不同pH條件下銻對水稻幼苗的毒害效應
2022-11-04 09:42:50姚麗華查建軍李亞麗潘小翠于紅梅束良佐
農業環境科學學報 2022年10期
姚麗華,查建軍,李亞麗,潘小翠,于紅梅,束良佐*
(1.浙江師范大學地理與環境科學學院,浙江 金華 321000;2.南方科技大學臺州研究院,浙江 臺州 318000;3.臺州學院生命科學學院,浙江 臺州 318000)
銻(Sb)是一種有毒的類金屬元素,在地殼中的自然含量約為0.2~0.3 mg·kg-1,土壤中銻的背景含量約為0.3~2.3 mg·kg-1[1]。我國是產銻大國,銻礦儲量位居世界第一,產量約占世界總產量的79%[2]。近年來,由于銻礦開采以及含銻產品的不合理利用,導致環境中銻的含量急劇上升[3-4],尤其是湖南、貴州、廣西等銻礦較集中的地區土壤銻含量遠超過其土壤背景值[5-8]。宋書巧等[9]在廣西刁江沿岸農田土壤的調查分析中發現,該地土壤中的銻含量高達1 000 mg·kg-1。何孟嘗等[1]對湖南錫礦山銻礦附近不同地點取樣調查發現,該區域土壤的銻濃度為100.6~5 045.0 mg·kg-1。環境中高含量的銻會影響作物生長,進而通過食物鏈被人體吸收,危害人體健康。
水稻(Oryza sativa)是世界第二大糧食作物,在我國三大糧食作物中產量居首位,其播種面積約占糧食作物播種總面積的30%,產量約占糧食總產量的40%[10-11]。香港食品安全指南限制了水稻中允許的最大銻含量為1 mg·kg-1[12]。然而,在銻污染地區種植的水稻中銻含量可高達5.79 mg·kg-1[13]。何孟嘗等[14]對湖南省錫礦山銻冶煉廠周邊生長的水稻進行調查研究,發現水稻體內富集了大量銻,其含量具體表現為根>莖>葉。水稻對銻具有一定的吸收富集能力,且水稻是銻礦富集區主要的糧食作物。對銻礦山周邊居民日常攝入食物中銻來源調查表明,攝入的銻中有33%來源于稻米,因此,種植和食用水稻已成為礦區居民接觸銻的主要途徑[15],這對礦區居民的健康造成很大的危害。
環境的pH值對植物吸收利用土壤養分有著重要影響,同時也是影響土壤中重金屬存在形態與活性的關鍵因素[16]。WILSON等[17]繪制了不同系統下的pe-pH圖,表明不同的pH值使得銻有著不同的沉淀形態和離子形態。酸性環境下,礦石中的碳酸鹽、碳酸氫鹽和氧化物會不斷溶解,使得銻從沉淀形態轉換為離子形態,并且礦石結構會變得十分疏松,進一步促進銻的釋放[18-19]。此外土壤吸附重金屬的能力以及重金屬的遷移能力在很大程度上也受土壤pH值影響。