腌制風干過程中卵形鯧鲹魚肉性質、蛋白質氧化及游離氨基酸的變化
2022-10-09 01:57:56張進偉陳勝軍趙永強吳燕燕王悅齊
食品科學 2022年18期
張進偉,胡 曉,陳勝軍,3,4,*,趙永強,4,吳燕燕,王悅齊,3,潘 創,王 迪
(1.中國水產科學研究院南海水產研究所,農業農村部水產品加工重點實驗室,國家水產品加工技術研發中心,廣東 廣州 510300;2.上海海洋大學食品學院,上海 201306;3.三亞熱帶水產研究院,海南 三亞 572000;4.大連工業大學 海洋食品精深加工關鍵技術省部共建協同創新中心,遼寧 大連 116034)
卵形鯧鲹(),俗稱金鯧、黃臘鯧,其肉質鮮嫩且無肌間刺,富含蛋白質、脂肪、多不飽和脂肪酸等營養物質,是海水養殖經濟魚種。2020年我國卵形鯧鲹養殖產量達到10.1萬 t。在我國南方地區,人們常通過腌制、風干工藝制作腌臘魚、風干魚等,從而提高原料的附加值,延長產品保質期,提高產品風味。蛋白質是水產品的主要成分之一,加工過程中的氧化水解影響產品品質。多肽和游離氨基酸是在內源蛋白酶的作用下降解蛋白質產生,是風味物質的前體物質。游離氨基酸可以與還原糖發生美拉德反應,生成揮發性氣味物質,對產品風味形成具有重要作用。此外,有研究表明肌原纖維蛋白降解會影響魚肉的硬度和彈性。因此,有必要對腌制風干過程中魚肉的蛋白質氧化及游離氨基酸變化規律進行探討。
目前已有一些國內外學者對水產品腌制風干加工過程中蛋白質氧化和游離氨基酸的變化進行報道。Zhang Jinjie等研究發現臘魚在腌制風干過程中游離氨基酸含量顯著上升。吳燕燕等研究發現帶魚在腌干過程中蛋白質在蛋白酶、氨肽酶的作用下發生水解,蛋白水解指數呈上升趨勢;而非蛋白含量在腌制后下降,在風干過程中增加。張會麗等在風干鱸魚過程中發現隨著風干溫度的升高,蛋白水解指數增加,而游離氨基酸總量呈先增加后減少的趨勢。本研究主要對卵形鯧鲹在腌制風干加工過程中魚肉含水率、鹽含量、質構特性、肌原纖維蛋白二級結構、羰基含量、巰基含量、蛋白質水解指數、十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)圖譜和游離氨基酸等進行測定分析,初步揭示卵形鯧鲹魚肉在腌制風干過程中蛋白質氧化及游離氨基酸變化規律,為腌制水產品加工過程中品質控制提供一定理論參考。
1 材料與方法
1.1 材料與試劑
卵形鯧鲹長(31.32±0.88)cm、體質量(537.80±52.38)g,2021年5月購于廣東廣州市新港西路華潤萬家超市,置于-20 ℃冰箱中保存。
