基于靶向代謝組學分析嫁接對茶樹代謝物的影響
2022-10-27 05:14:26茶鳳官趙一明周泳臣左登鴻呂才有丁海琴鄭文忠
食品工業科技 2022年21期
茶鳳官,劉 穎,高 峻, ,孔 勝,趙一明,周泳臣,左登鴻,呂才有,丁海琴,鄭文忠,3
(1.云南農業大學茶學院,云南昆明 650100;2.云南省現代農業茶葉產業體系建設栽培研究室,云南昆明 650201;3.云南省普洱茶樹良種場,云南普洱 665000)
茶樹是我國重要的經濟作物,茶樹的嫁接始于20世紀70年代。目前茶樹嫁接廣泛應用于老茶園及低產茶園的換種改造、野生茶樹的擴種和繁殖以及良種的保存等生產實踐中;同時隨著良種優勢在茶葉生產中逐步得到體現與發揮,低產、低效的老茶園改造已然成為當前茶產業發展中一個重要研究課題。而嫁接后的茶樹因結合了接穗和砧木各自的優勢,往往具有一種或多種顯著的增益效果;與此同時,嫁接后的茶樹利用原有的強大根系吸收水分和礦質營養來滿足接穗新梢發育的需要,所以可以比常規改植換種的茶樹提早2~3年投產,減少老茶園改植換種的投入成本,是老茶園改造、更換新品種等行之有效的方法之一。在生產上,選擇合適的砧木嫁接不僅可以增強對生物或非生物脅迫的抗性,還有助于提高產量、改良品質。
目前嫁接茶樹的相關研究表明,茶樹嫁接會使鮮葉中部分化合物含量發生改變;吳姍等通過HPLS對嫁接茶樹氨基酸含量變化進行分析,結果表明,兩組供試茶樣中都檢出17種游離氨基酸,其中嫁接茶樹中茶氨酸含量都高于對應的接穗品種;另外梁月榮等對嫁接茶樹及相應的接穗品種新梢的茶多酚、氨基酸、咖啡堿和兒茶素等生化成含量進行比較分析,結果表明,嫁接茶樹的氨基酸和咖啡堿含量高于相應的接穗品種,而茶多酚和主要兒茶素類含量低于相應的接穗品種。另外,隨著檢測技術的進步,代謝組學技術已經被廣泛運用于茶葉研究。例如,馬成英等利用UHPLC-QTOF/MS平臺結合代謝組學技術研究嫁接對茶葉次生代謝產物的影響,結果表明,不同砧木嫁接會對茶樹鮮葉的次生代謝產物產生明顯的影響;鄧威威等對茶/油茶嫁接體的次級代謝物含量進行分析發現,嫁接體葉片中的氨基酸、嘌呤堿和多酚含量比油茶葉片高,其中茶氨酸含量比油茶顯著提高,咖啡堿含量比茶樹葉片顯著降低,嫁接體葉片中含有酚酸,而在油茶中未被檢出。
近年來對茶樹嫁接的研究大多都集中在嫁接技術、生物學特性、產量與抗逆性以及品種選育等方面,對于茶樹嫁接后代謝物的變化研究卻很少,且缺乏系統性的研究和分析;所以本文以品種桃形葉為接穗、短接白毫為砧木進行茶樹嫁接,并利用廣泛靶向代謝物學技術分析嫁接前后茶樹代謝物的變化。這在一定程度能夠為嫁接后茶樹鮮葉的加工和生產提供理論參考。
1 材料與方法
1.1 材料與儀器
實驗茶樣來源于云南省普洱良種場茶園,以品種桃形葉為接穗,短接白毫為砧木,采用切接法對茶樹行嫁接;待嫁接成活、生長旺盛后分別采摘其嫩芽(一芽一葉)立即放入液氮中冷凍保存,且每組進行3次生物學重復,具體的樣品名稱和編號如表1所示。甲醇、乙腈 色譜級,Merck公司;標準品 純度≥95%,Bio BioPha公司。