TIGHE等[20]研究發現土壤中非結晶Fe氫氧化物對Sb(Ⅴ)的吸附量在pH為4時最大。CAI等[21]研究也發現銻在鈣質土壤中的遷移性和pH有關,鈣質土壤的pH大于7是銻遷移性高的重要原因。因此,不同的pH在不同的土壤環境中對銻的影響存在差異。由于土壤中重金屬的存在形態和遷移能力會嚴重影響植物對重金屬的吸收量,且我國銻污染區域土壤pH差異較大[22-23],但是pH對水稻吸收銻的影響機制尚不明確。
本研究以水稻為研究對象,通過水培試驗,探究不同pH條件下銻污染對水稻幼苗生長及生理機制的影響,為銻污染區水稻的安全生產提供參考依據。
1 材料與方法
1.1 試驗材料
化學試劑:酒石銻酸鉀(C8H4K2O12Sb2),分析純;供試材料選取水稻(Oryza sativaL.),品種為秈粳雜交稻嘉優中科3號(浙江省臺州市臺農種業有限公司)。
1.2 試驗方法與處理
挑選籽粒飽滿的種子用5%NaClO溶液消毒20 min后,用去離子水清洗干凈,將消毒洗凈的種子放入32℃的恒溫恒濕培養箱中避光催芽,催芽期間每日更換3次蒸餾水,3 d后將露白萌發的種子均勻灑在育苗盤(含1/2強度營養液)中培養,育苗盤中的營養液以剛好浸沒種子為宜。待幼苗長至兩片真葉時,挑選6株長勢基本一致的幼苗(用海綿固定在帶孔的蓋子上)移栽到2 L黑色塑料水培桶,水培桶中含有全強度的木村B營養液[24],每桶設置3穴,每穴2株。營養液 配 方為0.36 mmol·L-1(NH4)2SO4,0.55 mmol·L-1MgSO4·7H2O,0.36 mmol·L-1Ca(NO3)2·4H2O,0.27 mmol·L-1K2SO4,0.18 mmol·L-1KH2PO4,9.10 μmol·L-1MnCl2,20 μmol·L-1H3BO3,0.77 μmol·L-1ZnSO4·7H2O,0.32 μmol·L-1CuSO4·7H2O,0.15 μmol·L-1(NH4)6Mo7O24·4H2O,20 μmol·L-1Fe-EDTA。幼苗生長前期每隔1 d換一次營養液,待苗培養至30 d后每日更換營養液。用0.1 mol·L-1NaOH或HCl調節pH值,每日3次,保持pH值處于設定梯度,期間pH值變化不超過0.5。試驗在人工培養室中進行,光源由LED燈提供,光照時間設置為12 h·d-1,光照強度為200 μmol·m-2·s-1[25-26],溫度為26.0±1℃,相對濕度為60%~70%。
