
植物耐鹽機理研究進展
2022-11-18 06:46:38王瀚祥李廣存徐建飛王萬興金黎平
作物雜志 2022年5期
王瀚祥 李廣存 徐建飛 王萬興 金黎平
(中國農業科學院蔬菜花卉研究所/農業農村部薯類作物生物學和遺傳改良重點實驗室,100081,北京)
土壤鹽漬化導致的耕地土壤退化是全球農業最具破壞性的非生物脅迫之一。據統計,全世界有9.54億hm2土地受鹽漬化影響[1]。鹽脅迫對植物生長的影響體現在植株生長勢下降、衰老甚至死亡[2]。生理上,高鹽引起滲透脅迫,表現為植物根系攝水困難、葉面水分流失加速[3],隨后引發植物體內多種生理變化,如離子紊亂、質膜損傷和光合速率下降等[4]。鹽脅迫還會導致植株體內過量活性氧(reactive oxygen species,ROS)積累,即氧化脅迫。ROS化學性質活潑,對細胞結構和生物功能產生破壞,包括引起膜質過氧化和膜質脫氧化,破壞蛋白質、葉綠素和核酸分子[5]。
植物為適應高鹽環境,進化出了一套復雜的響應機制(圖1)。幾十年來,關于植物耐鹽脅迫的研究主要集中在以下方面:(1)鹽脅迫的信號轉導;(2)滲透保護劑及溶質的生物合成;(3)離子穩態及區域化;(4)抗氧化系統;(5)植物激素;(6)其他方式。本文將從這6個方面綜述前人研究。
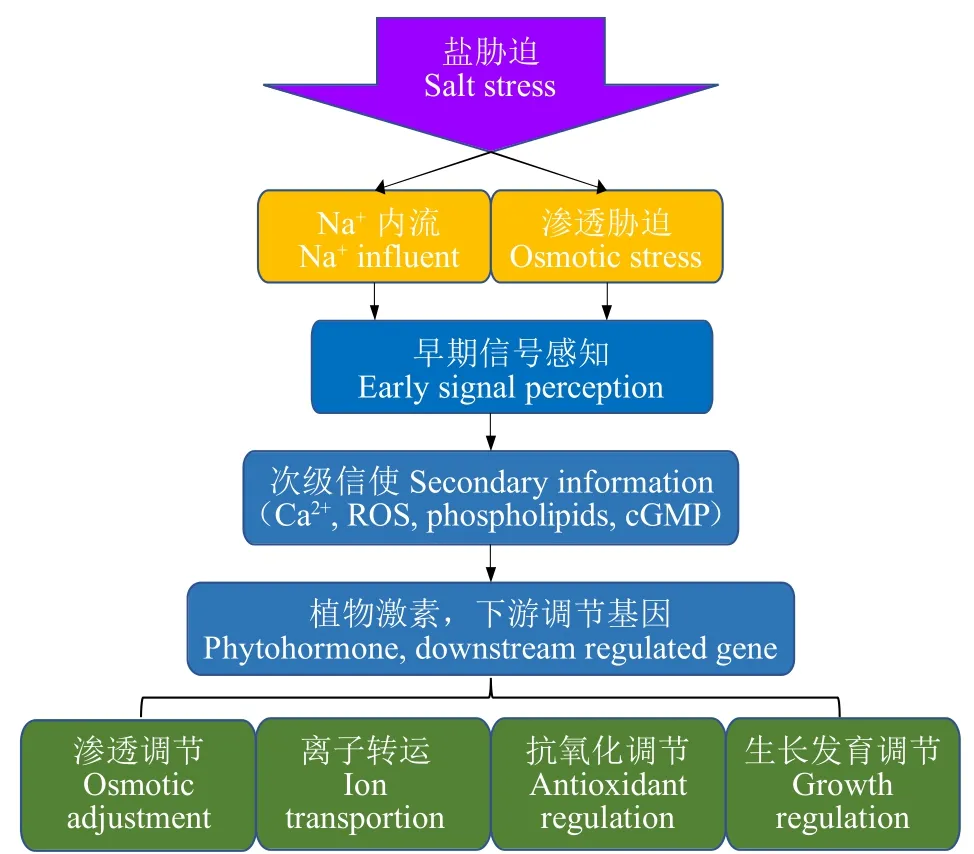
圖1 植物對脅迫信號的轉導Fig.1 Stress signal transduction in plants
1 鹽信號的早期感知
鹽脅迫發生后,植物最先感知鹽信號的分子機制還沒有明確的結論,通常存在3種猜測。一是植物最先感知鹽引起的滲透變化,二是根系中存在Na+感受器,三是鹽誘導的細胞壁變化可以通過受體型激酶(FER)感知。
1.1 滲透感知
高鹽引起的滲透脅迫造成細胞膜或細胞壁上的機械張力,激活了某些鈣離子通道蛋白,使胞質Ca2+濃度迅速增加。OSCA1(reduced hyperosmolalityinduced[Ca2+]iincrease 1)被鑒定為導致這種Ca2+信號的關鍵蛋白[6],是滲透感受器候選蛋白。基于這種蛋白的結構推測,滲透壓力在脂質雙分子層產生張力,導致其構象改變并激活Ca2+通道,允許Ca2+進入[7]。雖然這種信號的轉導途徑還不清楚,但是OSCA1的鑒定為了解植物細胞感知滲透脅迫的途徑開啟了一扇新的大門。
1.2 Na+感受器
MOCA1(monocation-induced[Ca2+]iincrease 1)的鑒定是植物細胞外鹽信號感知的突破性進展。moca1突變體缺乏對Na+、K+和Li+的早期Ca2+信號反應[8]。MOCA1作為葡萄糖醛酸轉移酶,在質膜上產生糖基肌醇磷酸化神經酰胺(GIPC)鞘脂。這些GIPCs可以結合單價陽離子(如Na+)打開Ca2+通道,隨即形成Ca2+波動誘導下游脅迫應答通路的響應[8]。
1.3 細胞壁信號感受器
膜內固有蛋白(如離子通道)可在雙層膜內進行一定程度的運動,這種運動主要受到細胞壁的影響[9]。鹽脅迫和滲透脅迫會引起細胞壁的機械張力產生應激信號,FER通路感知該信號防止細胞破裂[10]。該通路還引發短暫的Ca2+增加,但下游調控模式仍然未知。
1.4 脅迫信號的轉導