甲醇(色譜純)、乙腈(色譜純)、硝酸銀標準滴定溶液(AgNO濃度0.1 mol/L) 上海安譜實驗科技股份有限公司;17 種氨基酸混合標準品(2.5 μmol/mL)、天冬酰胺、谷氨酰胺、瓜氨酸、-正纈氨酸、色氨酸、羥脯氨酸、肌氨酸標準品 西格瑪奧德里奇(上海)貿易有限公司;考馬斯亮藍法蛋白檢測試劑盒、蛋白質羰基檢測試劑盒、總巰基檢測試劑盒 南京建成海浩生物科技有限公司;NuPAGE12% Bis-Tris預制膠英濰捷基(上海)貿易有限公司;苯甲基磺酰氟(phenylmethanesulfonyl fluoride,PMSF)、BeyoBlue?考馬斯亮藍超快染色液、SDS-PAGE蛋白上樣緩沖液(5×)、BeyoColor彩色預染蛋白分子質量標準 上海碧云天生物技術有限公司;三氯乙酸、溴化鉀(均為分析純) 廣州化學試劑廠;實驗用其他試劑均為分析純。
1.2 儀器與設備
RCX-1500-1540熱泵除濕干燥機 佛山市順德區勒流鎮顯高冷凍設備廠;H1850R臺式高速冷凍離心機湘儀離心機儀器有限公司;809 Titrando自動電位滴定儀瑞士萬通公司;T50均質機 德國IKA公司;Kjeltec2300全自動凱式定氮儀 丹麥福斯分析儀器公司;Brookfiled CT3型質構儀 美國博勒飛公司;IRAffinity-1傅里葉變換紅外光譜儀 日本島津公司;Mini Gel Tank PAGE系統 美國賽默飛科技公司;1100液相色譜儀(配有可變波長紫外檢測器) 安捷倫科技(中國)有限公司。
1.3 方法
1.3.1 樣品制備
卵形鯧鲹流水解凍后,剖開魚腹,除去魚鰓、內臟、黑膜等,用流動水清洗魚體內殘血等臟污,室溫下瀝干水分,以備后續腌制使用。稱取魚體質量10%的食鹽,將其均勻涂抹在魚體表面及內部,然后將魚整齊堆放于腌制容器中室溫下腌制2 d,每12 h將魚翻動一次。腌制2 d后將魚取出,除去魚體表面的鹽分,將魚置于熱泵除濕干燥機中干制4 d。參考任中陽等的工藝參數和預實驗設置熱泵干燥機工藝參數:溫度(30±2) ℃、相對濕度(30±5)%。
1.3.2 含水率測定
按照GB 5009.3—2016《食品中水分的測定》中直接干燥法進行測定。
1.3.3 鹽含量測定
按照GB 5009.44—2016《食品中氯化物的測定》中電位滴定法進行測定。
1.3.4 質構特性測定
參考Lerfall等的方法并稍作修改。取魚背部肌肉將其修整為2 cm×2 cm×1 cm(長×寬×高),使用Brookfiled CT3型質構儀及TA 19平底圓柱形探頭進行測定,測定模式為質地多面剖析(TPA)模式,測試速度1 mm/s,觸發點負載5.0 g,下壓距離5 mm,每組實驗重復8 次。
1.3.5 肌原纖維蛋白提取
