利用90K芯片分析煙農74(11)小麥衍生品種(系)的遺傳關系
2022-09-28 09:58:22翟曉靈王瑞霞張連焦娟李玉剛王圣健亓曉蕾蓋紅梅張勝全
中國農業科技導報 2022年8期
翟曉靈, 王瑞霞, 張連, 焦娟, 李玉剛, 王圣健, 亓曉蕾, 蓋紅梅*, 張勝全
(1.青島市農業科學研究院, 山東青島 266100;2.泰安市農業科學院, 山東泰安 271000;3.北京市農林科學院雜交小麥研究所, 北京 100097)
山東省煙臺市農業科學研究院于1974年以豐產、抗病、落黃優良的洛夫林13為母本, 以71(17)6-1-1株系為父本配制雜交組合, 系統選育出煙農74(11)及74(11)混-1-1-3和C149等優良品系。煙農74(11)含有洛夫林13、蚰包麥、農林10號、北陸13、小罌粟(農林89)、歐柔的血統。洛夫林13來自羅馬尼亞, 表現抗銹病和白粉病、耐高溫;蚰包麥是當地的當家品種, 表現抗病、高產;農林10號、小罌粟來自日本, 攜帶Rht1、Rht2矮稈基因, 表現抗旱、矮稈、抗條銹病;歐柔來自智利, 表現大穗、大粒、抗條銹[1-2]。由煙農74(11)育成了20世紀90年代初大面積推廣的魯麥14、魯麥13等品種, 2000年以后又育成了優質、高產品種煙農19。這些品種不僅在生產上發揮了很大的作用, 在小麥品種改良過程中也成為重要育種親本, 衍生了濟麥22、濟南17、泰農18等大面積推廣品種[3-5]。育種工作的突破性進展取決于關鍵遺傳資源的發現與創新[6]。
SNP(single nucleotide polymorphisms)標記于1996年由Lander提出[7], 是生物基因組中普遍存在的單核苷酸多態性, 具有密度高、代表性強、遺傳穩定性好和自動化程度高等優點, 得到專家學者的廣泛關注。目前, 在不同物種中已探索出30多種SNP檢測方法[8]。隨著基因芯片和測序技術的快速發展,SNP標記已成為更加高效的第3代分子標記[9]。并在人類[10-11]、果蠅[12]、大麥[13]、玉米[14-15]等的遺傳圖譜構建、關聯分析、QTL定位中發揮了重要作用。小麥基因組龐大, 其SNP研究滯后于水稻[16]、玉米[17]、大麥等物種[18]。2006年, 由Illumina公司研發的SNP GoldenGate芯片[19]大大推動了小麥基因組研究的發展, 目前, 小麥檢測SNP常用的方法有重測序、小麥55K芯片、90K芯片、660K芯片等, 并在遺傳多樣性[20-21]、數量性狀遺傳[22-24]、全基因組關聯分析[25-26]等方面得到廣泛應用。
