Cx43通過Erk1/2促進BMP-2誘導的人牙髓細胞成牙本質分化
2022-09-28 13:46:44丁燦燦朱浩張安妮何圓培聶敏海李適廷
實用口腔醫學雜志 2022年5期
丁燦燦 朱浩 張安妮 何圓培 聶敏海 李適廷
骨形態發生蛋白(bone morphogenetic protein,BMP)-2是轉化生長因子-β超家族成員之一,它可以通過與胞膜上的受體相結合,激活胞質中Smad1/5/9復合體磷酸化及其核轉運,促使靶基因轉錄,誘導骨細胞增殖、膠原分泌和骨形成等[1]。此外,BMP-2也可以激活MAPK通路(如:Erk1/2、p38和JNK),調節細胞和組織功能[2]。在牙髓損傷后修復過程中,BMP-2參與調控成牙本質分化標志性基因牙本質涎磷蛋白(dentin sialophosphoprotein,DSPP)的表達,并通過Smad1/5通路促進人牙髓細胞(human dental pulp cells,hDPCs)成牙本質分化[3]。
連接蛋白(connexin,Cx)43由Gja1基因編碼,是機體表達最廣泛的跨膜連接蛋白,它構成縫隙連接通道(gap junction,GJ)或半通道(hemichannel,HC),促使細胞間或胞內外傳遞分子量小于1 200的物質,介導多種組織細胞的生理和病理活動[4]。生理狀態下,Cx43主要表達于牙髓組織內的成牙本質細胞層;在牙齒損傷過程中,成牙本質細胞排列模式發生輕微改變伴隨Cx43的表達上調,并且在修復性牙本質形成過程中Cx43高表達于規則排列的成牙本質細胞樣細胞內[5-6],表明Cx43參與調控了牙髓組織的損傷和修復過程。研究發現,胞外Ca2+通過BMP-2介導Smad1/5/9和Erk1/2通路促進hDPSCs成牙本質分化,Cx43介導Erk1/2信號促進胞外Ca2+誘導的hDPSCs成牙本質分化,并且Cx43的功能實現可能與其介導的GJ有關[7-8]。
以上研究表明,BMP-2和Cx43在hDPCs成牙本質分化過程中均起到重要的調控作用,并且均可通過Erk1/2信號發揮生物學功能。然而,兩者在這一過程中是否相互調控還不明確。為此,本研究在hDPCs中過表達Cx43并施以BMP-2刺激,探討BMP-2和Cx43在hDPCs成牙本質分化過程中的聯系。
1 資料與方法
1.1 主要試劑