參考Pazos等的方法并稍作修改。精確稱取2 g絞碎魚肉樣品于50 mL離心管中,加入10 mL預冷卻的勻漿緩沖溶液(pH 7.2,5 mmol/L PMSF和10 mmol/L Tris-HCl緩沖液),冰浴條件下以12 000 r/min均質1 min(均質10 s,間隔10 s,防止過熱蛋白質變性)。均質后在4 ℃、12 000 r/min離心15 min,棄去上清液,向沉淀中加入10 倍體積的鹽溶液(pH 7.2,0.6 mol/L NaCl、5 mmol/L PMSF和10 mmol/L Tris-HCl緩沖液),均質混勻。冰水浴靜置25 min,再以4 ℃、12 000 r/min離心15 min,所得上清液為肌原纖維蛋白溶液,測定肌原纖維蛋白含量,并置于-80 ℃冰箱中保存。
1.3.6 巰基與羰基含量測定
按照蛋白巰基試劑盒與蛋白羰基試劑盒的說明書對提取的肌原纖維蛋白中巰基與羰基含量進行測定。
1.3.7 傅里葉變換紅外光譜測定
肌原纖維蛋白溶液置于-80 ℃冰箱冷凍4 h后取出,經真空冷凍干燥72 h后,得到肌原纖維蛋白凍干樣品以備后續使用。參考李銳等的方法并略作修改,將肌原纖維蛋白凍干樣品與溴化鉀按1∶100的質量比混合,充分研磨,壓制成薄片。使用傅里葉變換紅外光譜全波段掃描(4 000~400 cm),分辨率4.0 cm,掃描次數32 次。使用Peakfit v4.12軟件處理數據,分析蛋白質二級結構及其相對含量。
1.3.8 SDS-PAGE測定
參照Pan Chuang等的方法,使用Mini Gel Tank小型膠電泳槽對12%預制膠進行SDS-PAGE分析。用鹽溶液將肌原纖維蛋白溶液稀釋到1 mg/mL,取30 μL 1 mg/mL肌原纖維蛋白溶液與10 μL上樣緩沖液混合,沸水浴5 min,上樣量10 μL,電泳電壓設置為120 V。電泳結束時后取下凝膠,使用考馬斯亮藍超快染色液染色,脫色至背景無色,用凝膠成像系統掃描并分析條帶。
1.3.9 總氮與非蛋白氮含量測定
總氮測定:參考GB 5009.5—2016《食品中蛋白質的測定》中自動凱式定氮儀法進行測定。
非蛋白氮測定:參考吳燕燕等方法并稍作修改。準確稱取5 g魚肉(精確至0.001 g),加入5 倍體積預冷的質量分數10%的三氯乙酸溶液 ,22 000 r/min高速勻漿(3×10 s),4 ℃條件下放置12 h后取出,4 000 r/min離心5 min。用中速濾紙過濾上清液,用少量質量分數10%三氯乙酸洗滌沉淀,過濾后合并上清液并定容至100 mL。取10 mL濾液消化,用自動凱式定氮儀測定。
按下式計算蛋白水解指數:
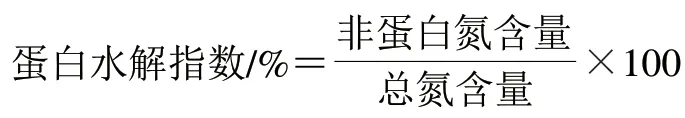
1.3.10 游離氨基酸含量測定
1.3.10.1 樣品前處理