本研究利用90K SNP芯片對煙農74(11)的衍生品種(系)、親本及近期育成小麥品種(系)進行了基因分型, 從系譜和分子水平探討煙農74(11)種質在山東省小麥育種中的重要作用, 以期為小麥品種的改良提供參考。
1 材料與方法
1.1 供試材料
供試小麥品種(系)120份(表1), 其中1~62號為煙農74(11)的衍生品種(系), 編號63~120為非煙農74(11)衍生品種(系), 用于進行遺傳多樣性的對比分析。

表1 供試品種(系)Table 1 List of tested wheat variety(line) 續表Continued

表1 供試品種(系)Table 1 List of tested wheat variety(line)
1.2 研究方法
1.2.1 DNA提取 供試材料種植于田間, 在幼苗期剪取2片幼葉冷凍保存, 并標記取樣單株。將冷凍樣品用液氮研磨后, 用試劑盒(天根生化科技有限公司)提取基因組DNA[27], 并用NanoDrop Spectrophotometer(ND-1000)檢測DNA樣品濃度和質量。
1.2.2 小麥90K芯片分析和SNP分型 供試材料DNA的Illumina 90K芯片分析由北京博奧晶典生物技術有限公司完成。采用Genomestudiov2011.1軟件包對結果文件*.bmp和*.idat進行SNP數據校正和分型[28]。數據校正基于芯片的原始數據, 即Genomestudiov2011.1軟件生成的每個SNP位點的聚類圖, 如果聚類圖分為2個組群(無雜合位點)或3個組群(有雜合位點), 而且每組表現為緊密簇擁在一起, 則這些聚類圖表示的SNP位點為有效SNP位點, 可用于供試材料的遺傳多樣性分析;如果聚類圖分為雜亂無章的多個組群, 則判定為無效SNP位點, 不能用于供試材料研究, 需在Genomestudiov2011.1軟件中, 將無效SNP位點標記出來, 待全部位點全部校正完, 導出SNP基因型數據, 將其中標記無效的位點去掉, 即完成全部SNP位點校正。