待幼苗在全強度營養液中適應一周后,以酒石銻酸鉀的形式向營養液中加入銻,銻濃度分別為0、0.016 mmol·L-1,該濃度的設置基于向猛等[26]和LONG等[27]的研究。銻處理40 d后收樣。用NaOH或HCl將營養液的pH值調節至4.0、5.0、6.0、7.0共4個梯度,因此試驗共8個處理:(1)pH=4 Sb0(營養液內不加Sb);(2)pH=5 Sb0;(3)pH=6 Sb0;(4)pH=7 Sb0;(5)pH=4 Sb(營養液內含0.016 mmol·L-1Sb,下同);(6)pH=5 Sb;(7)pH=6 Sb;(8)pH=7 Sb。每個處理重復4次,共32盆。在幼苗生長期內每日隨機調整桶的位置。
1.3 測定項目及方法
1.3.1 葉綠素含量的測定
選取植株第3片葉的中上部位(避開大葉脈),采用SPAD-502型便攜式葉綠素儀多點測定SPAD值。每盆測定4株,取各株平均值代替該盆水稻葉片葉綠素含量。
1.3.2 葉綠素熒光參數的測定
在水稻幼苗培養至第40天時,選取第3片葉片用暗適應葉夾對葉片進行充分的暗適應,30 min后,用葉綠素熒光儀MINI-PAM-II(Walz,德國)測定不同處理下水稻葉片葉綠素熒光參數,包括PSⅡ電子傳遞速率(ETR)、光化學猝滅系數(qP)和非光化學猝滅系數(NPQ)、最大光化學效率(Fv/Fm)。每盆選取4株水稻葉片測量并取其平均值代表該重復。
1.3.3 葉綠素熒光快速光響應曲線的測定
待幼苗培養至第41天,利用葉綠素熒光儀MINIPAM-II(Walz,德國)測定不同處理下水稻葉片的快速光響應曲線。儀器內置光強(PAR)梯度分別為0、63、86、120、184、276、406、607、793、1 110、1 447 μmol·m-2·s-1,每個光強梯度持續20 s,每盆以不同水稻葉片重復測量5次,取平均值。
1.3.4 幼苗生長特性的測定
每盆選4株水稻植株用卷尺測量其株高,并用直尺測定中上部充分展開葉片的葉寬、葉長。根據公式計算該葉片面積[28],葉面積(cm2)=長度(cm)×寬度(cm)×K,校正系數K=0.75。
1.3.5 保護酶和丙二醛含量
超氧化物歧化酶(SOD)活性采用抑制氮藍四唑光還原比色法測定;過氧化物酶(POD)活性采用愈創木酚法測定;過氧化氫酶(CAT)活性采用紫外吸收法測定;丙二醛(MDA)含量采用硫代巴比妥酸比色法[29]測定。均采用鮮樣含量。
1.3.6 根系與地上部銻的含量