植物感受到脅迫信號后,體內出現了3種調控信號因子:3′,5′環鳥苷酸(cGMP)、ROS 和 Ca2+。cGMP抑制Na+內流和K+外流,并促進胞質中Ca2+的增加[11];鹽脅迫會迅速導致質外體ROS分子水平升高,ROS最初與多胺相互作用增加胞內游離Ca2+,并在后續Ca2+的轉運過程中發揮關鍵作用[12-13]。上述調控信號因子主要引起體內Ca2+波動,隨后鈣調磷酸酶B型蛋白(CBLs)結合Ca2+,通過CBLs-蛋白激酶(CIPKs)互作引起蛋白磷酸化,激活耐鹽調控通路。
2 滲透調節物質的合成
植物為應對高鹽引起的滲透脅迫,體內合成脯氨酸(proline,Pro)、可溶性糖、甜菜堿(betaines)和多元醇(polyhydric alcohols)等滲透調節物質,在細胞水平促進滲透平衡,保持細胞膨壓和正常的新陳代謝活動[14]。
Pro作為一種重要的滲透調節物質,以自由狀態存在于植物細胞中,在生理pH范圍內具有分子量小、水溶性高、無凈電荷等特點。Pro作為滲透保護劑維持鹽脅迫下植物的生長,保護光合系統,維持離子穩態,增強抗氧化系統[15-16]。Pro在植物體內主要由谷氨酸(Glu)通路合成,Glu在吡咯啉-5-羧酸合成酶(P5CS)催化下生成谷氨酸-1-半醛(GSA),GSA自發環化為吡咯琳-5-羧酸(P5C),在吡咯琳-5-羧酸還原酶(P5CR)的作用下還原為Pro。Pro在植物體內的降解基本上是合成過程的逆反應,首先發生在線粒體中,Pro在線粒體中由脯氨酸脫氫酶(ProDH)催化生成P5C,P5C在吡咯琳-5-羧酸脫氫酶(P5CDH)作用下生成Glu。鹽脅迫下植株體內Pro積累的分子機制已被闡明,在非脅迫環境下,磷酸酶D(PLD)是Pro積累的反向調控因子;鹽脅迫下,鈣信號和磷脂酶C(PLC)觸發P5CS轉錄和Pro積累[17],Pro的積累反饋抑制P5CS的活性[18]。脅迫結束后,Pro的分解代謝被激活,主要控制基因為PDH與P5CDH[17]。
可溶性糖主要包括葡萄糖、蔗糖和海藻糖,主要功能是穩固質膜和原生質體,保護可溶性酶[19-20],為有機物的合成提供能量。鹽脅迫環境下,土壤滲透勢的增大導致植物吸水困難,此時植物通過積累一些小分子可溶性糖來緩解這種滲透壓力[21]。
甜菜堿主要包括甘氨酸甜菜酯(glycine betaine)、β-丙氨酸甜菜堿(β-alanine betaine)和脯氨酸甜菜堿(proline betaine)。甘氨酸甜菜酯的研究最廣泛,它在植物應對非生物脅迫方面起到提高細胞的滲透壓、保護細胞、穩固蛋白質、保護光合系統和提高抗氧化酶活性等作用[22-23]。甘氨酸甜菜堿在調控基因表達方面也有重要作用,它可以誘導特定基因(尤其是抗氧化酶)和轉錄因子的表達[24-25]。甘氨酸甜菜堿主要由膽堿合成,膽堿經膽堿單加氧酶(CMO)和甜菜堿醛脫氫酶(BADH)的先后酶促作用轉化為甘氨酸甜菜堿,這2個基因可以通過其順式調控元件(CREs)參與不同生物過程的轉錄調控,如脅迫和激素反應[26]。研究[27-28]發現,敲除CMO基因的甜菜植株比野生型甜菜植株更容易受到鹽脅迫,BADH也在諸多轉基因試驗中被證明對植物耐鹽性有促進作用。
多元醇是具有多個羥基的化合物,可作為有機反應原料。多元醇的積累,無論是甘露醇和山梨醇等直鏈代謝物,還是肌醇及其甲基化衍生物等環狀多元醇,都與耐旱或耐鹽有關,這是基于多元醇在許多物種中的分布情況而得出的結論[29]。植物遭受脅迫時,會激活依賴NADPH的甘露糖-6-磷酸還原酶,生成甘露醇,這類滲透調節物質發揮著保護質膜及酶活性的功能[30]。
3 離子穩態
