四川涼山苦蕎種子攜帶細菌多樣性
2022-10-13 11:50:02白凱紅阿別小兵李健強羅來鑫
植物保護 2022年5期
白凱紅, 阿別小兵, 陳 星, 蔣 娜, 李健強, 羅來鑫
(中國農業大學植物病理學系, 農業農村部有害生物監測與綠色管理重點實驗室, 種子病害檢驗與防控北京市重點實驗室, 北京 100193)
苦蕎Fagopyrumtataricum是蓼科Polygonaceae蕎麥屬Fagopyrum的一年生作物,其營養豐富,含有豐富的蛋白質、纖維素、碳水化合物、微量元素等[1-2],還可以提高人體免疫力[3-4]、抗衰老[5]等,享有“五谷之王”的美譽。四川省涼山地區是我國蕎麥起源中心之一,擁有全國乃至全世界最豐富的苦蕎資源,也是種類最多樣、種植最廣泛、分布最集中的地區[6-8]。作為當地的主要糧食之一,涼山州的17個縣均有苦蕎種植,其中越西、昭覺、喜德、冕寧、美姑等縣是苦蕎的主產區[9-10]。涼山地區特殊的地理環境與獨特的氣候有利于蕎麥的產量與品質,全州常年種植面積上百萬畝,約占全國總種植面積的一半以上[6, 11]。
真菌、細菌、病毒、線蟲等病原物均可引起蕎麥病害[12],其中真菌病害報道較多,而細菌性病害報道卻很少,僅見丁香假單胞菌Pseudomonassyringae和銅綠假單胞菌P.aeruginosa[13-14]引起的細菌性葉斑病,而關于蕎麥種子攜帶細菌的報道更少。種子上攜帶的病原菌是田間病害發生的重要初侵染來源,種子攜帶的有益微生物也會對作物生長起到促進作用。加強對種子攜帶微生物的檢測,有利于明確種子健康狀況,挖掘有益微生物群體。種子帶菌檢測的技術主要有直接檢測、洗滌檢測、瓊脂培養檢測、免疫學檢測、分子生物學檢測等,近年來高通量測序技術也陸續應用在種子帶菌檢測中,但在蕎麥種子健康檢測中還未見報道。
本研究運用傳統的分離培養和基于16S rRNA基因的擴增子測序方法,對涼山主要產區的苦蕎種子攜帶的細菌進行了檢測,旨在明確涼山州苦蕎種子攜帶細菌的多樣性,并為種子健康檢測提供新的技術手段和方向。
1 材料與方法
1.1 材料
蕎麥種子樣品:采自四川省涼山州越西縣、昭覺縣、喜德縣、冕寧縣、普格縣、美姑縣和鹽源縣不同鄉村的46份苦蕎種子(表1)用于種子攜帶細菌的檢測,其中選取不同地點的13份種子用于16S rRNA基因擴增子測序。

表1 來自四川省涼山苦蕎種子樣品信息1)