pLVX-GJA1-EGFP-3FLAG慢病毒(lv-Cx43)和對照空病毒(mock)(上海生博);DSPP抗體(Santa cruz公司,美國);Cx43和osterix抗體(Abcam公司,英國);p-Smad1/5/9和p-Erk1/2抗體(CST公司,美國);rhBMP-2、PD98059(Sigma公司,美國)等。
1.2 hDPCs提取和培養
選擇14~28 歲患者因正畸或阻生拔除的完整、健康牙齒。收集樣本前,所有患者知情同意且實驗方案獲得西南醫科大學附屬口腔醫學院倫理委員會批準(批號:20180511001)。hDPCs參照文獻報道的方法培養細胞[9]。以5×104/mL密度接種于無菌塑料培養瓶,細胞培養液為α-MEM+10%FBS,培養條件為100%濕度、37 ℃、5%CO2孵箱。每3 d換液。實驗所用hDPCs為3~10 代。BMP-2刺激濃度為300 ng/mL,刺激時間為1 h[3]。
1.3 細胞轉染
將hDPCs接種培養皿的30%~40%,更換為含有lv-Cx43和mock的培養液(滴度分別為40和20)感染細胞24 h,隨后更換為生長培養基孵育3 d,0.6 μg/mL嘌呤霉素篩選陽性細胞5 d,qRT-PCR和Western blot檢測轉染效率。
1.4 實時定量聚合酶鏈式反應(qRT-PCR)
hDPCs培養至接近融合,提取細胞內總RNA。將0.5 μg RNA去除gDNA 污染,反轉錄cDNA,于-80 ℃冰箱保存、備用。qRT-PCR反應按照產品說明書使用(Takara公司)。應用PRISM?7500 Real-time PCR System(ABI,美國)進行擴增反應。每一樣品做3 個復孔,取其平均值。引物序列見表1。
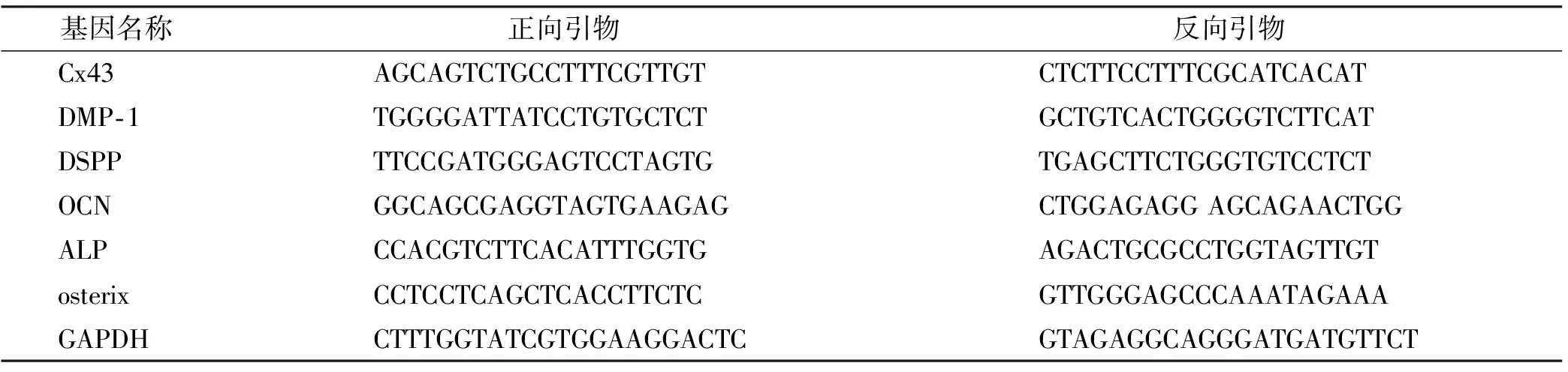
表1 qRT-PCR所用引物序列
1.5 蛋白質印跡法(Western blot)
提取細胞總蛋白,BSA法檢測蛋白質濃度。取等量蛋白裂解液(50~100 μg)進行SDS-PAGE電泳、轉膜和封閉;分別加入p-Smad1/5/9(1∶1 000)、p-Erk1/2(1∶1 000)和osterix(1∶500)一抗4 ℃過夜,TBST洗膜,浸入二抗稀釋液,37 ℃孵育2 h,Image quant Las 4000數字成像系統采集影像,以β-actin為內參進行分析。蛋白條帶使用Image J軟件定量分析。
1.6 免疫熒光法(IF)