植物暴露在高鹽環境中時,除了面臨上述滲透脅迫導致的攝水困難外,還面臨著如Na+積累造成的離子毒害。植物通過離子轉運蛋白將體內多余的Na+轉出或區域化,模式植物中關于這類蛋白的研究較多。
3.1 NHX型Na+/H+反轉運蛋白與Na+區域化
NHX蛋白(NHX Na+/H+antiporters)是離子區域化的重要轉運器,所有已測序的植物基因組都含有NHX轉運蛋白。擬南芥中有8種NHX亞型(AtNHX1-AtNHX8),其中4種亞型(AtNHX1-AtNHX4)定位于液泡膜,2種亞型(AtNHX5和AtNHX6)定位于核內體,其余2種亞型(AtNHX7/SOS1和AtNHX8)定位于質膜[31-32]。其中一些液泡NHX的過表達和異種表達導致植物的耐鹽性增強[33]。AtNHX1蛋白C端以Ca2+依賴的方式與液泡內的鈣調蛋白(CaM)相互作用,這種相互作用抑制AtNHX1的轉運活性,說明離子區域化受Ca2+信號調控[34]。另外,NHX還參與維持植物體內K+穩態[31]。該通路調控模式如圖2。
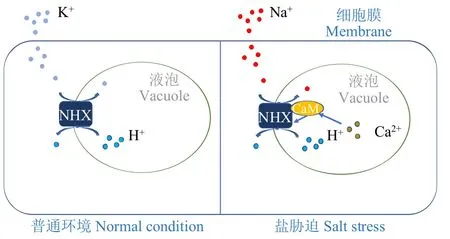
圖2 離子區域化示意圖Fig.2 Schematic diagram of ionic separation
3.2 SOS通路與Na+轉運
高等植物體內存在一個高度保守的鹽過敏感(salt overly sensitive,SOS)通路,SOS通路主要由SOS1、SOS2、SOS3和類SOS3蛋白(SCaBP8)組成。SOS1是植物質膜上的Na+/H+反轉運器,主要負責將細胞內的Na+外排[35]。SOS2是一種絲氨酸/蘇氨酸激酶,該酶會被鹽脅迫引起的Ca2+信號激活[36]。SOS3/SCaBP8 是 Ca2+接收器[37-38]。SOS1在正常情況下處于自抑制狀態[39],ABI2、14-3-3、GI和SOS2類似蛋白PKS5,作用于SOS2并抑制其活性[40]。鹽脅迫下,胞外鹽離子結合到植物細胞質膜外側GIPC,引起細胞表面電勢變化,打開質膜的Ca2+通道,細胞內Ca2+濃度增加形成Ca2+信號[8],14-3-3蛋白解碼鹽誘導的Ca2+信號后與PKS5相互作用抑制其活性[41],SOS2蛋白被釋放,SOS3及SCaBP8結合Ca2+并與SOS2相互作用,增強SOS2活性并將其帶到質膜,激活SOS1,促進Na+外排[37-38]。
Li等[40]發現糖原合成酶激酶類似物(BIN2)在鹽脅迫結束后恢復期對于SOS通路的關閉起到關鍵作用,BIN2在鹽脅迫過程中抑制BZR1/BES1等轉錄因子,從而抑制植物生長,在脅迫后恢復階段BIN2與SOS3/SCaBP8形成復合物共同抑制SOS2活性并釋放BZR1/BES1,使植物恢復生長。
3.3 HKT型轉運器
HKT型轉運器在植物Na+轉導中的重要性最早在擬南芥上報道,研究[42]表明,擬南芥AtHKT1參與根系中Na+內流,AtHKT1通過韌皮部將莖、葉、花中大量Na+轉運到根部以減少Na+在地上部的積累。其他作物如水稻有2類7種HKT轉運器,第1類為Na+轉運器,第2類則為Na+/K+通用離子轉運器,在鹽脅迫下離子穩態中都有重要作用[42]。
4 活性氧及抗氧化系統
4.1 活性氧