銻處理40 d后收樣,將植株地上部和根部用去離子水洗凈后,放置于70℃烘箱中干燥48 h后研磨,稱取過200目篩的植物樣品0.1 g于聚四氟乙烯管中,并加入2 mL HNO3和1 mL H2O2溶液密封24 h后,在180℃的烘箱中消化12 h,冷卻至室溫,然后放置在160℃的電熱板上加熱趕酸,直至液體呈黃豆粒大小,用去離子水將其轉移至50 mL容量瓶,定容、過濾。采用電感耦合等離子體質譜儀(ICP-MS)測定水稻各部位銻含量。同時做空白和標準樣品進行質量控制。
1.4 數據處理
試驗數據采用Excel 2016處理,結果以平均值±標準誤差表示,采用SPSS 22.0軟件對數據進行統計分析,在95%置信水平下,采用LSD法進行單因素方差分析,復合實驗采用雙因素方差分析。采用Sigma-Plot 14.0軟件制圖。
2 結果與討論
2.1 不同pH處理對銻脅迫下水稻植株銻吸收的影響
銻脅迫下,不同pH處理對水稻植株各器官銻的吸收量存在顯著差異(表1)。在pH 7條件下,水稻根部吸收的銻含量分別較pH 4、pH 5、pH 6條件下減少了21.76%、27.30%、25.12%。pH 4處理時,水稻植株地上部吸收的銻含量達到167 mg·kg-1,是中性pH(pH 6、pH 7)處理均值的1.41倍,與pH 5處理之間的差異未達到顯著水平。這可能是營養液中酸性條件下銻主要以Sb(Ⅲ)為主,隨著pH增加水稻根表的鐵斑會將其氧化形成Sb(Ⅴ)[30],水稻對Sb(Ⅴ)的吸收能力弱于Sb(Ⅲ),進而導致水稻中吸收的銻含量減少[31]。
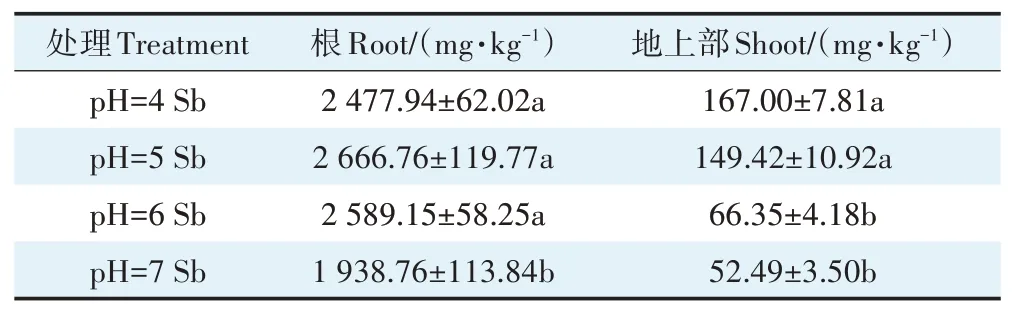
表1 不同pH值條件下銻處理對水稻植株銻含量的影響Table 1 Effects of Sb stress on Sb uptake in rice plants under different pH conditions
2.2 對水稻生長特性的影響
銻對植物是一種非必需元素,當植物體內銻累積到一定含量時會對植物產生毒害效應[32]。具體表現為植株矮小、葉片發黃、生長緩慢以及生物量減少等。經銻處理的植株,其株高均顯著低于無銻處理組(表2),pH 7、pH6、pH5、pH 4分別下降了4.3%、4.98%、11.72%和9.16%。但銻處理后的葉片長、水稻葉寬和葉面積受毒害作用的大小隨pH變化存在較大差異。從pH 7降到pH 4,地上部株高和葉片長分別降低了4.09%、4.42%,葉寬、葉面積未產生顯著性變化。低pH條件和銻處理均會顯著降低水稻地上部及根部的生物量(圖1)。這與前人在水稻、玉米和小麥中的研究結果相似[33-36]。