稱取1 g絞碎魚肉樣品于10 mL離心管中,與5 mL 0.01 mol/L鹽酸溶液混勻,沸水浴30 min后,10 000 r/min離心10 min,取上清液。向沉淀中加入2 mL 0.01 mol/L鹽酸溶液,懸浮超聲5 min,10 000 r/min離心10 min,合并上清液并定容至10 mL,過0.22 μm濾膜后上機測定。
1.3.10.2 液相色譜條件
ZORBAX Eclipse AAA氨基酸分析色譜柱(4.6 mm×150 mm,3.5 μm);流速:1 mL/min;流動相:A為40 mmol/L磷酸二氫鈉(pH 7.8),B為乙腈-甲醇-水(45∶45∶10,/);梯度洗脫:0~1 min,100% A、0% B;1~23 min,43% A、57%B;23~34 min,0% A、100% B;34~41 min,100%A、0% B;紫外檢測器波長:338 nm(0~19 min),266 nm(19.01~25 min)。
1.4 數據處理
每個指標設置3 個平行。采用Excel進行數據處理和作圖,采用SPSS Statistics 23.0軟件進行統計分析,Duncan法進行差異顯著性分析。<0.05,差異顯著。
2 結果與分析
2.1 腌制風干對卵形鯧鲹含水率和鹽含量的影響
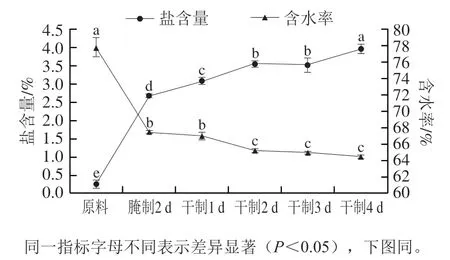
圖1 卵形鯧鲹在腌制風干過程中鹽含量和含水率的變化Fig. 1 Change in salt content and water content of T. ovatus during curing and air-drying
由圖1可知,在腌制、風干過程中卵形鯧鲹的鹽含量呈上升趨勢,其中腌制2 d后卵形鯧鲹的鹽含量急劇增加(<0.05);而在此過程中含水率呈下降趨勢,分別在干制1、3、4 d后含水率變化差異不顯著(>0.05)。腌制是滲透擴散的過程,干腌是將魚體表面及內部均勻涂抹食鹽,使其內外形成高滲透壓,從而使鹽含量顯著增加,水分含量下降。在風干過程中鹽含量逐漸增加,但干制2 d與3 d后卵形鯧鲹鹽含量差異不顯著(>0.05),這可能是由于卵形鯧鲹表皮水分快速脫除,內部水分緩慢遷移,形成硬殼,使含水率變化緩慢,鹽含量變化不顯著。此外,在干制4 d后,卵形鯧鲹的鹽含量達到最高值,為(3.96±0.13)%。
2.2 腌制風干對卵形鯧鲹質構特性的影響
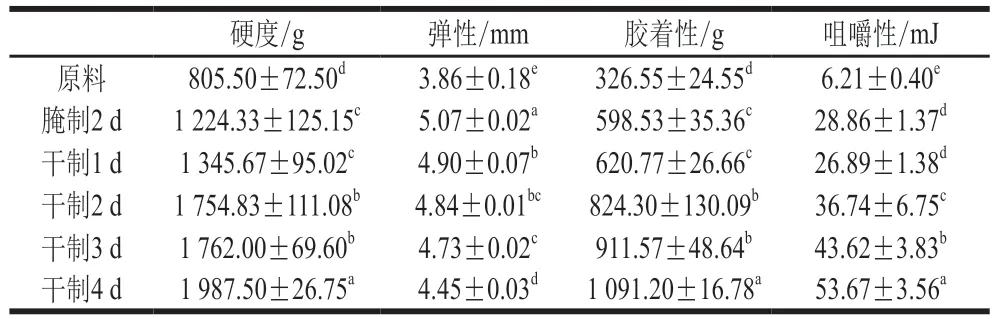
表1 卵形鯧鲹在腌制風干過程中質構特性的變化Table 1 Change in texture characteristics of T. ovatus during curing and air-drying processes