hDPCs接種于蓋玻片,4%多聚甲醛固定20 min,0.1% Triton X-100可滲透化處理15 min,1% BSA封閉1 h。隨后,細胞孵育一抗(DSPP 1∶10),4 ℃過夜,TBST沖洗,孵育FITC熒光二抗(1∶300)1.5 h,細胞核DAPI 復染5 min。激光共聚焦顯微鏡(upright DM4000B,Leica,德國)觀察細胞熒光強度。
1.7 統計學分析
2 結 果
2.1 過表達Cx43上調hDPCs內Cx43表達
轉染mock和lv-Cx43的hDPCs培養7 d,細胞接近融合,形態呈梭形,鏡下呈綠色熒光(圖1A);分別提取轉染后的hDPCs RNA和蛋白,qRT-PCR和WB檢測結果顯示,過表達Cx43顯著上調hDPCs內Cx43 RNA(P<0.05)和蛋白水平(圖1B~C)。
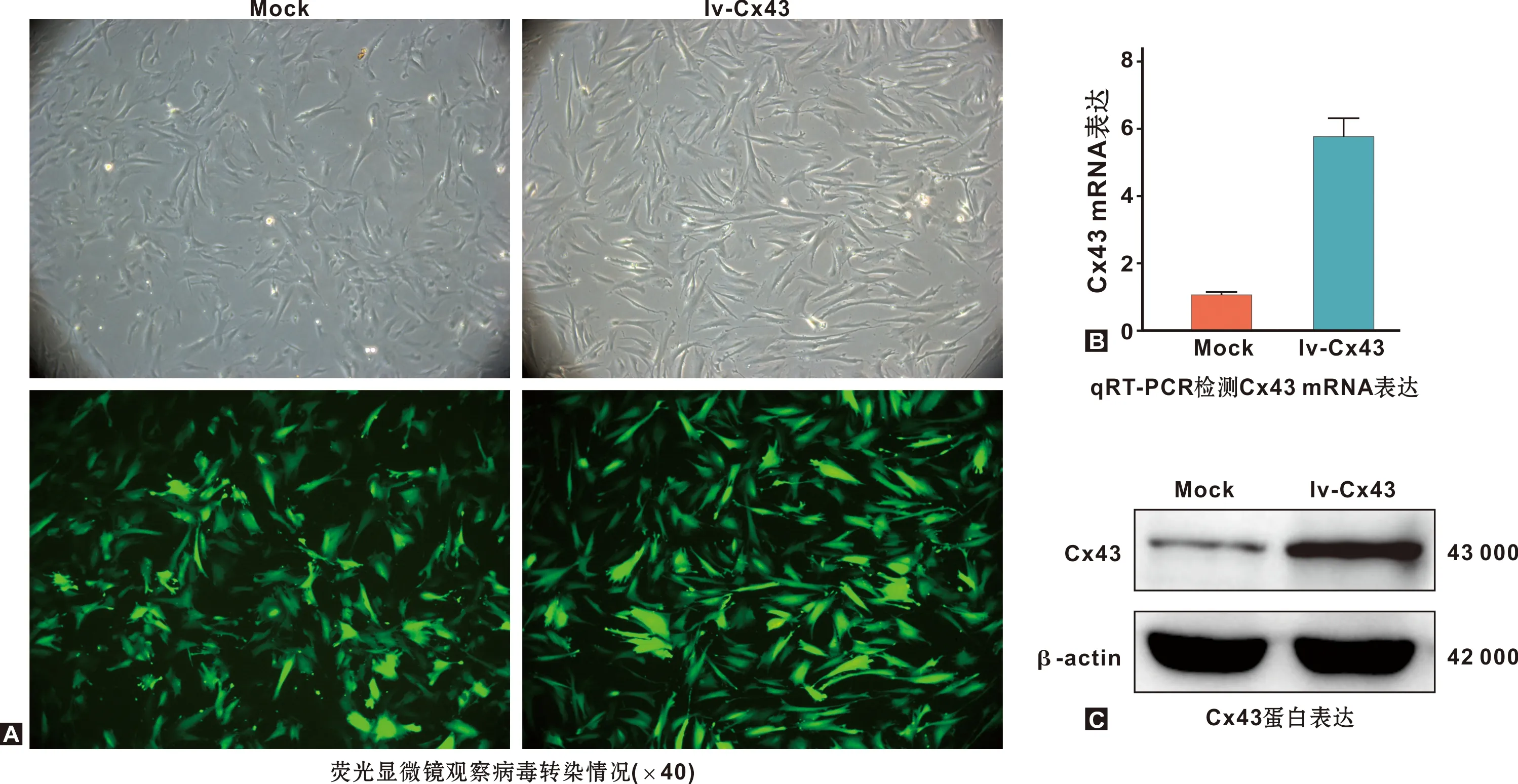
圖1 hDPCs中分別轉染lv-Cx43和mock
2.2 過表達Cx43促進BMP-2誘導的hDPCs成牙本質分化
與mock組相比,BMP-2刺激hDPCs 1 h顯著上調hDPCs內osterix、DSPP、DMP-1、ALP和OCN RNA水平;過表達Cx43明顯增強BMP-2對DSPP等的誘導作用(P<0.05),特別是osterix RNA表達為單獨施加BMP-2刺激的2 倍,但僅過表達Cx43并未顯著上調hDPCs內DSPP 等的表達水平(P>0.05)(圖2A)。同樣,BMP-2明顯上調hDPCs內osterix蛋白水平,過表達Cx43增強hDPCs內BMP-2誘導的osterix蛋白表達。定量分析顯示,其與BMP-2+mock組比存在顯著性差異(P<0.05)(圖2C);與mock組相比,過表達Cx43上調了osterix蛋白表達,但無顯著性差異(圖2B和C)。IF結果顯示,DSPP表達于細胞質內;與mock組相比,BMP-2增強hDPCs內DSPP熒光強度,過表達Cx43促進BMP-2誘導的hDPCs內DSPP表達(圖2D)。結果表明,BMP-2促進DSPP等的表達及hDPCs成牙本質分化;僅過表達Cx43不會上調hDPCs內DSPP等表達水平,但卻增強BMP-2對hDPCs成牙本質分化的誘導作用。
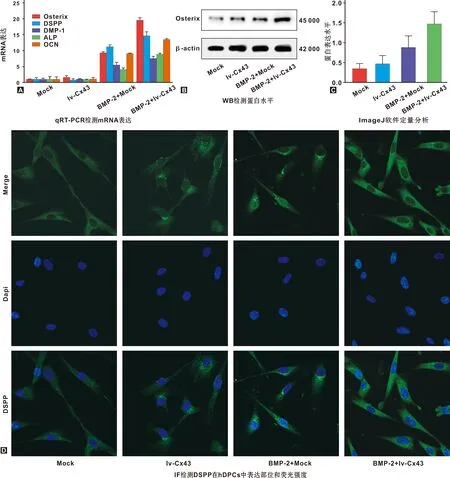