植物吸收的銻會占用必需元素Fe2+、Zn2+、Ca2+等轉運離子通道并與鋅、鈣轉運蛋白結合,從而影響蛋白質結構及功能,破壞植物體內離子平衡,引起植株營養失衡[36],同時又會抑制可溶性蛋白質、可溶性糖和淀粉的合成進而影響植物的生長[37]。由表2可知,pH和銻對水稻地上部生長特性均有極顯著的交互作用(P<0.01),兩者的交互作用使低pH條件(pH 4、pH 5)下水稻的株高、葉長下降均值是中性pH條件(pH 6、pH 7)的2.25倍和2.29倍;水稻葉寬、葉面積也較中性pH下減少了18.32%和23.95%。隨著pH降低,水稻植株地上部和根部的生物量下降幅度也逐漸增加。這表明重金屬脅迫和低pH可能產生復合脅迫,進一步抑制水稻的生長。一方面在低pH條件下,水稻幼苗根系細胞膜透性增大,促進銻的跨膜運輸,增加細胞內銻的含量,加重銻對根系的傷害,另一方面銻脅迫下,水稻植株根系的細胞膜被破壞,H+-ATP酶活性受到抑制,增加細胞外H+進入,減少細胞內H+流出,進而導致細胞質酸化加劇,使根系損傷加重[38-39]。
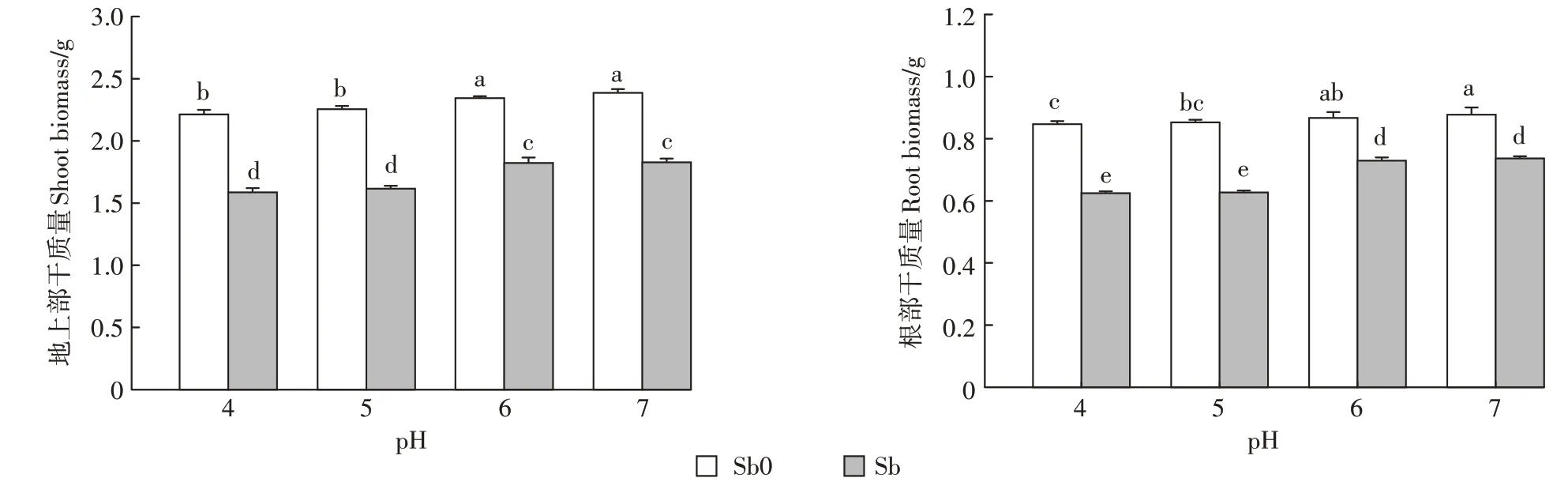
圖1 不同pH條件下銻對水稻生物量的影響Figure 1 Effects of antimony on biomass accumulation of rice seedlings under different pH conditions
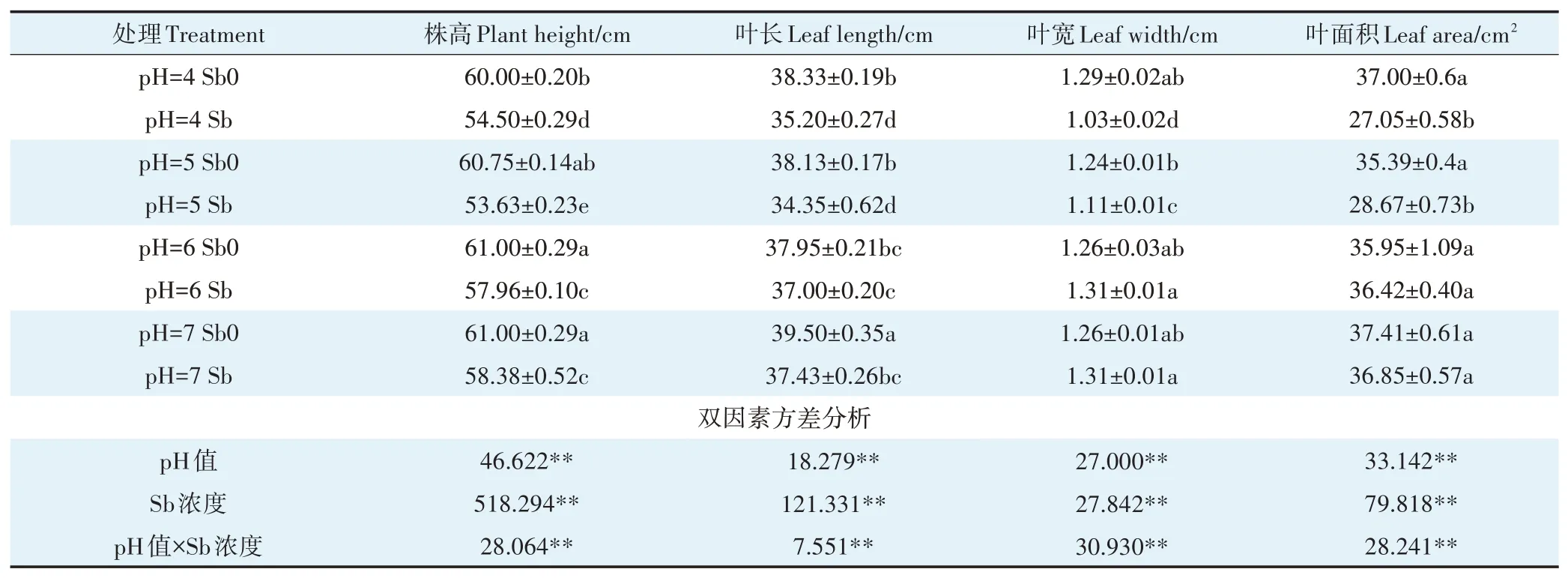
表2 不同pH條件下銻對水稻地上部生長的影響Table 2 Effects of Sb on shoot growth of rice seedlings under different pH conditions
2.3 對水稻葉片SPAD值的影響
葉綠素是植物光合作用的物質基礎,其含量高低在一定程度上能夠反映植物葉片光合作用功能強弱。本研究中經銻處理的植株,pH 4、pH 5、pH 6和pH 7條件下葉片SPAD值較無銻處理的植株分別下降了14.64%、15.15%、12.73%和12.23%(圖2)。一方面銻脅迫可能會抑制有關葉綠素合成酶的活性,使葉綠素的合成受阻;另一方面會大量誘導產生的活性氧破壞細胞膜的結構與功能,使葉綠體膜膨脹破裂,進而導致葉綠素被降解[40-41]。pH 6和pH 7條件下水稻葉片SPAD值無顯著性差異,但其均值較低pH條件(pH 4、pH 5)顯著增加。由方差分析結果可知,銻處理和pH值對水稻葉片SPAD值存在極顯著的交互作用(P<0.01),在低pH條件下(pH 4、pH 5)降幅高于中性pH處理。