1.2.3 遺傳多樣性分析 利用Excel軟件進行SNP分型數據的整理和存儲, 采用自主研發程序將原始SNP數據的AA、AB、BB基因型轉換為Powermarker3.25軟件[29]需要的輸入格式。利用PowerMarker 3.25軟件分析多態性信息含量(polymorphism information content, PIC)、基因多樣性、每個SNP位點的等位變異數、主效等位變異頻率、期望雜合度及遺傳距離, 并計算遺傳相似性系數。其中, PIC、基因多樣性等用于評估群體的遺傳多樣性, 遺傳相似性系數用于評估2個供試材料之間的遺傳關系。

2 結果與分析
2.1 煙農74(11)種質對山東省冬小麥品種的支撐作用
由圖1可看出, 以煙農74(11)種質為親本, 選育出了魯麥14、魯麥13、煙農19等大面積推廣品種。這些品種不僅在小麥生產中大面積推廣應用, 還在小麥育種中發揮了重要作用。
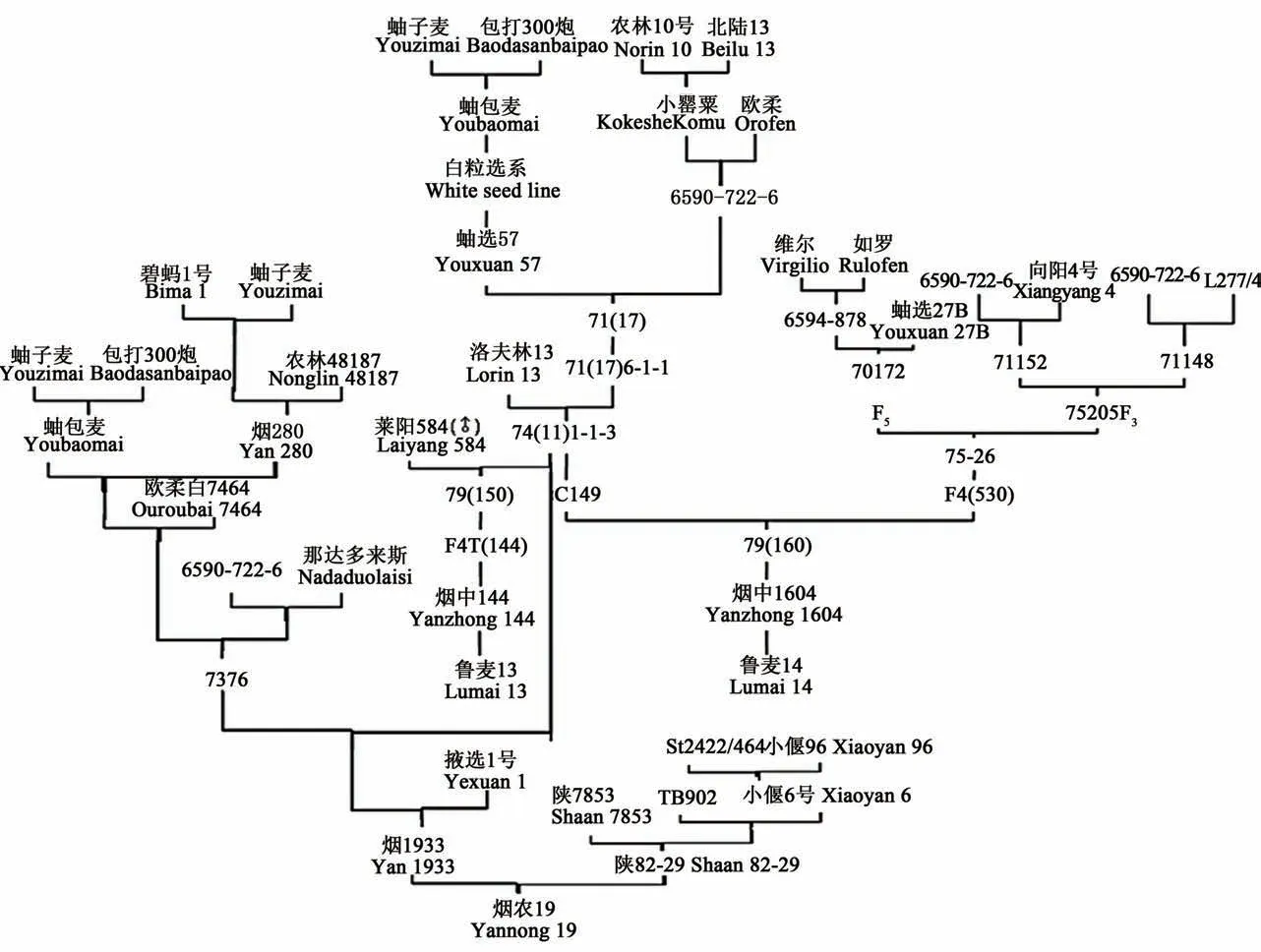
圖1 小麥關鍵種質煙農74(11)的系譜追蹤及主要衍生品種Fig.1 Pedigree tracking of 74(11)and the principal derived variety
對2000—2020年山東省審定小麥品種進行了系譜溯源, 21世紀以來20年間共審定小麥品種197個, 有55.33%的品種含有煙農74(11)種質血統, 36.04%的品種含有魯麥14血統, 21.32%的品種含有魯麥13血統, 10.66%含有濟麥22血統, 4.57%含有煙農19血統。由圖2看出, 不同年代材料在育種中發揮的作用不同, 就煙農74(11)種質而言, 在國家“十五”到“十三五”計劃期間, 其直接衍生的品種占比分別為28.57%(36個品種)、66.67%(36個品種)、62.07%(29個品種)、58.76%(97個品種), 表明該種質對山東省小麥新選育品種具有系譜支撐作用。此外, 魯麥14作為煙農74(11)種質的直接衍生品種, 在4個五年計劃中的衍生品種占比分別為20.00%, 41.67%、37.93%和39.18%, 表明魯麥14在小麥育種中成為重要的育種親本。