質構特性主要由硬度、彈性、膠著性、咀嚼性等性質組成,可以反映魚肉的組織狀態,是衡量其食用價值的重要指標。魚肉質構特性的變化與其水分、pH值、細胞外基質(膠原蛋白)結構、蛋白質水解等因素有關。由表1可知,隨著腌制風干的進行,卵形鯧鲹背部肌肉硬度增加,造成該現象的原因是魚肉經腌制、風干后水分含量減少,鹽含量升高,肌纖維收縮,肉質緊密,從而使魚肉硬度增加。Yang Wenxian等研究發現草魚在腌制2 d后,魚肉硬度顯著增加,與本研究結果相似。由表1可知,魚肉在腌制2 d后彈性增加,并隨著風干的進行魚肉彈性降低。通常魚肉的鹽含量較低時,其彈性會有所增加,而在風干過程中魚肉的鹽含量明顯增加,水分含量減少,彈性隨含水率降低而降低,同時隨著魚肉中內部水分向外擴散,肌纖維直徑及其橫截面積減小,肌纖維收縮,導致魚肉彈性下降。由表1可知,在腌制風干加工過程中,魚肉的膠著性呈上升趨勢。有研究表明脂肪含量對膠著性具有一定影響,而在腌制風干過程中魚肉水分含量減少,脂肪相對含量增加,所以可能導致魚肉膠著性的變化。由表1可知,卵形鯧鲹魚肉咀嚼性整體呈上升趨勢;其中,在腌制2 d后,魚肉咀嚼性顯著增加,而在風干1 d后略有下降但并不顯著,之后呈顯著增加趨勢(<0.05)。
2.3 腌制風干對卵形鯧鲹肌原纖維蛋白含量的影響
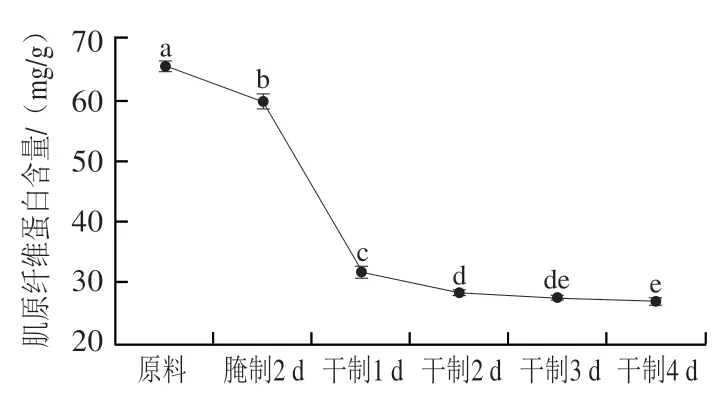
圖2 卵形鯧鲹在腌制風干過程中肌原纖維蛋白含量的變化Fig. 2 Change in myofibrillar protein content of T. ovatus during curing and air-drying
肌原纖維蛋白是魚肉中重要蛋白質,約占總蛋白含量的50%~70%,決定魚肉質構、熱凝膠等加工特性,影響產品品質。由圖2可知,卵形鯧鲹在腌制風干過程中肌原纖維蛋白含量呈下降趨勢;在腌制2 d后、干制1 d后和2 d后,肌原纖維蛋白含量顯著降低(<0.05),可能是加工過程中肌原纖維蛋白的溶出或水解產生多肽和游離氨基酸所致。
2.4 腌制風干對卵形鯧鲹蛋白質羰基含量的影響

圖3 卵形鯧鲹在腌制風干過程中羰基含量的變化Fig. 3 Change in carbonyl content of T. ovatus during curing and air-drying
蛋白質羰基主要由賴氨酸、脯氨酸、精氨酸等直接氧化生成,是評價蛋白氧化的重要指標。由圖3可知,隨著腌制風干的進行,羰基含量明顯增加,蛋白質氧化程度增加;卵形鯧鲹原料的蛋白質羰基含量為1.37 nmol/mg,腌制2 d后蛋白質羰基含量顯著增加(<0.05)至1.94 nmol/mg,風干過程中蛋白質羰基含量持續增加,而干制1 d和2 d后蛋白質羰基含量差異不顯著(>0.05)。趙亞南等研究發現在一定范圍內隨著氯化鈉添加量的增加,蛋白質羰基含量增加。本實驗中,卵形鯧鲹在腌制過程中蛋白質羰基含量的增加,可能是氯化鈉滲入魚肉內部,增強Fe的活性,Fe進一步影響魚肉蛋白氧化所致。此外,卵形鯧鲹魚肉中富含脂肪,在腌制風干過程中脂肪發生氧化降解,而脂肪氧化過程中產生自由基,自由基攻擊蛋白質氨基酸側鏈、肽鏈骨架,導致氨基酸側鏈暴露,發生蛋白質聚合、交聯等反應,可能使蛋白質羰基含量增加。