ROS是一種信號分子,參與代謝、抗病、細胞凋亡和消耗過剩光能等過程。遭遇非生物脅迫時,植物光能利用和碳同化受抑制,光合鏈中電子傳遞給O2的幾率變大,在線粒體、葉綠體和過氧化酶體中產生大量的ROS,包括超氧陰離子(O2-)、羥基自由基(-OH),單線態氧(1O2)和過氧化氫(H2O2)。O2-造成脂質過氧化,危害細胞膜,還能氧化組氨酸、甲硫氨酸和色氨酸等特定氨基酸,在細胞中富集的O2-還會通過梅勒反應生成更多有害的ROS[43];H2O2在植物細胞中大量積累會氧化一些酶的硫醇基,使酶失活[44];-OH是ROS中最活躍、危害最大的物質,它對其生成點上的生物分子有很高的親和力,氧化反應幾乎與擴散同步。
ROS不僅損害質膜和光系統,甚至參與植物某些基因的表達,引起DNA剪切、損傷和修飾,致使蛋白質合成減緩或降解,在細胞水平和分子水平產生不可逆的損傷,導致細胞死亡[45-46]。植物為了抵抗ROS的毒害作用,進化出一套包括超氧化物歧化酶(superoxide dismutase,SOD)、過氧化氫酶(catalases,CAT)、抗壞血酸過氧化物酶(ascorbate peroxidase,APX)、谷胱甘肽過氧化物酶(glutathione peroxidase,GPX)和谷胱甘肽還原酶(glutathione reductase,GR)等抗氧化酶系統,以及依賴谷胱甘肽(glutatbione,GSH)、脫落酸(abscisic acid,ABA)、生育酚等非酶抗氧化劑的非酶抗氧化系統[47],共同參與植物體內多余ROS的清除。
4.2 抗氧化系統
SOD是細胞內最有效的透明質酸抗氧化劑,廣泛存在于所有易受ROS侵害的細胞中。SOD在植物抗逆中的主要功能是降低體內O2-含量,它為植物抵抗高水平ROS的毒害提供了第1道防線,SOD因其輔酶因子的不同分為3類:銅/鋅-超氧化物歧化酶(Cu/Zn-SOD)、錳-超氧化物歧化酶(Mn-SOD)和鐵-超氧化物歧化酶(Fe-SOD)[48]。
CAT將H2O2還原為水和O2,CAT是所有酶中轉化效率最高的酶類,每分鐘大約可以將600萬個H2O2分子轉化成水和O2。CAT可以抵消鹽脅迫的不利影響,提高葉綠素熒光參數和光合色素含量,避免葉綠體聚集[49]。
APX被認為是高等植物中清除活性氧并保護細胞最重要的酶。APX家族至少由4種不同的亞型組成,包括類囊體形式(tAPX)、乙醛酸循環體膜形式(gmAPX)、葉綠體基質可溶性形式(sAPX)和胞質形式(cAPX)[50]。鹽脅迫下APX調節是一種后發性反應,鹽處理下蘿卜葉片和根中APX活性均增強,并且這種增強是在鹽脅迫幾天后才變得明顯[51]。
GPX是機體內廣泛存在的一種重要的過氧化物分解酶。GPX的活性中心是硒半胱氨酸,其活力大小反映機體硒水平。硒是GPX酶系的組成成分,它能催化GSH變為氧化型谷胱甘肽(GSSG),使有毒的過氧化物還原成無毒的羥基化合物,從而保護細胞膜的結構和功能。
GR是一種黃素蛋白氧化還原酶,存在于原核生物和真核生物中。GR是抗壞血酸-谷胱甘肽(ASA-GSH)循環的潛在酶,通過維持GSH低含量,在ROS防御系統中發揮重要作用。GR主要分布于葉綠體,少量分布于線粒體和細胞質[52-53]。
單脫氫抗壞血酸還原酶(monodehydroascorbate reductase,MDHAR)和脫氫抗壞血酸還原酶(dehydroascorbate reductase,DHAR)也是植物體內重要的活性氧清除酶體。
抗氧化酶功能和轉基因工作一直在不斷深化,表1列出了抗氧化酶還原活性氧的反應式,表2為抗氧化酶及其在轉基因植物抗鹽脅迫中的作用。