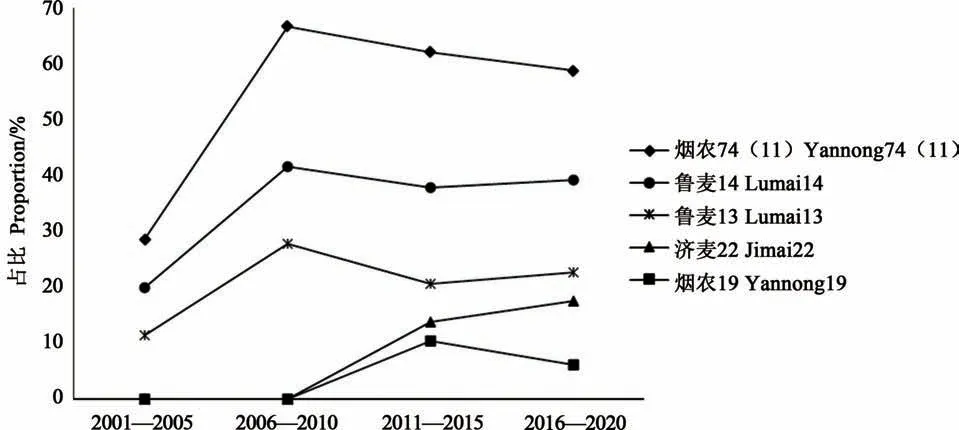
圖2 2000—2020年煙農74(11)及其衍生品種(系)在山東省的衍生材料統計Fig.2 Statistics of wheat materials derived from Yannong 74(11)and the principal variety(line)inShandong province from 2000 to 2020
2.2 供試材料的SNP分型及遺傳多樣性分析
120份供試材料共獲得有雜交信號的SNP位點81 587個, 校正后獲得有效SNP位點26 026個, 有染色體定位的SNP位點20 953個, 其中A基因組8 373個, B基 因 組10 592個, D基 因 組1 988個, 覆蓋小麥基因組3 646.01 cM。
小麥90K芯片獲得的SNP基因型數據在染色體間分布非常不均勻, 變異系數為58.17%, SNP個數最少的染色體是4D(73個), 最多的是1B(1 984個), 且在3個基因組中, 均為D基因組的SNP個數最少。SNP標記的平均等位變異數為2.28個, 變異系數為1.80%, 變幅很小。平均主效等位基因頻率為0.77, 平均基因多樣性為0.31, 平均期望雜合度為0.03。平均多態性信息含 量(PIC)為0.25, 變 幅 為0.18(4D)~0.31(2D), 明顯低于SSR標記揭示的遺傳多樣性(表2)。
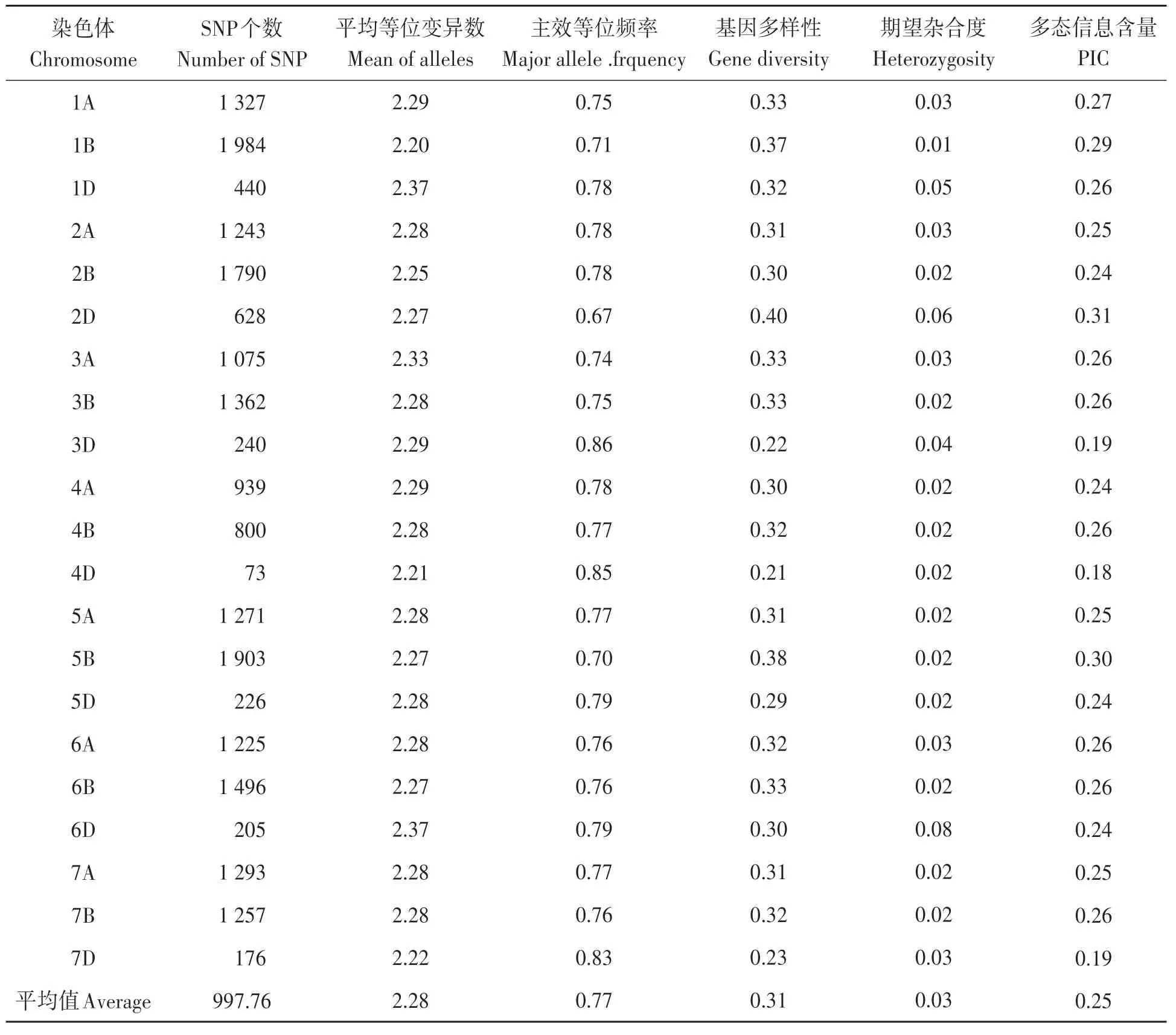
表2 供試材料在染色體水平的遺傳多樣性Table 2 Genetic diversity of SNP marker at chromosome level with tested materials
2.3 不同供試材料分組的遺傳關系
對供試材料進行了分組, 分為全部材料組(組Ⅰ)和煙農74(11)種質衍生材料組(組Ⅱ)(圖3)。對2組供試材料分別進行了遺傳相似性系數的計算, 組Ⅰ共有7 140個兩兩組合模式, 遺傳相似性系數范圍為0.56~1.00, 平均為0.70;根據遺傳相似性系數區間對材料進行了統計分析:0.51~0.60的 材 料 占0.66%、0.61~0.70的 占59.44%、0.71~0.80的占32.98%、0.81~0.90的占6.12%、≥0.91的占0.80%, 近60%的材料間遺傳相似性系數在0.61~0.70區間, 表明組Ⅰ材料的遺傳關系相對較遠。