2.5 腌制風干對卵形鯧鲹巰基含量的影響
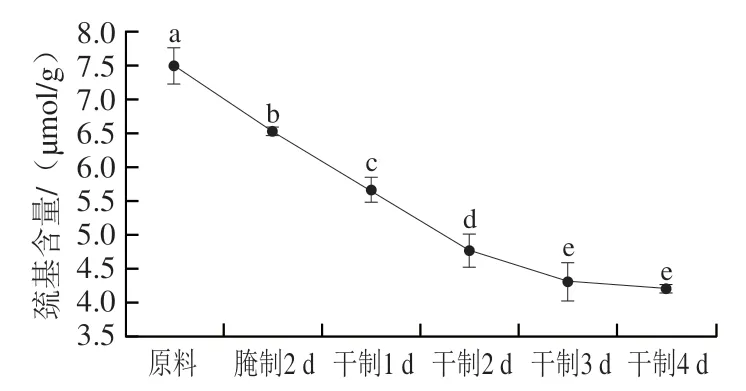
圖4 卵形鯧鲹在腌制風干過程中巰基含量的變化Fig. 4 Change in sulfhydryl content of T. ovatus during curing and air-drying
活性巰基主要存在于半胱氨酸,是蛋白質結構和生物體內某些氧化還原反應的重要基團。在同一肽鏈或不同肽鏈間的半胱氨酸殘基的巰基脫氫氧化形成二硫鍵,二硫鍵含量上升,蛋白質變性,所以巰基含量能夠表征蛋白質氧化程度。由圖4可知,巰基含量的變化趨勢與羰基整體相反,隨著腌制風干的進行,巰基含量顯著下降(<0.05),在干制4 d后巰基含量差異不顯著(>0.05)。造成該現象的原因是腌制、風干工藝使魚肉蛋白質結構被破壞,導致埋藏在蛋白質內部的巰基暴露出來,產生二硫鍵,使總巰基含量下降。葉路漫研究發現魚肉總巰基含量隨著腌制、風干工藝時間推移而下降,與本研究結果相近。
2.6 腌制風干對卵形鯧鲹肌原纖維蛋白二級結構的影響
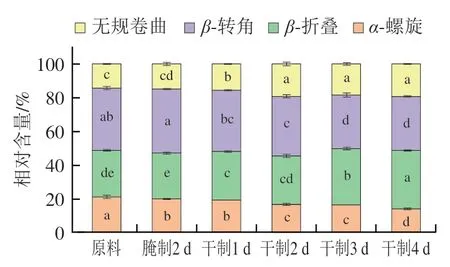
圖5 卵形鯧鲹在腌制風干過程中肌原纖維蛋白二級結構組成變化Fig. 5 Change in secondary structure composition of myofibrillar protein from T. ovatus during curing and air-drying
蛋白質二級結構在紅外光譜區有若干吸收帶,其中酰胺I帶(1 600~1 700 cm)是二級結構的關鍵吸收帶,主要由羰基C=O伸縮振動引起。蛋白質的二級結構主要包括-折疊(1 600~1 639 cm)、無規卷曲(1 640~1 650 cm)、-螺旋(1 651~1 660 cm)、-轉角(1 661~1 700 cm)。-螺旋具有高度穩定性,主要由多肽鏈上羰基和氨基之間的氫鍵維持,是蛋白質中最主要最穩定的結構,而無規卷曲和-結構的緊密程度和構象穩定性相較于-螺旋較差。