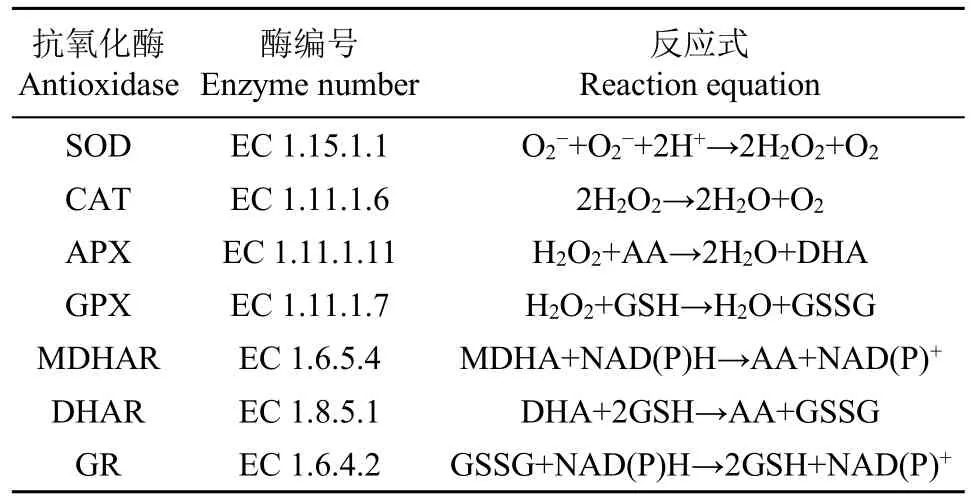
表1 抗氧化酶及抗氧化反應式Table 1 Antioxidant enzymes and antioxidant reaction equation
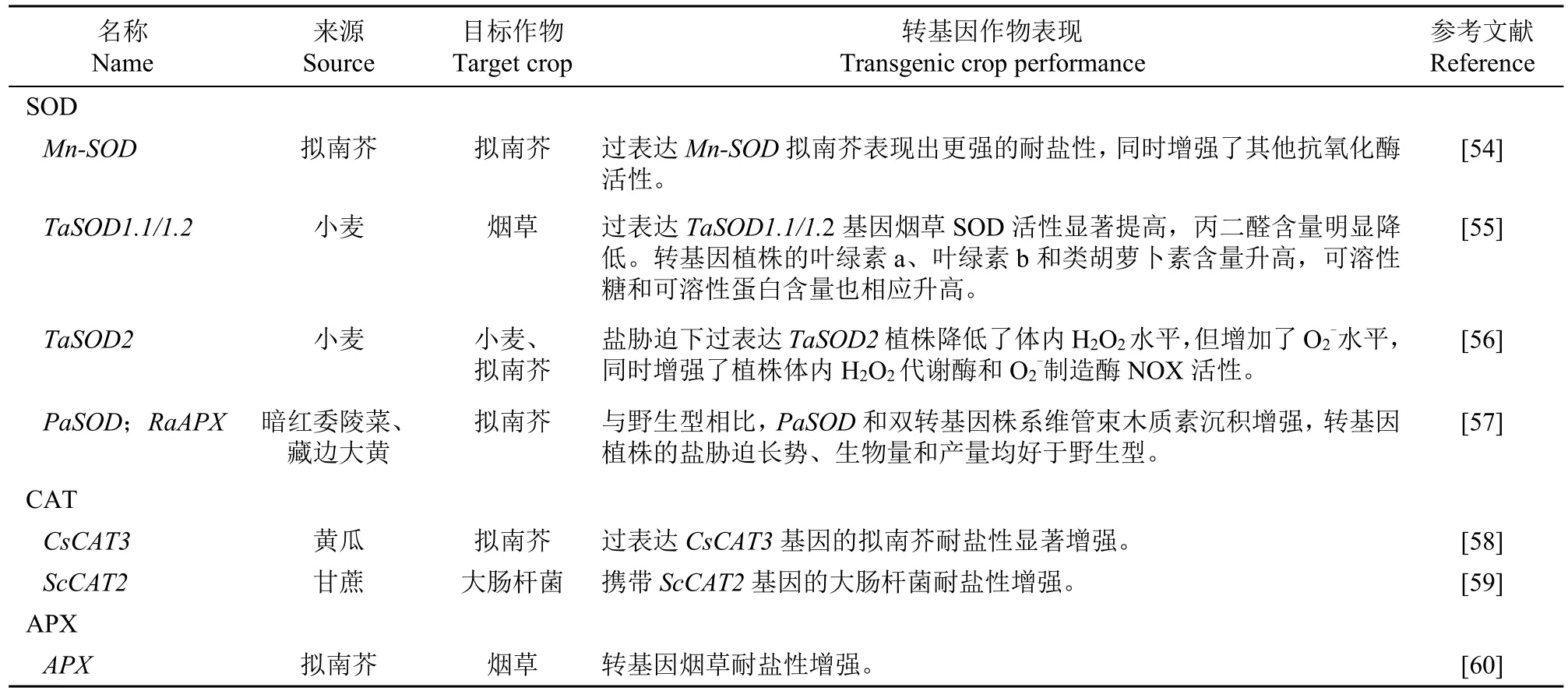
表2 活性氧清除酶及其在轉基因植物抗鹽脅迫中的作用Table 2 Reactive oxygen scavenging enzymes and their roles in salt resistance of transgenic plants
續表2 Table 2(continued)
4.3 非酶抗氧化劑
抗壞血酸(ascorbic acid,ASA)被認為是最有效的活性氧清除物質,它能夠通過將酚氧(tocopheroxyl)還原為α-生育酚(α-tocopherol)的方式直接清除-OH和O2-,保護細胞膜[47]。ASA的功能在許多作物中都有報道,如小麥[73]和葫蘆巴[74]的研究發現,外源ASA增加鹽脅迫下植株的發芽率和幼苗長度,促進抗氧化系統,提升光合作用能力。最近一項研究[75]表明,ASA促進鹽脅迫早期Ca2+積累并參與激活SOS通路。
Pro不僅是一種滲透保護劑,也是重要的非酶抗氧化劑,微生物、動物和植物都需要在體內合成Pro以減輕ROS的不利影響。Pro可以作為一個能量庫來調節氧化還原電位[76],或作為一種溶質保護大分子不變性[77]。
GSH在植物受到鹽脅迫時維持細胞正常還原狀態,抵消ROS誘導的氧化脅迫[47]。關于大麥的研究[78]表明,外源GSH處理可明顯提高大麥抗氧化酶活性以及抗氧化劑含量,降低細胞膜脂過氧化水平。
α-生育酚(α-tocopherols,vitamin E)是一種低分子量親脂性抗氧化劑,它使細胞膜免受氧化和脂質過氧化損傷,提高細胞膜的穩定性和透性,還有助于為光合作用器官提供最佳環境[79]。鹽脅迫下,外源生育酚保護光合作用器,恢復植株體內重要營養物質的平衡,最大限度保障蠶豆的生長和產量[80]。
黃酮類(flavinoids)化合物最突出的功能是賦予水果顏色,吸引種子傳播者。花青素(anthocyanins)是植物體內常見的黃酮類天然色素,在植物的抗氧化脅迫中有重要作用[81],其主要功能為:(1)攔截和吸收光能,避免非生物脅迫下光能過剩對植物葉片造成光損傷;(2)保護抗氧化酶;(3)清除自由基;(4)與其他信號通路中的分子相互作用,間接清除ROS[82]。Oosten等[83]在salt overly sensitive3-1(sos3-1)突變體的第2位點抑制子的篩選中,分離出了anthocyaninimpaired-response-1(air1)突變體。在鹽脅迫下,air1無法積累花青素,這是sos3-1在高NaCl水平(120mmol/L)下的關鍵表型。air1突變體在鹽脅迫下花青素產生缺陷,而在其他脅迫如強光、低磷、高溫或干旱脅迫下則沒有。這一特異性表明,air1突變并不影響花青素的生物合成,而是影響其對鹽脅迫的調控。該研究說明,鹽脅迫下花青素的積累可能受SOS通路調控,此研究為非生物脅迫與黃酮類花青素形式的抗氧化劑積累之間的關系開辟了新的道路。
5 植物激素與耐鹽調控
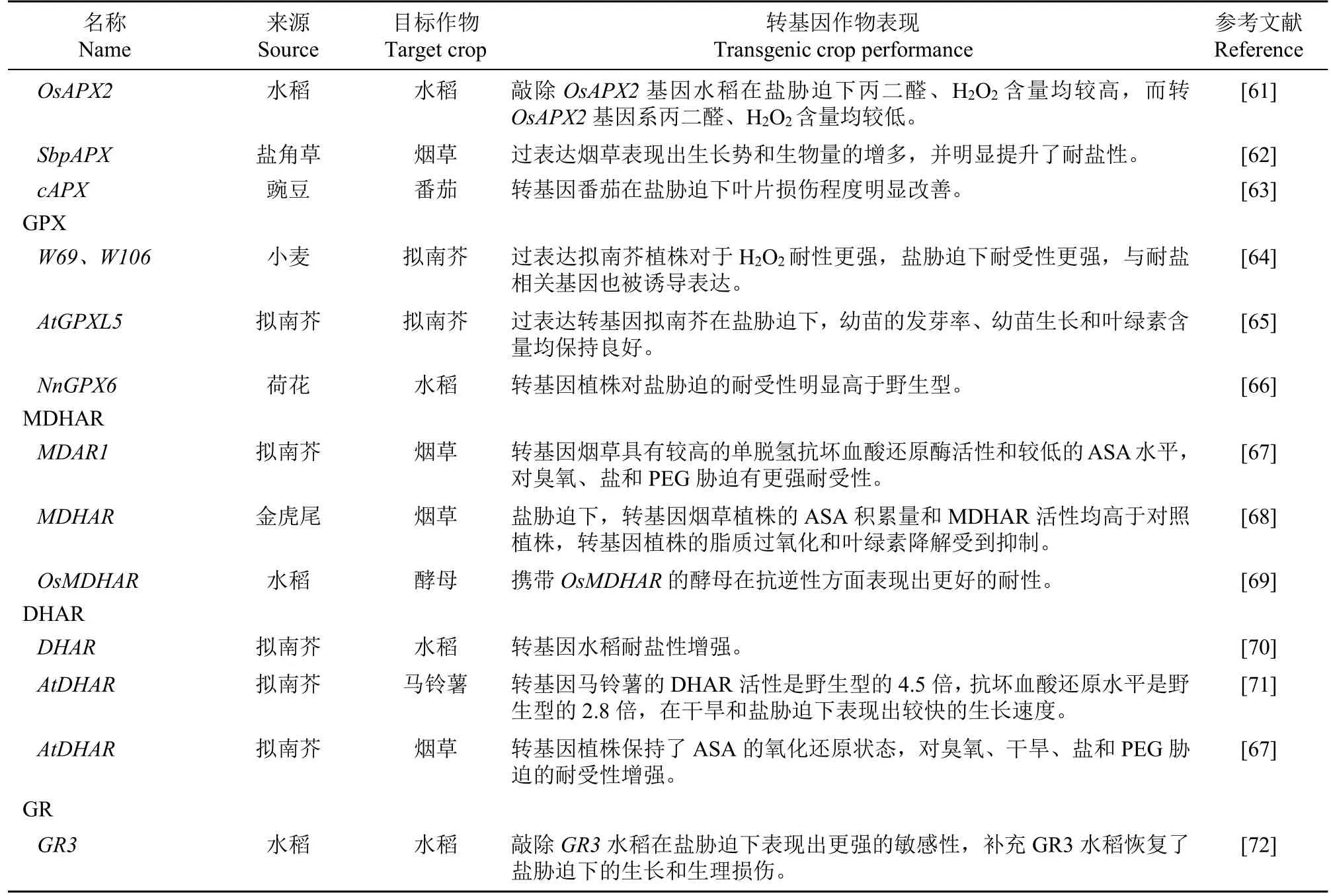