續表1 Table 1(Continued)
1.2 試劑與儀器
LBA培養基:5 g/L酵母浸粉、5 g/L氯化鈉、10 g/L胰蛋白胨、16 g/L瓊脂。
供試試劑:1% NaClO;AxyPrepDNA凝膠回收試劑盒,AXYGEN;Omega-soil DNA Kit,Omega Bio-Tek;TruSeqTM DNA Sample Prep Kit,TransGen。
供試儀器:K30型金屬浴,干式恒溫器,Alphamager HP凝膠成像儀,H1650R離心機(cence?湘儀),ABSON MiFly-6小型離心機(合肥艾本森科學儀器有限公司),球磨儀(Retsch MM400),MyCycler580BR型PCR儀(Bio-Rad),ABI GeneAmp?9700型PCR儀(ABI),DYY-12型電泳儀與DYY-6C電泳儀(北京市六一儀器廠),粉碎研磨儀TL-48R(上海萬柏生物科技有限公司),NanoDrop 2000(Thermo Scientific),微型熒光計TBS380(TurnerBioSystems),MISEQ測序儀Illumina Miseq(Illumina)。
1.3 分離培養結合16S rRNA基因序列鑒定
分離培養:從每一份供試的苦蕎種子中隨機選取100粒,放入滅菌的50 mL錐形瓶中,加入10 mL無菌水,置于搖床中振蕩培養30 min(120 r/min),之后吸取2 mL培養液,用無菌水進行10倍梯度稀釋,稀釋液均勻涂布于LBA培養基上,將培養皿置于28℃培養箱中黑暗培養,2 d后開始觀察、記錄、計算分離頻率(分離頻率=某分離物總數/分離物的總數×100%),并進行細菌的純化分離培養。
形態學鑒定:觀察菌落顏色、大小、形狀、邊緣特征、有無分泌物等。
分子生物學鑒定:挑取1個單菌落置于10 μL無菌水中,在金屬浴100°C加熱10 min破壁獲得總DNA。利用細菌通用引物27F/1492R(5′-AGAGTTTGATCCTGGCTCAG-3′/5′-GGTTACCTTGTTACGACTT-3′)[15]擴增16S rRNA基因序列。PCR體系(25 μL)為:16.89 μL ddH2O、2.5 μL 10×rTaqDNA 聚合酶緩沖液、2.4 μL dNTPs(2.5 mmol/L)、1 μL 上游引物(10 μmol/L)、1 μL 下游引物(10 μmol/L)、0.25 μLrTaqDNA 聚合酶(2.5 U/uL)、1 μL DNA模板。PCR反應程序為:95℃預變性10 min;95℃變性30 s,55℃退火30 s,72℃延伸1 min 30 s,35個循環;72℃延伸10 min。最后將符合目標條帶的樣品進行測序(北京博邁德科技發展有限公司),并將所得序列在NCBI(https:∥blast.ncbi.nlm.nih.gov/Blast.cgi)數據庫中進行同源性比對。
1.4 基于16S rRNA基因擴增子測序的多樣性分析
樣本預處理:在50 mL離心管里加入種子至10 mL刻度線,再加入10 mL滅菌水,渦旋振蕩5 min,種子洗脫液轉移至新管中備用。
DNA 提取、文庫構建、數據分析:利用Omega-soil DNA Kit對基因組DNA進行抽提,利用擴增16S rRNA V3-V4區域通用引物385F/806R(5′-ACTCCTACGGGAGGCAGCAG-3′/5′-GGACTA-
CHVGGGTWTCTAAT-3′)進行PCR擴增,以檢測是否滿足后續擴增子測序要求。滿足要求后,通過文庫構建,進行Miseq測序,對數據進行質控過濾處理以及去嵌合體后獲得最終的有效數據(effective tag),之后進行物種注釋和分析,包括OTU(operational taxonomic units)分析、Alpha多樣性分析、Beta多樣性分析。
2 結果與分析
2.1 分離培養結合16S rRNA基因序列鑒定苦蕎種子攜帶細菌
對46份苦蕎種子攜帶的細菌進行分離培養,共獲得618株細菌分離物。經形態學和分子生物學鑒定,獲得的細菌分離物隸屬于10個屬,分別為賴氨酸芽胞桿菌屬Lysinibacillus、節桿菌屬Arthrobacter、芽胞桿菌屬Bacillus、假單胞菌屬Pseudomonas、微桿菌屬Microbacterium、泛菌屬Pantoea、短小桿菌屬Curtobacterium、鞘氨醇單胞菌屬Sphingomonas、類芽胞桿菌屬Paenibacillus、根瘤菌屬Rhizobium,具體分離物的菌落形態如圖1所示,上述細菌的16S rRNA基因序列與NCBI上已知對應屬的細菌序列的比對結果相似性均在97%以上。通過對苦蕎種子細菌分離物的分離頻率計算與分析,泛菌屬的分離頻率最高,為52.5%,短小桿菌屬、假單胞菌屬、微桿菌屬分別為20.7%、16.5%、5.5%,其他菌群占比為4.8%(圖2)。
2.2 基于16S rRNA基因擴增子測序多樣性分析的苦蕎種子攜帶細菌的檢測
2.2.1OTU分析
對選取的13份苦蕎種子攜帶的細菌進行高通量測序,基于16S rRNA基因擴增子測序獲得的細菌被劃分為19個門(phylum)、40個綱(class)、81個目(order)、151個科(family)、279個屬(genus)、430個種(species),共624個OTUs。

圖1 來自于苦蕎種子的細菌分離物在LBA培養基上的菌落形態Fig.1 Colony morphology of bacterial isolates from tartary buckwheat seeds on LBA medium