圖2 過表達Cx43促進BMP-2誘導的hDPCs成牙本質分化
2.3 Cx43通過Erk1/2增強BMP-2誘導的hDPCs成牙本質分化
明確Cx43對BMP-2誘導hDPCs成牙本質分化的作用后,進一步探討Cx43發揮功能的途徑。前文已述,BMP-2與Cx43均可通過Erk1/2發揮生物學功能。由此推測:BMP-2與Cx43可能通過Erk1/2信號發生串聯,實現對hDPCs成牙本質分化的促進作用。研究顯示,在hDPCs中,BMP-2激活Smad1/5通路[3]。于是,分別檢測了Smad1/5/9和Erk1/2。結果顯示,BMP-2激活了hDPCs內Smad1/5/9信號;然而,與mock組相比,Erk1/2磷酸化水平無明顯改變(圖3A),提示BMP-2促進hDPCs成牙本質分化可能與Erk1/2無關聯;過表達Cx43顯著上調BMP-2刺激下Erk1/2磷酸化水平(P<0.05),但對Smad1/5/9無明顯促進作用(P>0.05)(圖3A)。結果表明,Cx43促進hDPCs成牙本質分化可能是通過Erk1/2,而與BMP-2介導的Smad1/5/9無關。
為進一步確定Erk1/2的作用,在BMP-2誘導前施加10 μM PD98059(Erk1/2抑制劑)刺激1 h。結果顯示,與BMP-2+lv-Cx43組相比,抑制Erk1/2顯著下調osterix、DSPP、DMP-1、ALP和OCN RNA水平(P<0.05)(圖3B),且明顯減弱osterix蛋白表達(圖3C~D)和DSPP熒光強度(圖3E),表明Cx43通過Erk1/2促進BMP-2誘導hDPCs成牙本質分化。
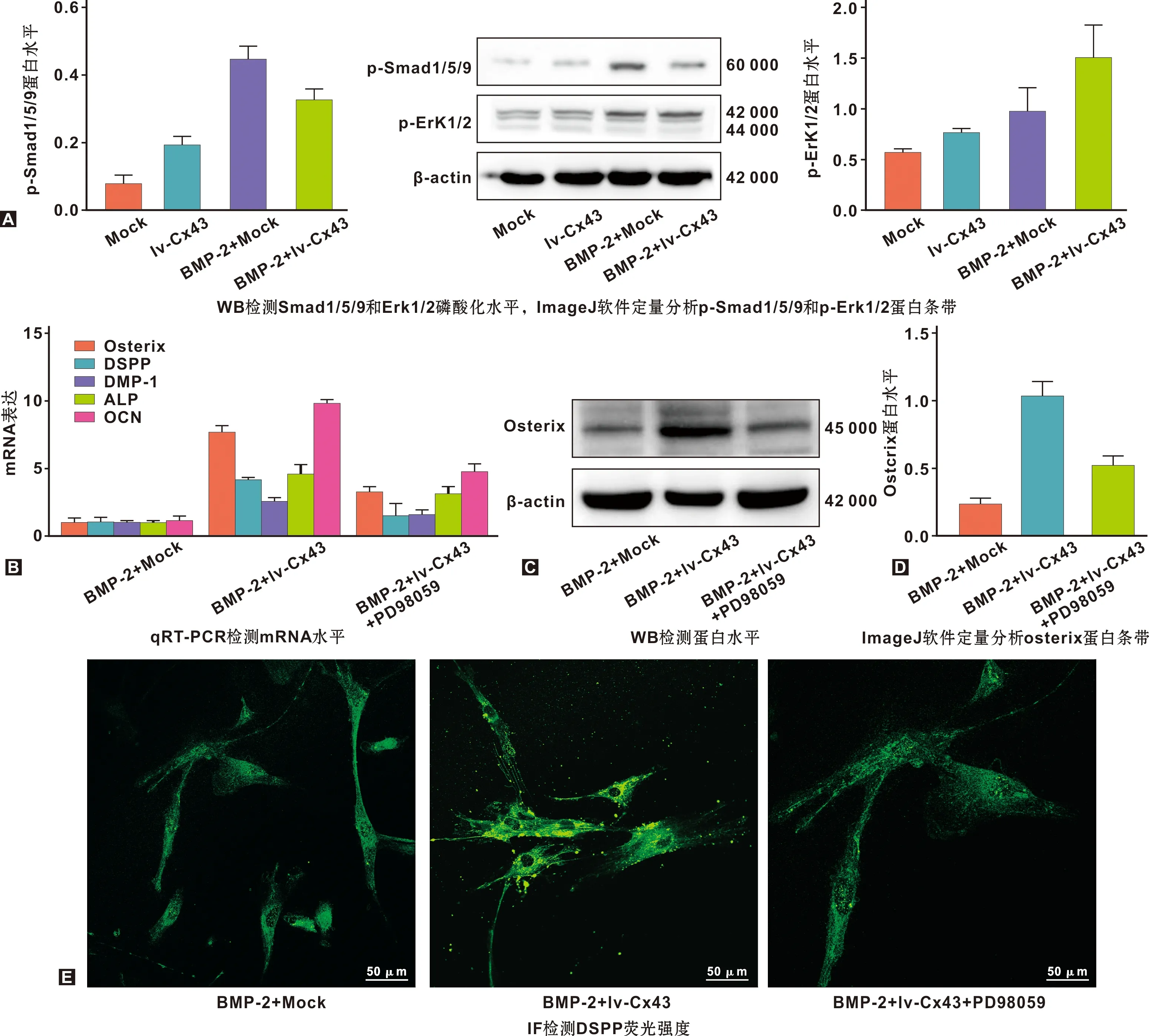
圖3 Cx43通過Erk1/2促進BMP-2誘導hDPCs成牙本質分化
3 討 論
Cx43表達于牙髓中的成牙本質細胞內。牙齒損傷過程中,Cx43表達水平上調[6],表明Cx43參與調控牙齒的損傷后修復,但作用機制不明。本研究顯示,BMP-2激活hDPCs中Smad1/5/9信號,而不是Erk1/2;過表達Cx43不能上調DSPP等表達,但增強BMP-2誘導的hDPCs成牙本質分化,Cx43功能的實現與Erk1/2信號有關。