ABA是重要的抗逆激素。經典的ABA信號通路認為,正常生長條件下,Ca2+和ABI1/PP2C(abscisic acid insensitive 1/protein phosphatase 2C)抑制SnRK2/3/6/7/8/CDPK(蔗糖非酵解型蛋白激酶/鈣依賴型蛋白激酶,sucrose non-fermenting 1-related protein kinases/calcium dependent protein kinase)活性,繼而抑制下游信號[84-85]。非生物脅迫導致ABA水平升高,此時ABA信號被ABA受體PYR/PYL(pyrabactin resistance/pyrabactin resistancelike)感知并抑制ABI1/PP2C活性,誘導RCARs和H2O2水平升高,進而通過SnRKs2/3/6/7/8/CDPK將下游靶標磷酸化實現滲透調節,目前已經發現了幾種SnRK2底物,包括AREB/ABF家族轉錄因子、RAV1(與ABI3/VP1相關)和離子通道(如SLAC1),這些基因是啟動ABA下游響應基因和氣孔運動的關鍵組分[86]。此外,在啟動子中含有其他脅迫響應順式調控元件的轉錄因子家族可能參與介導ABA響應基因的表達,已經鑒定的轉錄因子包括 NAC-、MYB-、HD-Zip、AP2/ERF-和 WRKY-型轉錄因子,它們在不同植物中被誘導響應鹽脅迫[87]。
研究[88-89]表明,ABA通路可能與Ca2+信號通路整合,觸發細胞ABA水平的應激信號也可以在植物中調用顯著的細胞Ca2+信號,鈣調磷酸酶b樣蛋白(CBLs)/CBL-相互作用蛋白激酶(CIPKs)可感知這些信號,引發下游靶基因響應。ABA通路模型如圖3。
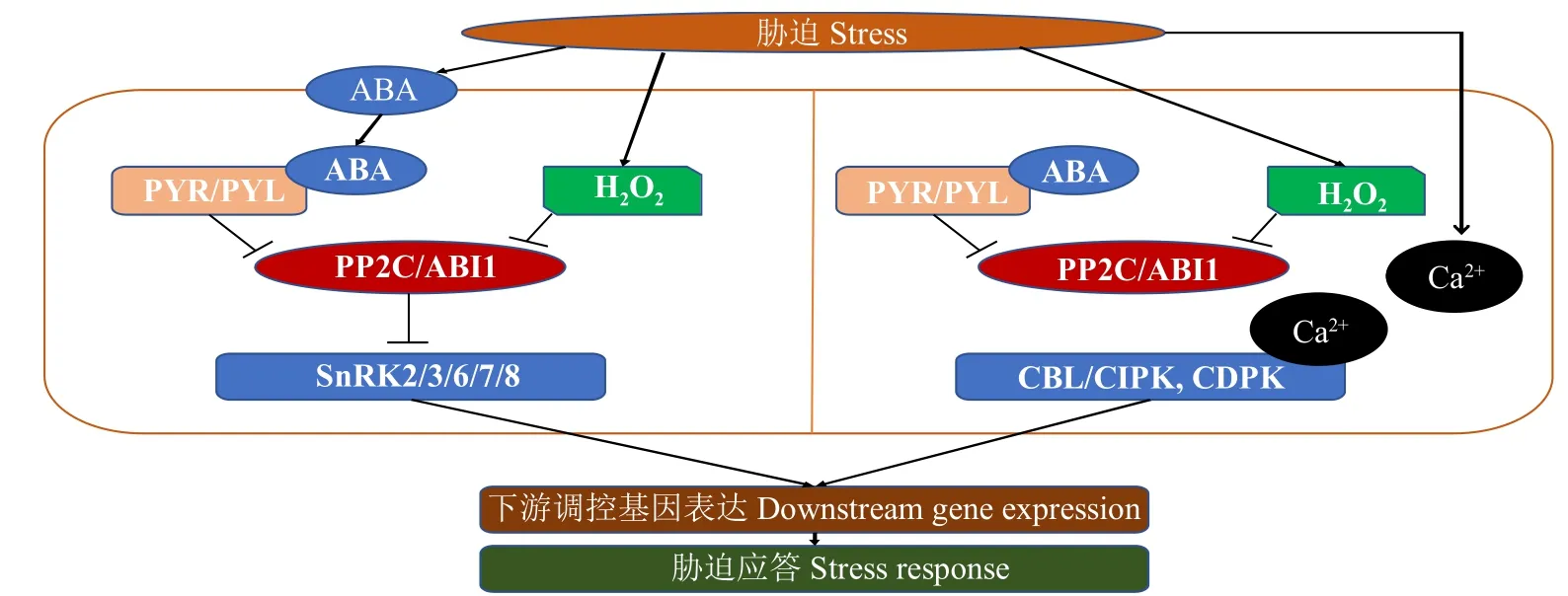
圖3 ABA通路示意圖Fig.3 Schematic diagram ofABApathway
油菜素內酯(brassinosteroid,BR)是植物中廣泛存在的激素,在細胞分化、植物生長和非生物脅迫應答中有關鍵作用[90]。棉花葉片和根系上施用BR降低了鹽脅迫下棉花幼苗根、莖、葉中Na+、Cl-和丙二醛含量,提高Pro、葉綠素含量,提高棉花根系活力和總生物量[91]。BR對ROS代謝的調節作用也是明顯的,它們可以誘導和調節某些抗氧化基因的表達,增加關鍵抗氧化酶的活性[92]。最近的一項研究[93]發現,鹽脅迫早期有限度的BR信號激活或OsGSR1(BR合成調控基因)的正常表達,在低濃度ABA介導的鹽脅迫耐受性中發揮關鍵作用,這項研究揭示了BR與ABA共同調控植物耐鹽的分子機制。
乙烯是最常見的植物逆境調節激素之一,主要作用包括解除植物種子休眠,刺激植物莖、根的生長和分布,促進植物葉片和果實的脫落,影響雌雄異花植物雌性花的分化,抑制植物花的開放,促進花和葉片的衰老及果實成熟。鹽脅迫下,乙烯信號元件EIN3/EIL1激活ROS清除基因表達,抑制過量ROS積累,增強擬南芥的耐鹽性,也有研究[94-95]表明,乙烯信號的阻斷會降低擬南芥的耐鹽性。鹽脅迫下乙烯維持芽和根中K+促進離子穩態,增加擬南芥鹽脅迫能力[96]。該激素在鹽脅迫中的信號轉導模式已經被很好地總結[97]。
葉面噴施茉莉酸(jasmonic acid,JA)和水楊酸(salicylic acid,SA)的大豆細胞中Na+含量降低,K+和Ca2+含量上升,植株中甜菜堿、可溶性蛋白和葉綠素含量升高,抗氧化酶活性上調,細胞膜的穩定性和葉片含水量上升,并最終提高大豆產量[98]。