表1 樣品信息及編號Table 1 Sample information and number
Scientz-100F冷凍干燥機 寧波新芝;5424R離心機 艾本德中國有限公司;MM400研磨機 德國RETSCH公司;UPLC SHIMADZU Nexera X2超高液相色譜儀 日本島津公司;Applied Biosystems 4500 Q TRAP質譜儀 AB SCIEX公司;Direct-Q3純水儀 美國默克密理博公司。
1.2 實驗方法
1.2.1 樣品提取 將采摘回來的茶葉嫩芽放入凍干機中進行真空冷凍干燥,并將干燥后的茶樣用研磨機研磨至粉狀,取100 mg碾磨后的組織樣溶解于1.2 mL 70%的甲醇提取液中,將溶液渦旋振蕩后置于4 ℃冰箱過夜,12000 r/min離心10 min后,吸取上清,用0.22 μm微孔濾膜過濾樣品,并保存于進樣瓶中,用于UPLC-MS/MS分析。
質控樣本(QC)由樣本提取物混合制備而成,用于分析樣本在相同的處理方法下的重復性,在儀器分析的過程中,每10個檢測分析樣本中插入一個質控樣本,以監測分析過程的重復性。
1.2.2 色譜條件 色譜柱:AgilentSB-C(1.8 μm,2.1 mm×100 mm);柱溫 40 ℃;流速:0.35 mL/min;進樣量4 μL;流動相A:0.1%的甲酸,流動相B:乙腈,洗脫梯度如表2所示。

表2 色譜階梯洗脫條件Table 2 Chromatographic step elution conditions
1.2.3 質譜條件 ESI源操作參數如下:離子源,渦輪噴霧;源溫度550 ℃;離子噴霧電壓5.5 kV;簾氣流速 25 psi;GSI氣體流速 50 psi;GSII氣體流速 60 psi;碰撞誘導電離參數設置為高;QQQ掃描使用MRM模式,并將碰撞氣體設置為中等。
1.3 數據處理
基于自建數據庫MWDB和二級譜信息進行物質定性;通過三重四級桿篩選出每個物質的特征離子,在檢測器中獲得特征離子的信號強度(CPS),用MultiaQuant軟件打開樣本下機質譜文件,根據代謝物保留時間與峰型的信息對色譜峰的進行積分和校正工作,每個色譜峰的峰面積(Area)代表對應物質的相對含量,然后導出所有色譜峰面積積分數據保存并利用(Unit Variance Scaling)進行數據的歸一化處理,最后得到數據的定性和定量結果。利用Office 2010和Origin 2019進行數據的基本處理和相關圖表的繪制;通過R軟件中的MetaboAnalystR包進行多元統計分析。
2 結果與分析
2.1 樣本質控(QC)分析
通過對不同質控QC樣本質譜檢測分析的總離子流(TIC)圖進行重疊分析,可以判斷代謝物提取和檢測的重復性,能為數據的重復性和可靠性提供重要的保障。QC質控樣本的總離子流圖如下圖1所示,代謝物檢測總離子流的曲線重疊性高,即保留時間和峰強度均一致,表明樣品離散少,儀器穩定,檢測結果可靠。

圖1 QC樣本質譜檢測TIC重疊圖Fig.1 TIC overlap diagram of QC sample mass spectrometry detection
2.2 主成分分析
PCA 分析(Principal Component Analysis),即主成分分析,PCA分析本質上是一種無監督的多元統計分析方法,能從總體上反應各組樣本之間的總體差異和組內樣本之間的變異度大小;PCA結果如圖2所示,第一主成分(PC1)的貢獻率為63.28%,第二主成分(PC2)的貢獻率為10.56%,A1和B1兩組樣品表現出明顯的分離趨勢,說明嫁接前后的茶樹代謝物具有較大差異。