組Ⅱ材料之間共有2 219個兩兩組合模式, 遺傳相似性系數范圍為0.61~1.00, 平均為0.75;遺傳相似性系數區間在0.61~0.70的材料占26.77%、0.71~0.80的占53.22%、0.81~0.90的占17.26%、≥0.91的占2.75%, 表明僅含有煙農74(11)種質衍生后代之間的遺傳相似性系數一半以上為0.71~0.80, 近20%的材料組合為0.81~0.90, 表明組Ⅱ的材料兩兩間遺傳關系比組Ⅰ更近。

圖3 供試材料兩兩之間的遺傳相似性系數統計Fig.3 Statistical of genetic identity coefficient between every two tested materials
2.4 典型組合遺傳關系
煙農74(11)種質衍生后代中系譜關系比較完整且在小麥生產中大面積應用的小麥品種與親本之間的遺傳關系如表3所示。主要雜交組合為:魯麥13/臨汾5064、魯麥14/豫麥13、魯麥14/魯884187、魯麥13/豫麥2號。由于濟麥22的父本材料未得到妥善保存, 故只分析了濟麥22與其母本935106的遺傳關系。分析發現, 這4個雜交組合的父母本之間的遺傳相似性系數為0.69、0.65、0.68、0.71, 平均為0.68;子代與上緣親本間的遺傳相似性系數為0.67~0.87, 平均為0.79;姊妹系之間的遺傳相似性系數為0.84~0.87, 平均為0.855。濟麥22與母本的遺傳相似性系數為0.87, 與骨干親本魯麥14的遺傳相似性系數為0.85。從父母本之間的遺傳關系可以看出, 這些組合雙親的遺傳關系相對較遠;從子代與親本的遺傳關系可以看出育種中存在偏向性選擇現象, 其中子代與關鍵親本的遺傳相似性系數達0.80以上, 甚至0.85以上。

表3 基于小麥90K芯片的典型雜交組合親本及后代的遺傳關系Table 3 Genetic relationship of typical test material based on wheat 90K array 續表Continued