圖2 基于分離培養法對供試苦蕎種子攜帶細菌的優勢菌群分析結果Fig.2 Analysis of dominant microflora from tartary buckwheat seeds based on agar plating
2.2.2Alpha多樣性分析
Alpha多樣性分析結果表明(表2),13份苦蕎種子樣品文庫的覆蓋率(coverage)均在99%以上,說明測序深度高,樣本中序列均能被測出,能反映種子真實的帶菌情況。Alpha多樣性分析中,ACE、Chao、Sobs的數值越大,表明菌群中所含有的OTU數目越多,菌群的豐富度(community richness)越大;另外,Shannon的值越大、Simpson的值越小,說明菌群的多樣性(community diversity)越高。表2的結果說明,來自冕寧縣的樣品MNCQ-4的ACE(497.85)、Chao(501.70)、Sobs(483)、Shannon(4.304 8)的數值最大,Simpson(0.043 6)的值最小,說明MNCQ-4菌群的豐富度與多樣性在13份種子樣品中最大。而來自昭覺縣的樣品ZJ-5的ACE(162.22)、Chao(130.40)、Sobs(107)的數值最小,Simpson(0.272 7)的值最大,表明ZJ-5的豐富度與多樣性最小。其余11份樣品的細菌菌群豐富度與多樣性介于MNCQ-4和 ZJ-5之間。
對苦蕎種子攜帶細菌的物種組成進行分析(圖3),結果表明,種帶細菌主要有泛菌屬、甲基桿菌屬Methylobacterium、鞘氨醇單胞菌、馬賽菌屬Massilia、薄層菌屬Hymenobacter、假單胞菌屬、藍細菌屬Cyanobacteria、短小桿菌屬、伯克氏菌屬-副伯克氏菌屬Burkholderia-Paraburkholderia、葡萄球菌屬Staphylococcus、腸桿菌屬Enterobacter、貪噬菌屬Variovorax,其中泛菌屬、甲基桿菌屬、鞘氨醇單胞菌屬、馬賽菌屬、薄層菌屬、假單胞菌屬在13份苦蕎種子樣品中均能被大量檢測到。泛菌屬所占比率最高,為最主要的優勢菌群。
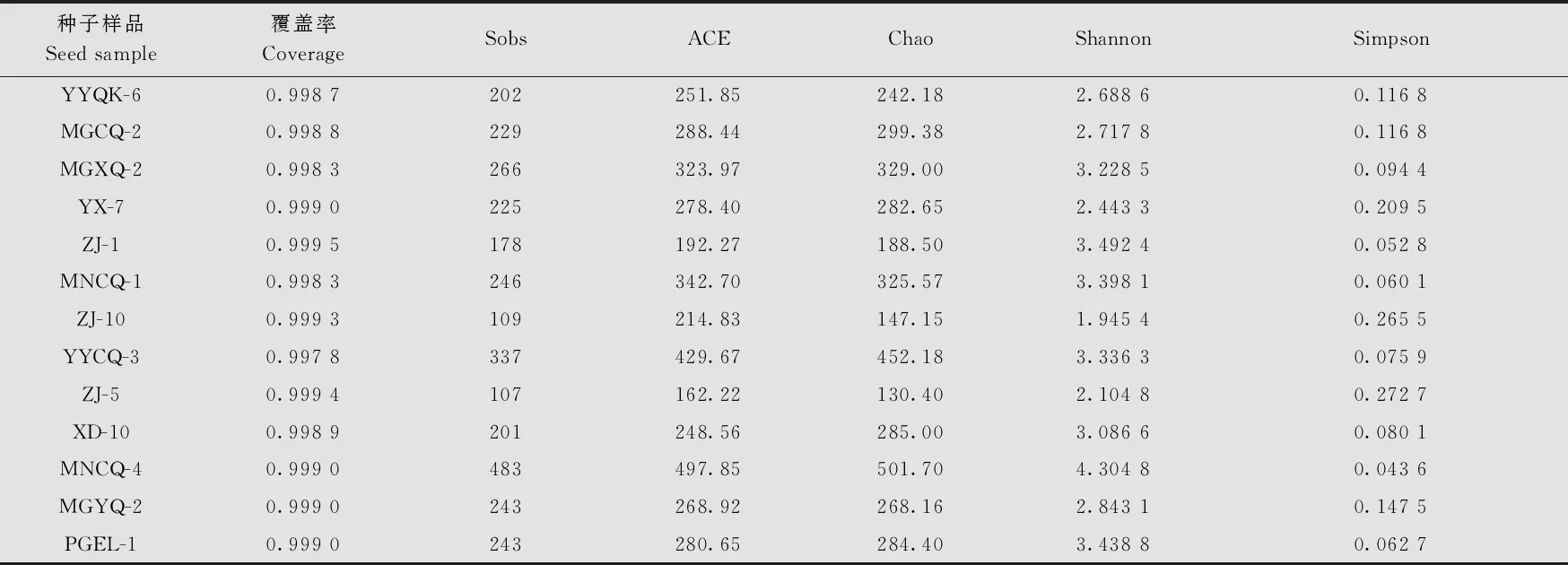
表2 基于16S rRNA基因擴增子測序對供試的13份苦蕎種子攜帶細菌的Alpha多樣性的評估結果統計