由圖5可知,隨著腌制風干的進行,肌原纖維蛋白二級結構中-螺旋含量減少,而無規卷曲含量增加。有研究表明氯化鈉可能對肌原纖維蛋白的二級結構具有一定破壞作用。可能是因為卵形鯧鲹在腌制風干過程中食鹽滲透進入魚肉內部,肌原纖維蛋白質結構被破壞,羰基含量上升,從而使-螺旋含量減少,無規卷曲含量增加。此外,曹錦軒等研究發現臘肉肌原纖維蛋白在加工過程中-螺旋含量減少,與本研究結果類似。通常在加熱條件下,-螺旋含量降低代表蛋白質分子展開程度增加,-折疊含量增加代表蛋白質分子間聚集程度增加。由圖5可知,卵形鯧鲹在加工過程中-轉角含量減少,而-折疊含量增加,蛋白質分子間聚集程度增加。
2.7 卵形鯧鲹肌原纖維蛋白的SDS-PAGE分析
在肉質品加工過程中,肌原纖維蛋白是主要的蛋白質,與肉制品質構特性密切相關。如圖6所示,在加工過程中,卵形鯧鲹肌原纖維蛋白的SDS-PAGE條帶發生明顯變化,其中主要的蛋白質有肌球蛋白重鏈(myosin heavy chain,MHC)、肌動蛋白、原肌球蛋白等。蛋白分子降解主要表現為分子質量較高的條帶出現模糊、弱化、擴展,較低分子質量區域出現新條帶或條帶顏色加深。由圖6可知,MHC條帶和肌動蛋白發生明顯變化;其中MHC條帶在腌制2 d后變化較小,可能是腌制時肌原纖維蛋白發生交聯或聚集;而風干后MHC條帶消失,表明大分子質量的蛋白質在內源蛋白酶作用下發生降解。肌動蛋白條帶在腌制2 d后變細,造成此現象的原因可能是氯化鈉濃度升高,導致肌動蛋白溶出;而干制1 d后肌動蛋白條帶又變粗,可能是由于在風干過程中大分子質量蛋白降解形成較小分子質量蛋白所致。
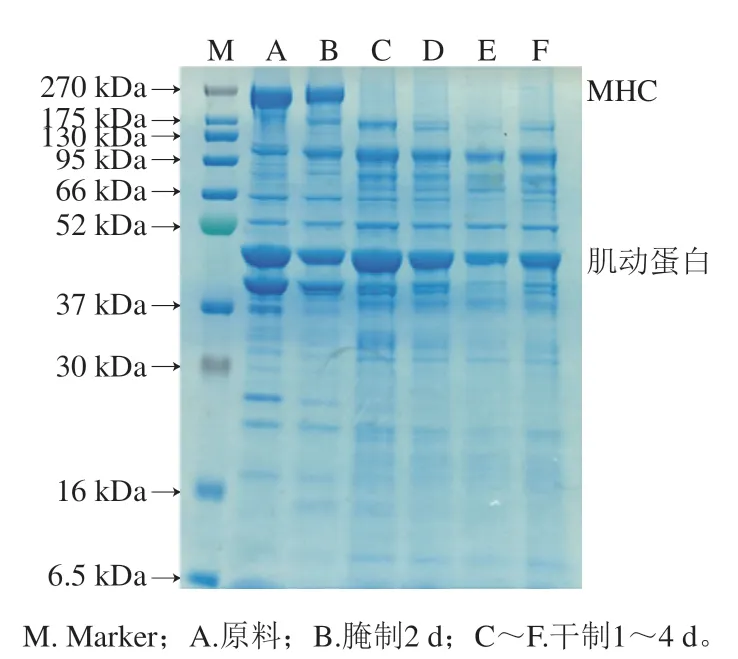
圖6 卵形鯧鲹在腌制風干中肌原纖維蛋白的SDS-PAGE圖譜Fig. 6 SDS-PAGE pattern of myofibrillar protein from T. ovatus during curing and air-drying
2.8 腌制風干對卵形鯧鲹總氮與非蛋白氮含量的影響
非蛋白氮是蛋白質和多肽在內源蛋白酶和氨肽酶的作用下降解產生。如圖7所示,在加工過程中,卵形鯧鲹總氮含量在腌制2 d后顯著下降(<0.05),可能是在腌制過程中水溶性蛋白質溶出所致,這與張會麗的研究結果一致;而在風干過程中總氮含量整體呈緩慢下降的趨勢。此外,在腌制加工后非蛋白氮含量下降,造成該現象的原因是水溶性含氮物質溶出;而在風干過程中非蛋白氮含量明顯增加,主要是在魚肉內源蛋白酶的作用下,蛋白質水解為多肽、游離氨基酸等造成的。史培磊等研究發現風鵝加工過程中非蛋白氮含量在腌制后下降,而在風干過程中先上升后下降,這與本研究結果有所不同,可能是風干工藝時間不同所致。
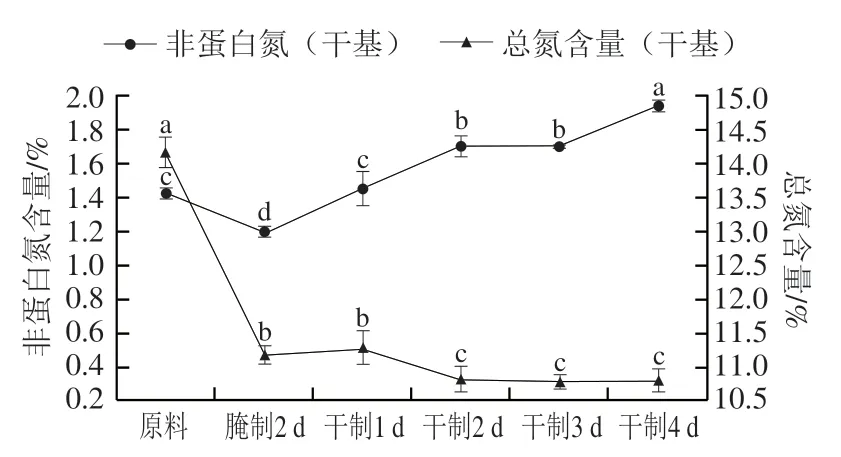
圖7 卵形鯧鲹在腌制風干中總氮、非蛋白氮的變化Fig. 7 Change in total nitrogen and non-protein nitrogen contents of T. ovatus during curing and air-drying