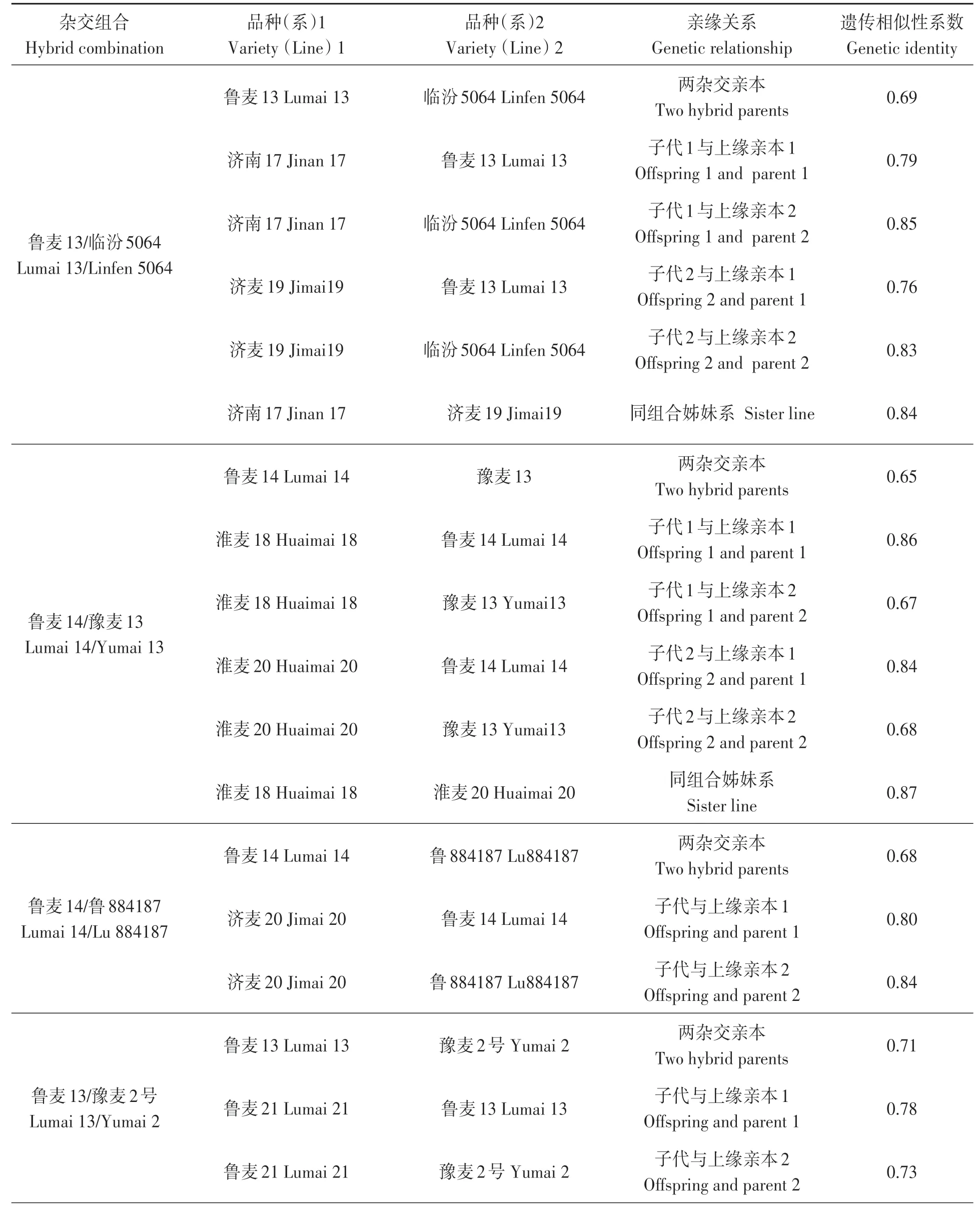
表3 基于小麥90K芯片的典型雜交組合親本及后代的遺傳關系Table 3 Genetic relationship of typical test material based on wheat 90K array
3 討論
本文對120份供試小麥材料進行了90K芯片分析, 結果表明,這些供試材料的平均遺傳相似性系數為0.70, 如果排除蚰包麥、農林10號等早代育種親本, 則當代育成品種(系)的遺傳關系更近:62份煙農74(11)種質的衍生后代間的平均遺傳相似性系數為0.75。當前, 極端災害天氣頻發, 病蟲害日益加重, 遺傳種質的單一化, 容易導致抗災能力減弱, 甚至有“全軍覆沒”的風險。劉建軍等[30]認為, 由于長期對產量、抗性、品質等性狀的持續定向選擇, 導致了當代育成品種遺傳多樣性急劇下降。遺傳多樣性的降低在一定程度上阻礙了小麥突破性品種的選育, 從而阻礙小麥育種的發展和潛力的提高。李邦發[31]指出, 繁6及其衍生品種在四川占統治地位30年, 但由于2001年條銹病大流行, 超過90%的衍生品種感染條銹病, 造成小麥生產巨大損失, 在育種領域應引以為戒。因此, 應擴大小麥育種材料的遺傳基礎, 尋找更加多樣的優良骨干親本。
煙農74(11)種質來自20世紀50、60年代的蚰子麥衍生的蚰包麥, 其在山東省連續推廣種植15年, 是黃淮麥區首個產量超過7 500 kg·hm-2的小麥品種[5], 被莊巧生先生列為16個骨干親本之一[1]。張學勇等[32]認為, 早期的品種或骨干親本無論當時多么優秀, 其田間表現和遺傳背景與今天的優良品種或骨干親本相比都有很大差距, 這反映了小麥品種改良是早期品種的繼承和符合現代目標的優良基因積累的過程。蚰子麥→蚰包麥→煙農74(11)種質→魯麥14→濟麥22的衍變過程, 也表明不同年代的當家品種首先是在繼承和綜合中發展, 必需有經過生產檢驗的品種基因組為基礎;此外, 從一代當家品種發展到新一代當家品種, 必需有突破性的進展, 必需解決當家品種與生產實際之間的主要矛盾。
致謝:感謝煙臺市農業科學研究院方正先生在煙農74(11)種質的特征特性及育種地位等方面給予的指導。