這和前人在花椰菜、菹草上的研究結果相似[16,42],其研究表明植物在酸性條件下易失綠,葉綠素轉變為葉褐素,進而影響植物的光合作用。本研究結果也表明在相同銻濃度下,pH 6、pH 7條件下水稻葉片光合色素遭受的損傷較小,受銻脅迫影響較輕。磷、鎂、鉀等營養元素在保證葉綠體膜的結構和功能上至關重要,而在pH處于中性條件時大多數營養元素有效性高,促進水稻植株對營養元素的吸收[43],從而在一定程度上緩解了銻脅迫造成的水稻吸收營養元素減少帶來的毒害效應[44]。
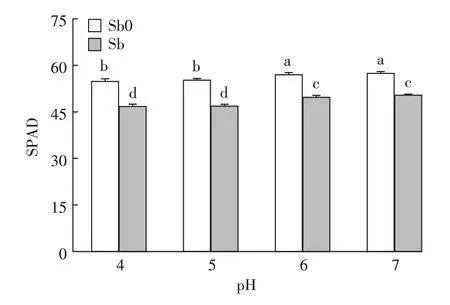
圖2 不同pH條件下銻對水稻葉綠素SPAD值的影響Figure 2 Effects of antimony on chlorophyll SPAD values of rice seedlings under different pH conditions
2.4 對水稻葉綠素熒光特性的影響
葉綠素熒光參數是探討植物光合性能的重要探針[41]。Fv/Fm能夠反映PSⅡ原初光能轉化效率,Y(Ⅱ)反映植物葉片實際光合效率,qP表征PSⅡ反應中心吸收的光能用于光化學電子傳遞的份額,ETR反映實際光反應過程中的表觀電子傳遞效率[45-46]。PSⅡ是植物光合作用的重要部位,同時也是重金屬脅迫的重要位點[45]。研究表明重金屬脅迫會影響水裂解端的電子流,抑制PSⅡ活性,降低PSⅡ的原初光能捕獲能力和電子傳輸能力,從而抑制植物光合作用[46-47]。本試驗中經銻處理的植株,Fv/Fm、Y(Ⅱ)、ETR、qP值均顯著下降(表3),其中ETR值在pH 4、pH 5、pH 6和pH 7條件下較無銻處理分別下降了17.32%、17.32%、14.65%和14.11%;而表征熱耗散引起的熒光猝滅qN、NPQ表現出的規律正好相反,即銻處理后,qN、NPQ值均顯著上升,其在各pH下的均值分別上升了25.67%、16.83%。Fv/Fm、Y(Ⅱ)、ETR和qP值均隨pH的降低而降低,其中在pH 7和pH 6之間無顯著性差異,但均高于pH 5和pH 4處理;而qN值和NPQ值則在低pH下增加。這說明銻脅迫破壞了光合機構,使PSⅡ反應中心光能轉換效率和電子傳遞能力降低,進而抑制了植物的光合作用;而幼苗能夠通過自身的調節機制將過多激發能以熱能的方式耗散,從而降低對PSⅡ反應中心的破壞程度,實現植株的自我保護。