在擬南芥中發現了植物響應鹽脅迫的新通路,該通路整合了JA、SA和ABA在植物響應鹽脅迫的作用[99]。該通路由細胞壁富含亮氨酸重復伸展蛋白(LRX3/4/5)、快速堿化因子RALF22/23和受體激酶(FER)組成。正常情況下,LRX3/4/5蛋白與細胞壁中的RALF22/23結合,阻止RALF22/23與質膜定位的FER結合,促進植物生長,抑制脅迫反應。在鹽脅迫下,LRX3/4/5和RALF22/23通過一種尚未明確的機制被分離。釋放的RALF22/23肽結合FER并誘導其內化,暫停植物生長,FER的內化也會激活脅迫應激反應。另外,細胞外LRX3/4/5-RALF22/23-FER負調控JA、SA和ABA的水平[100],且這些基因的突變體植株表現出對鹽脅迫的超敏感性與ROS的積累,在突變體中阻斷這些通路或ROS的積累可以一定程度緩解鹽的超敏感反應,說明LRX3/4/5-RALF22/23-FER通過調節激素穩態和ROS積累來控制植物的鹽脅迫反應。該通路調控模式如圖4。
圖4 LRX3/4/5-RALF22/23-FER調控通路Fig.4 LRX3/4/5-RALF22/23-FER regulatory pathway
6 其他方式
Zhu等[101]研究表明,添加硅(Si)顯著降低了鹽脅迫下2個黃瓜品種葉片中電解質泄漏比例、H2O2和巴比妥酸反應物質的含量,顯著提高了抗氧化酶的活性,并且這種效應隨試驗的進行而增強。Zhang等[102]研究了外源Si對盆栽甘草鹽脅迫的影響,結果發現外源Si一定程度上恢復了甘草的生長,植株體內可溶性糖和蛋白含量顯著增加,攝水困難癥狀得到緩解,并且體內抗氧化酶活性顯著提高,細胞膜脂質過氧化程度減弱,膜透性增強。
硒(Se)在植物應對鹽脅迫時也有重要作用。鹽脅迫下,外源施加Se顯著增加玉米生物量、根表面積和體積,提高植株內可溶性糖含量,降低葉片中 K+、Na+和 O2-含量[103]。
多胺類(polyamines,PA)是一種低分子量、普遍存在于植物界的脂族分子。在植物生長發育過程中,多胺起著調節細胞增殖、體細胞胚胎發生、分化和形態發生,打破塊莖和種子休眠,促進花果發育和衰老的作用。多胺在植物鹽脅迫中的積極作用體現在維持膜的完整性、調控抗氧化酶系統、減少葉綠素損失、降低ROS含量、維持植物組織和細胞中的Na+/K+、保護植物組織的完整和功能、減少地上部分Na+和Cl-含量等[104]。
一氧化氮(NO)是一種氣體小分子,參與調控植物的各種生長發育過程,如根系生長、呼吸、氣孔關閉、開花、細胞死亡、種子萌發等,同時也是一種脅迫信號分子。NO直接或間接觸發許多氧化還原調節基因的表達。NO可以與脂質自由基反應以減輕脂質過氧化,清除超氧化物自由基和其他細胞過程生成的過氧亞硝基來保護細胞。它還有助于激活SOD、CAT和GPX等抗氧化酶[105]。NO會激活H+-ATPase,建立H+梯度,進而為Na+/H+離子反轉運器提供動力,維護植物體內Na+/K+[106]。200mmol/L NaCl會抑制羽扇豆發芽,但是經過NO供體(sodium nitroprusside,SNP)預處理后,這種抑制作用會被抵消[107]。