圖2 樣品質譜數據PCA得分圖Fig.2 PCA score of sample mass spectrometry data
2.3 代謝物差異分析
嫁接前后茶樣中共檢測到804種代謝物(包括正離子和負離子兩種模式);為了進一步分析茶樹嫁接前后代謝物的變化情況,利用偏最小二乘法判別(OPLS-DA)進行多元統計分析,同時基于OPLSDA模型中的變量重要性投影(VIP)值和差異倍數(FC)值進行差異代謝物的篩選;當代謝物同時滿足VIP≥1和FC值≥2或≤0.5這兩個條件時,則認為該代謝物具有顯著的差異性;如下圖3所示,兩組樣品中共篩選出10類105種具有顯著差異的代謝物;與嫁接前相比,嫁接后的茶樣中有44種差異代謝物顯著上調,61種差異代謝物顯著下調,差異代謝物下調的數目大于上調,在這些差異代謝物中占比較多的是黃酮、脂質、酚酸、其他和核苷酸及其衍生物這5種類別,分別占差異代謝物總數的31%、15%、9%、9%和8%。在105種具有顯著差異的代謝中有15種脂質、9種黃酮、9種核苷酸及其衍生物、6種鞣質、4種酚酸、4種生物堿、3種有機酸、2種氨基酸及其衍生物和10種其他物質在嫁接前的茶樣中相對含量更高;而有25種黃酮、6種酚酸、5種氨基酸及其衍生物、4種有機酸、1種脂質、1種鞣脂、1種生物堿、1種木脂素和香豆素物質在嫁接后的茶樣中相對含量更高。
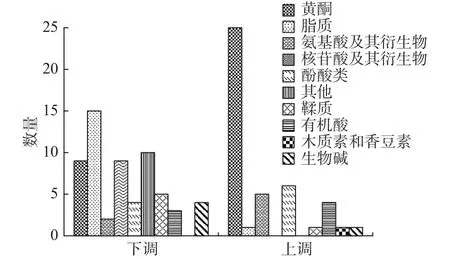
圖3 A1 vs B1差異代謝物分類圖Fig.3 A1 vs B1 differential metabolite classification map
為了更加直觀地比較茶樹在嫁接前后差異代謝物相對含量的變化情況,利用R軟件包繪制了如圖4所示的差異代謝物分類熱圖;結果表明,與嫁接前相比,嫁接后的茶樹大部分的黃酮類物質的相對含量顯著增加,而脂質類、核苷酸及其衍生物和以糖醇類為主的其他類物質的相對含量則顯著減少。

圖4 差異代謝物分類熱圖Fig.4 Heat map of differential metabolite classification
為了更清楚地了解茶樹在嫁接前后代謝物差異倍數的變化情況,制作了如圖5所示的代謝物差異倍數柱狀圖,并將上調和下調的差異代謝物中排名靠前的20位代謝物進行展示。嫁接后茶樹中有7種黃酮(山奈酚-3-O-桑布雙糖苷、山奈酚-3-O-蕓香糖苷(煙花苷)、木犀草素-7-O-(2''-O-鼠李糖基)蕓香糖苷、葫蘆巴堿、木犀草素-6-C-葡萄糖苷-7-O-(6''-對香豆酰)葡萄糖苷、山奈酚-3-O-刺槐糖苷-7-O-鼠李糖苷(刺槐苷)、5,7,4'-三羥基異黃酮-7-O-半乳糖苷-鼠李糖、金圣草黃素-8-C-葡萄糖苷(金雀花素)),2 種氨基酸(N--乙酰-L-谷氨酰胺、N-乙酰-L-亮氨酸)的相對含量要明顯高于嫁接前;而有3種其它類物質(D-果糖-1,6-二磷酸、吡哆素、1-(甘油-3-磷酸)-1D-肌醇)、2 種有機酸(2-羥基異己酸、DL-甘油醛-3-磷酸),2種酚酸(紫丁香苷、3,4'-二羥基-3'-甲氧基苯戊酸),1 種黃酮(山奈酚-3-O-(2-O-木糖基-6-O-鼠李糖基)葡萄糖苷),1 種鞣脂(2,3-二-O-沒食子酰-D-葡萄糖)和1種核苷酸及其衍生物(2-脫氧核糖-5'-磷酸)的相對含量則明顯低于嫁接前。說明嫁接后代謝物中相對含量較高的是黃酮和氨基酸,而嫁接前相對含量較高的是有機酸和酚酸。