據雙因素方差分析可知銻處理和pH值對Fv/Fm、Y(Ⅱ)、qP、qN和NPQ值均有顯著的交互作用,使Fv/Fm、Y(Ⅱ)、qP值在低pH條件下的下降均值是中性pH條件下的2.22、1.48倍和1.65倍,而二者交互作用對ETR值未產生顯著性影響。這說明銻脅迫對水稻幼苗光合作用的破壞程度在低pH條件下最重,可能是在低pH條件下,葉片內蛋白酶活性降低,相關電子傳遞受阻,光能轉換效率隨之降低[48];且幼苗在低pH和銻脅迫下qN和NPQ值增幅最大,一方面反映了葉片的自我保護機制,另一方面也表明了幼苗吸收的大量能量以熱能形式耗散,只有小部分能量被利用,從而導致其不能更好地進行光合作用。
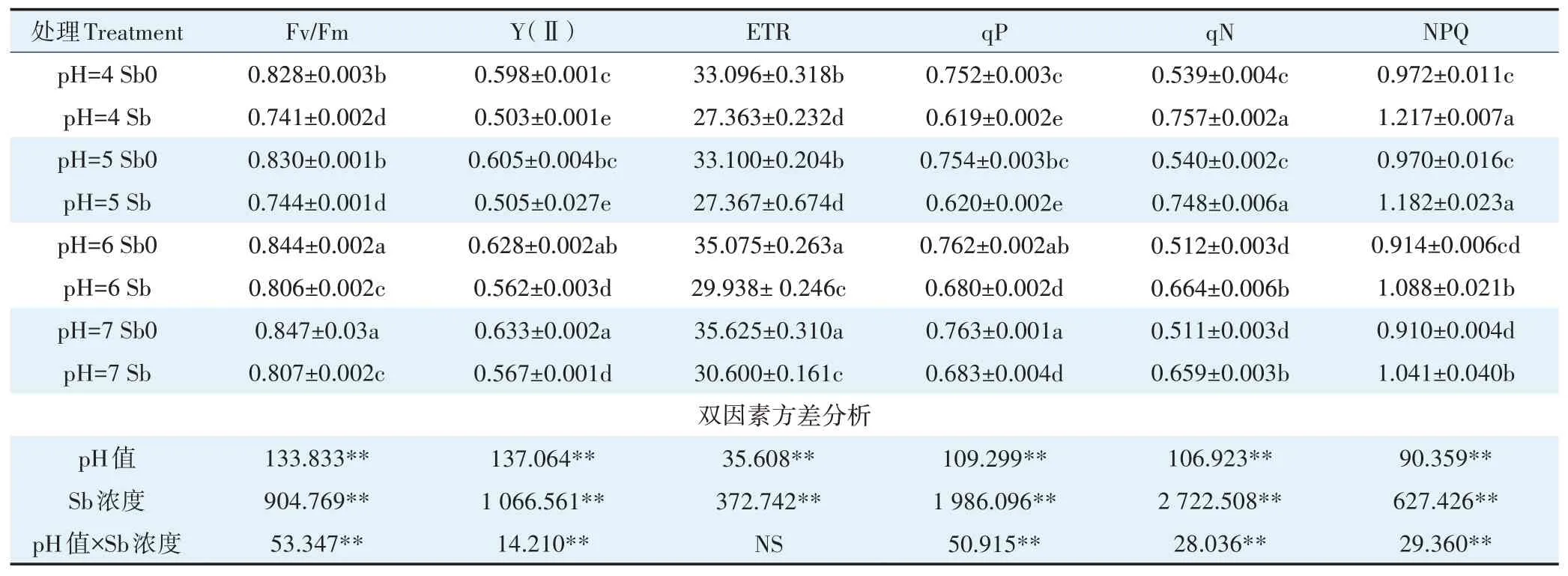
表3 不同pH條件下銻對水稻葉綠素熒光的影響Table 3 Effects of Sb on chlorophyll fluorescence of rice seedlings under different pH conditions
2.5 對水稻葉綠素快速光響應曲線的影響
α值、ETRm值、IK值是反映植物光合能力的重要指標。不同pH下(pH 4~pH 7)銻處理使得水稻葉片的α值、ETRm值、IK值均較無銻處理分別下降了19.75%、24.15%和15.80%(表4);α值、ETRm值、IK值均隨pH值升高而上升(表4),與pH 4相比,α值、ETRm值、IK值在pH 7條件下分別增加了0.07、0.15、0.05倍。pH值和銻濃度的交互作用對α、ETRm和IK的影響雖未達到顯著水平(P<0.01),但均在低pH時下降幅度最大,其下降均值分別是中性pH條件下的1.15、1.14、1.07倍。這表明銻脅迫和低pH均會破壞植物的光合中心,降低植物的光合能力;且銻脅迫下,低pH值會加重破壞程度。

表4 不同pH值條件下Sb對水稻葉綠素熒光光響應曲線參數的影響Table 4 Effects of Sb on parameters of chlorophyll fluorescence photoresponse curve of rice under different pH conditions
2.6 對水稻抗氧化酶和MDA含量的影響
逆境脅迫會大量誘導植株產生活性氧自由基,產生過氧化反應,使細胞膜系統受損,進而影響植物的生長[49],其產物MDA含量常作為反映細胞脂質過氧化強度和質膜受破壞程度的重要參數[50]。本試驗中在銻處理后MDA含量顯著增加了28.27%,表明細胞膜受到破壞。植株為了抵抗這種逆境脅迫,自身會產生一種抗氧化酶保護系統來清除過多的自由基并維持正常生長[51]。