微生物對植物耐鹽脅迫也有促進作用,如耐鹽性強的緩生根瘤菌可以促進柱花草在鹽脅迫下的生長[108]。某些植物生長促進菌(plant growth promoting bacteria,PGPB)對促進植物耐鹽性和生長具有特殊機制。這些細菌誘導植物合成植物生長素、細胞分裂素和赤霉素以及揮發性有機化合物。這些細菌也產生生長調節劑,如具有固定氮、溶解有機和無機磷酸鹽的載體鐵[109]。
7 問題與展望
鹽脅迫是影響作物生長發育的最主要的不利環境因素之一,它通過水分脅迫、氧化脅迫、營養失衡、離子毒害、膜紊亂、細胞分裂伸長的抑制、關鍵代謝過程的中斷以及遺傳毒性影響植物發芽、生長、生活力和產量。幾十年來,許多關于植物耐鹽機理的關鍵基因和信號通路被提出,涉及鹽脅迫信號的感知和轉導,以及體內離子和ROS穩態的維持。但對植物脅迫應答網絡的認知仍然十分有限。
第一,植物細胞最開始如何感知鹽(Na+輸入)的機制一直沒有明確,前文提到2種可能的感知方式,一是尋找Na+感受器,通過篩選Ca2+瞬態,已經有少數候選的應激感受器,今后研究的重點應放在這些感受器的下游基因和調控網絡。二是關于質膜和細胞壁界面的壓力信號,LRX-FALF-FER通路已經被確定,但是LRX如何感知來自細胞壁的脅迫信號還不清楚。該通路除了通過3種植物激素調節耐鹽性外還有沒有其他的作用,是后續研究中應著重解決的問題。
第二,植物不同細胞和組織如何響應脅迫,不同細胞和組織的響應如何通過細胞間通訊和遠距離信號協調和整合,目前尚不清楚[110]。具體表現如前文提到的Na+會在植物不同部位積累或者外排,什么因素致使植物各器官對鹽的不同敏感性,敏感性是否依賴于不同的鹽積累或組織特異性的信號傳導,這些問題的答案會為提高植物耐鹽性起到關鍵作用。今后的研究中,將離子轉運相關基因的表達調控建立在特定組織或特定細胞水平上或許可以回答這些問題。
第三,植物耐鹽脅迫應答機制在擬南芥的研究中已日漸清晰,但作物抗逆性分子機制的系統研究仍然滯后。與擬南芥相比,其他作物往往更復雜,可能有額外的機制來應對鹽脅迫,不同的物種可能具有不同的應激反應機制。隨著基因組測序、組裝和注釋技術的發展,種質資源的收集和分析,大量作物中耐鹽脅迫基因甚至新的調控機制會在不久的將來被發現。此外,鹽生植物的研究略顯不足,過表達蘇打豬毛菜NHX基因的紫花苜蓿可以在高達400mmol/L NaCl的環境中生活50d[111],鹽生植物特殊的耐鹽基因和分子機制可能對未來耐鹽作物育種提供基因資源和理論基礎。
第四,現有的研究多集中在植物本身對鹽脅迫的感知與應答,但植物所處的環境空間中的其他因素如光照、溫度、濕度和微生物等,這些因素之間對于植物耐鹽性是否有影響,需要更多深入的研究。
第五,盡管有很好的研究前景和大量的信息積累,通過轉基因手段開發既耐鹽又具有優良農藝性狀(如產量)的作物依然困難,沒有增加耐鹽性的轉基因作物商業化[112]。因為植物耐鹽性往往與體內離子穩態直接相關,離子穩態的改變往往會導致產量的降低[110]。一些基于田間的研究如轉HAL1基因番茄T-6在鹽脅迫下的產量比原品系提高了27%[113],在250mmol/L NaCl下,轉P5CS基因的鷹嘴豆在種子質量和數量上均沒有明顯下降[114],這種在商業水平開發耐鹽作物值得嘗試。除傳統育種和基因工程育種外,一些研究有望在未來生產實踐中應用,低鹽馴化或在基質中混入生物炭均能有效地提高作物耐鹽性[115-116]。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
