. Nature, 216(5111): 168-169. PIANKA E R, 1973. The structure of lizard communities [J]. Annual Review of Ecology and Systematics, 4(1): 53-74. SCHLUTER D, 1984. A variance test for detecting species associations, with some example applications [J]. Ecology, 65(3): 998-1005. SHANNON C E, WEAVER W, 1963. The Mathematical Theory of Communication [M]. Chicago: University of Illinois Press: 3-55. NICHE AND INTERSPECIES ASSOCIATION OF MAJOR EPIFAUNA IN SANMEN BAY OF ZHEJIANG PROVINCE, CHINA MENG Zhou1, WEI Yong-Jie2, WANG Xiao-Bo3, HAN Qing-Xi1 (1. School of Marine Sciences, Ningbo University, Ningbo 315832, China; 2. Marine Environmental Monitoring Centre of Ningbo, SOA, Ningbo 315012, China; 3. Yuzhi Environmental Technology (Zhejiang) Co., Ltd., Ningbo 315199, China) In order to explore the resource utilization and ecological relationship of epifauna in Sanmen Bay, the bottom trawl was carried out to collect the epifauna in this area. According to the investigation results in 2017 and 2018 in Sanmen Bay, Zhejiang Province, dominance (), niche breadth, niche overlap, the variance ratio method (VR), chi-square test, association coefficient (AC), and percentage of co-occurrence (PC) were adopted to study the role of ecological position and interpolation. The results showed that a total of 53 epifaunal species were collected in the Sanmen Bay during 2017~2018, and 22 epifauna were determined as major species because they were collected in both sampling years. Among the major species, 3 dominant species were identified, including,and, which were moderate niche breadth; all epifaunal species were divided into 3 groups, wide niche breadth species, moderate niche breadth and narrow niche breadth species. The overall difference between the coupling pair niche overlap values was significant, closely related to food and habitat; according to the overall coupling analyses, the major epifauna was generally positively related, and the community structure was relatively stable; chi-square test, association coefficient (AC) and percentage of co-occurrence (PC) showed there were weak interspecific associations, independent to each other. The community structure of Sanmen bay was relatively stable, but the association between coupling pairs showed slow decrease. epifauna; dominant species; niche width; niche overlap; interspecific association Q958.8 10.11693/hyhz20211200314 *國家自然科學基金項目, 42076156號, 41676139號。孟 周, 碩士研究生, E-mail: mz17805843093@163.com 韓慶喜, 副研究員, E-mail: hanqingxi@nbu.edu.cn 2021-12-07, 2022-02-123 討論
3.1 生態位寬度

3.2 生態位重疊

3.3 種間聯結性

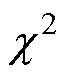
4 結論
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
保健醫苑(2021年7期)2021-08-13 08:48:02
趣味(語文)(2020年3期)2020-07-27 01:42:46
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
領導文萃(2015年4期)2015-02-28 09:19:05