本試驗中銻處理下水稻葉片的SOD、POD、CAT活性顯著增加,其均值較無銻處理分別增加了13.20%、20.28%和21.73%(表5),相較于pH 4,pH 7條件下水稻葉片的SOD、POD、CAT活性分別降低了5.60%、8.33%、12.50%。抗氧化酶(SOD、CAT、POD)活性的增加(表5),提高了清除自由基的能力,從而緩解了活性氧對細胞膜的傷害,但這是防止活性氧自由基累積從而導致植株死亡的刺激性機制[52],故逆境脅迫造成的生長抑制作用仍舊難以避免。方差分析結果可知pH值和銻處理對SOD和POD活性具有顯著的交互作用,而兩者的交互作用對MDA含量、CAT活性未產生顯著性影響,但其均使三種抗氧化酶活性在低pH條件下增長最為明顯,分別是中性pH條件的1.51、1.32倍和1.05倍。這表明在pH 6、pH 7條件下,水稻受銻脅迫引起的氧化損傷較小,在pH 6、pH 7處理組中過氧化反應產物MDA含量增幅較小也可證明這一點。

表5 不同pH值條件下銻脅迫對水稻植株抗氧化酶和MDA含量的影響Table 5 Effects of Sb stress on antioxidant enzymes and MDA content of rice plants under different pH conditions
3 結論
(1)銻處理后,水稻幼苗地上部和根部均大量吸收銻造成銻脅迫,其雖然能夠誘導水稻體內的過氧化氫酶、過氧化物酶和超氧化物歧化酶活性增加,但仍然使丙二醛含量升高,抑制水稻地上部生長,降低干物質質量。
(2)銻處理顯著降低水稻葉片葉綠素相對含量,降低PSⅡ原初光能轉化效率、最大相對電子傳遞效率和半飽和光強等葉綠素熒光參數,同時葉片非熒光猝滅系數顯著提高。銻脅迫通過破壞水稻的細胞膜系統和光合中心來降低水稻光合能力。
(3)營養液在pH=4、pH=5條件下會促進水稻地上部和根部吸收銻,同時加劇銻對水稻的生長、光合性能以及細胞膜的破壞,而在pH=6、pH=7條件下減輕了銻脅迫對水稻的毒害作用。綜上,對存在銻污染風險地區可通過適當改變土壤pH值來緩解銻的毒害作用。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
軍事文摘(2021年22期)2021-11-26 00:43:51
當代陜西(2021年2期)2021-03-29 07:41:24
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