Runx2和osterix均為重要調節牙本質發生的轉錄因子[10-11]。項目前期研究發現,Runx2促進牙乳頭細胞向成牙本質細胞分化,但抑制成牙本質細胞終末成熟,導致牙本質發生異常[12]。Osterix能夠促進成牙本質細胞增殖和成熟,對牙本質發生不可或缺[13];此外,BMP-2通過osterix誘導成牙本質細胞DSPP表達[14-15]。因此,本研究僅檢測了與BMP-2關系密切的osterix。
本研究發現,僅過表達Cx43未能上調DSPP等的表達,說明過量Cx43對hDPCs成牙本質分化無作用,但過表達Cx43卻能促進BMP-2的誘導作用,提示Cx43功能實現可能是通過間接作用。研究顯示,BMP-2可以通過Smad1/5促進hDPCs成牙本質分化[3]。本研究驗證了前人的結果,并檢測了Cx43對BMP-2介導的Smad1/5/9的影響。結果發現,過表達Cx43并未上調BMP-2介導的Smad1/5/9磷酸化水平,說明Cx43可能是通過其他途徑發揮作用。因此,檢測了Erk1/2信號的活性。結果顯示,過表達Cx43顯著上調了hDPCs中Erk1/2磷酸化水平。隨后,施加了PD98059刺激,發現抑制Erk1/2減弱Cx43上調的osterix和DSPP等表達水平,表明Cx43促進BMP-2誘導的hDPCs成牙本質分化與Erk1/2有一定的關聯。
然而,BMP-2并未上調hDPCs中Erk1/2的磷酸化水平,過表達Cx43又是如何通過Erk1/2促進BMP-2的誘導作用呢?Cx43主要通過形成縫隙通道在細胞間或胞內外傳遞小分子物質而發揮作用。在hDPCs中,BMP-2刺激可能激活了細胞內與Cx43相關的小分子物質,過表達Cx43上調其介導的通道活性,增加了細胞間的通透性,可能促進被BMP-2激活的小分子物質在細胞間傳遞,進而激活Erk1/2信號并促進hDPCs成牙本質分化,而跟BMP-2介導的Smad1/5/9可能不存在必然聯系。因此,Cx43在hDPCs成牙本質分化中的機制還需進一步研究。
總之,本研究發現,BMP-2誘導hDPCs成牙本質分化與Smad1/5/9有關,非Erk1/2;過表達Cx43不能上調DSPP等表達,卻通過Erk1/2促進BMP-2誘導的hDPCs成牙本質分化。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
人大建設(2019年12期)2019-05-21 02:55:32
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