圖5 代謝物差異倍數柱狀圖Fig.5 Histogram of metabolite difference multiple
2.4 差異代謝物通路分析
利用KEGG數據庫對差異代謝物進行通路富集分析,結果表明,共有38條通路,且差異代謝物主要分布在嘌呤代謝、煙酸酯和煙酰胺代謝、甘油磷脂代謝、類黃酮生物合成、黃酮和黃酮醇生物合成、光合生物的固碳作用、咖啡堿代謝、次級代謝產物的生物合成等20條代謝途徑中,如圖6所示。另外,在篩選出的105種具有顯著差異的代謝物中被KEGG注釋到的有24種,主要包括7種核苷酸及其衍生物、6種黃酮、3種其他、2種氨基酸及其衍生物、2種生物堿、2種有機酸、1種酚酸和1種脂質;差異代謝物被注釋最多的是黃酮和核苷酸及其衍生物這2種類別;其中核苷酸及其衍生物類主要參與嘌呤代謝、次級代謝產物的生物合成和咖啡因代謝等多條代謝通路,且被KEGG注釋到的尿苷5'-二磷酸、7-甲基黃嘌呤、3-甲基黃嘌呤、鳥嘌呤、黃苷、2'-脫氧肌苷-5'-單磷酸和2-脫氧核糖-5'-磷酸這7種核苷酸及其衍生物均全部顯著下調;而黃酮類物質主要參與類黃酮生物合成、黃酮和黃酮醇生物合成、次級代謝產物的生物合成等多條代謝通路,且被KEGG注釋的6種黃酮中山奈酚-3-O-蕓香糖苷(煙花苷)、山奈酚-3-O-鼠李糖苷(阿福豆苷)(番瀉葉山奈苷)、矢車菊素-3-O-(6''-O-對香豆酰)葡萄糖苷、根皮素和芹菜素這5種代謝物則顯著上調,而柚皮素-7-O-葡萄糖苷(櫻桃苷)則顯著下調。

圖6 嫁接前后差異代謝物KEGG富集圖Fig.6 KEGG enrichment diagram of differential metabolite before and after grafting
3 討論
嫁接是茶樹良種繁育最直接有效的方法之一,通過嫁接可以最大程度地保留接穗品種的優良特性,而茶樹作為一種重要的經濟作物,其主要的價值就在茶樹的鮮葉上。鮮葉質量是導致茶葉品質形成的重要因素之一,而鮮葉質量在很大程度是由里面的內含物質含量所決定的,所以研究茶樹嫁接前后代謝物含量的變化,能夠為嫁接茶樹的茶葉加工提供一定的理論參考。
通過前文的研究發現,茶樹嫁接前后代謝物具有明顯的差異性,分析代謝物的差異倍數可知,導致差異的主要原因可能是嫁接后茶樹中氨基酸和黃酮類物質大量積累,以及酚酸和有機酸等物質的消耗。另外,通過對KEGG代謝通路的分析,發現黃酮類物質顯著上調,而核苷酸及其衍生物則顯著下調;這可能與嫁接后茶樹中相關代謝通路中酶的活性或者基因的表達有關,但目前針對茶樹嫁接后代謝物的變化的研究較少,因此,嫁接茶樹代謝物的具體變化機理還需做進一步探索。除此之外,吳珊等研究也發現嫁接后茶樹體內游離氨基酸含量有所增加,同樣王文建將鐵觀音嫁接到6個不同品種砧木上,結果表明嫁接后的茶樹中氨基酸、茶多酚和水浸出物等含量增加,這與本文的研究結果大體一致。所以在接下來的研究中可以通過代謝組和轉錄組聯合分析的方法,對相關通路中的差異代謝物進行進一步的研究和分析。
4 結論
利用廣靶代謝組學技術對茶樹(桃形葉)嫁接前后的代謝物進行檢測,兩組茶樣PCA表現出明顯的分離趨勢,說明茶樹嫁接前后代謝物具有較大的差異。另外,嫁接前后的茶樣中共篩選出10類105種具有顯著差異的代謝物,這些差異代謝物中的大部分黃酮類和氨基酸及其衍生物相對含量明顯增加,而脂質類、核苷酸及其衍生物、酚酸類等物質的相對含量則明顯減少,同時,分析代謝物的差異倍數,發現在嫁接后的茶樹中相對含量較高的代謝物是黃酮和氨基酸,而嫁接前相對含量較高的代謝物是酚酸和有機酸,這些滋味物質含量的變化會對茶樹鮮葉品質造成影響。此外,通過KEGG代謝通路的分析,發現代謝物在38條代謝通路中富集,且大部分的差異代謝物分布在嘌呤代謝、類黃酮生物合成、次級代謝產物的生物合成等20條代謝途徑中;與此同時,被KEGG注釋到的大部分黃酮類物質顯著上調,而核苷酸及其衍生物則顯著下調。綜上所述,品種桃形葉通過嫁接會使代謝物的含量發生明顯的改變,這在一定程度上能夠為嫁接茶樹的茶葉加工、生產等提供參考。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
中成藥(2017年4期)2017-05-17 06:09:50
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