圖3 基于16S rRNA擴增子測序對供試的13份苦蕎種子攜帶細菌的物種組成分析結果Fig.3 Community analysis of 13 tartary buckwheat seeds carrying bacteria by 16S rRNA amplicon sequencing
2.2.3Beta多樣性分析
2.2.3.1樣本層級聚類分析
13份苦蕎種子攜帶的細菌聚類分析結果如圖4所示,來自同一個縣的樣品有的能聚在一支, 有的卻相距很遠。例如,來自昭覺縣的樣品ZJ-10與ZJ-5聚在一起,而另一個采自昭覺縣的樣品ZJ-1卻與來自冕寧縣樣品MNCQ-1聚在一支;來自美姑縣的樣品MGXQ-2與MGYQ-2聚在一支,而采自美姑縣的樣品MGCQ-2卻與來自鹽源縣的YYQK-6聚在一支。其中菌群多樣性最相近的兩個樣品為來自鹽源縣的YYCQ-3與來自普格縣的PGEL-1,來自冕寧縣的MNCQ-4與其他樣品的差異性最大,且與同是冕寧縣的MNCQ-1差異性也很大。

圖4 基于16S rRNA擴增子測序對供試的13份苦蕎種子攜帶細菌的樣本層級聚類樹分析結果Fig.4 Sample hierarchy clustering tree analysis of 13 tartary buckwheat seeds carrying bacteria by 16S rRNA amplicon sequencing
2.2.3.2PCA分析
對苦蕎種子攜帶細菌進行PCA分析,結果表明(圖5),來自同一個地區的種子樣品菌群多樣性的差異性小,來自不同地區的種子樣品差異性大,但其中來自冕寧縣(MN)的種子與來自喜德縣(XD)的種子攜帶細菌多樣性相似,且冕寧縣(MN)和喜德縣(XD)種子與普格縣(PG)種子、鹽源縣(YY)種子相似性大。在13份種子樣品中,來自越西縣(YX)種子樣品與來自普格縣(PG)種子樣品的菌群多樣性的差異性最大。

圖5 基于16S rRNA擴增子測序對供試的13份苦蕎種子攜帶細菌的PCA分析結果Fig.5 PCA analysis of 13 tartary buckwheat seeds carrying bacteria by 16S rRNA amplicon sequencing
3 結論與討論
本研究通過分離培養法從46份供試苦蕎種子中共鑒定到10個屬的細菌,其中泛菌屬為苦蕎種子攜帶的優勢細菌菌群。泛菌屬部分種是重要的病原細菌,其寄主范圍廣,廣泛分布在土壤、植物、動物上,其中成團泛菌Pantoeaagglomerans能夠引起稻谷內穎褐變[16],是藜麥葉斑病[17]和煙草細菌性葉枯病的病原菌[18],并能影響紫花苜蓿種子的萌發及幼苗生長[19]等,因此本研究分離得到的泛菌屬細菌是否能引起苦蕎病害,還需進一步鑒定到種,并進行致病性試驗后確認。
基于16S rRNA基因擴增子測序法僅從13份種子樣品中就檢測得到了279個屬的細菌,表明其檢測效率遠高于傳統的分離培養方法,其中泛菌屬為優勢菌群,與分離培養法的結果一致,說明泛菌屬在苦蕎種子中的豐度最高,推測苦蕎所在的環境有利于泛菌屬細菌生存。然而,兩種方法都有彼此未檢測和鑒定到的菌群,如分離培養法未鑒定到高通量測序中菌群豐度高的甲基桿菌屬,16S rRNA基因擴增子測序未鑒定到分離培養法中分離頻率較高的微桿菌屬,可能由于專性厭氧的甲烷細菌屬生長極其緩慢,分離培養比較困難,而16S rRNA基因擴增子測序法在擴增時豐度極高的泛菌屬等其他優勢菌群可能掩蓋了豐度極低的菌群(如微桿菌屬)。因此,想要準確了解種子攜帶細菌的多樣性,還需要分離培養法與高通量測序相結合。
在PCA分析中,來自冕寧縣(MN)的種子和來自喜德縣(XD)、鹽源縣(YY)的種子所攜帶細菌多樣性相近,推測是因為地理上冕寧縣與喜德縣、鹽源縣相鄰,苦蕎品種有交叉使得菌群豐度相似。昭覺縣雖然與喜德縣、普格縣相鄰,但菌群多樣性卻相差很大,可能由于其獨特的地形、環境溫濕度、海拔或品種的原因等引起,不同地點樣品攜帶細菌的多樣性結果還需要通過對更多樣本的檢測進行深入分析。
本研究從苦蕎種子中鑒定到大量菌群,雖然大部分細菌目前還不確定是否是蕎麥的致病菌,但仍有侵染蕎麥或其他作物的可能,因此在蕎麥播種前,要加強種子檢疫和種子處理、選用抗病品種,以提高蕎麥的品質與產量。
猜你喜歡
農產品加工(2023年14期)2023-08-17 10:32:28
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
青年歌聲(2018年5期)2018-10-29 03:18:40
廣東飼料(2016年3期)2016-12-01 03:43:12
海峽科技與產業(2016年3期)2016-05-17 04:32:12
家庭百事通·健康一點通(2016年4期)2016-04-22 09:16:57