蛋白質水解主要通過內源蛋白酶、微生物酶等作用產生降解,蛋白質的水解可以增加肉質的柔軟性,主要與肌原纖維的破碎有關,同時產生多肽、游離氨基酸等物質,對風味形成有間接作用。由圖8可知,蛋白水解指數在腌制2 d后變化較小,差異不顯著(>0.05);而在風干過程中蛋白水解指數明顯升高,原因可能是在風干過程中酶活性提高,魚肉蛋白質降解為多肽、游離氨基酸等,同時含氮物質溶出較少,從而使蛋白水解指數增加。
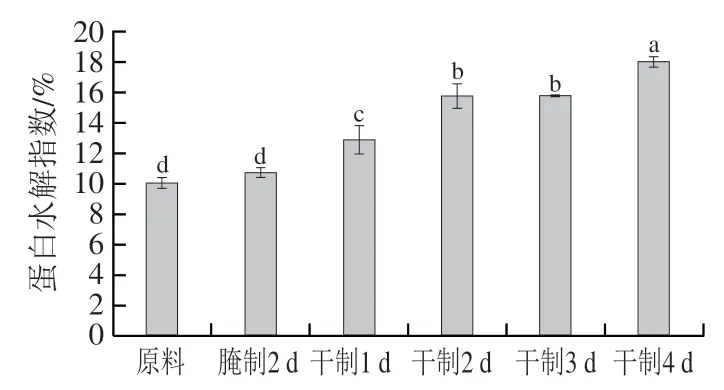
圖8 卵形鯧鲹在腌制風干中蛋白水解指數的變化Fig. 8 Change in proteolysis index of T. ovatus during curing and air-drying
2.9 腌制風干對卵形鯧鲹游離氨基酸含量的影響

表2 卵形鯧鲹在腌制風干中游離氨基酸的變化Table 2 Change in free amino acids of T. ovatus during curing and air-drying mg/100 g
游離氨基酸通過蛋白質降解產生,是水產品中重要的風味物質,其呈味特征與含量、閾值有關。其中丙氨酸、甘氨酸、蘇氨酸、絲氨酸、脯氨酸呈現甜味;天冬氨酸、谷氨酸呈現鮮味。如表2所示,原料中游離氨基酸總量為380.03 mg/100 g,其中丙氨酸、羥脯氨酸、甘氨酸、谷氨酸含量較高;而在干制4 d后游離氨基酸總量為1 596.07 mg/100 g,其中丙氨酸、甘氨酸、谷氨酸、亮氨酸、賴氨酸含量較高,約占總游離氨基酸含量的50.67%。游離氨基酸總量在腌制2 d后顯著增加(<0.05),這與吳燕燕等研究結果相近;在干制前3 d游離氨基酸總量仍顯著增加(<0.05),表明在風干過程中蛋白質在內源酶的作用下水解產生游離氨基酸;干制4 d后游離氨基酸總量下降,可能是由于部分氨基酸與還原糖發生美拉德反應以及在微生物作用下生成風味物質所致。鮮味氨基酸和甜味氨基酸在加工過程中含量總體呈上升趨勢,有利于魚肉形成良好風味,而在干制4 d后其含量略有下降。
3 結 論
在腌制風干過程中,卵形鯧鲹魚肉的鹽含量整體呈上升趨勢,同時其質構特性發生變化:硬度、膠著性、咀嚼性整體呈上升趨勢,而彈性為先上升后下降。在加工過程中,魚肉蛋白質發生氧化反應,羰基含量增加,而巰基含量下降;肌原纖維蛋白二級結構中-螺旋含量下降,而無規卷曲含量增加;MHC條帶和肌動蛋白發生明顯變化:MHC條帶變淺并在風干后消失,肌原纖維蛋白產生降解;蛋白質水解指數增加,非蛋白氮含量在腌制后下降而風干過程中增加;游離氨基酸總量在干制1 d后達到最高,鮮味氨基酸和甜味氨基酸含量總體呈上升趨勢,有利于提高產品風味。
