特基拉芽孢桿菌液態發酵制備菜籽多肽的工藝研究
2022-11-24 04:56:40鐘曉霞劉志云周曉容黃金秀
中國糧油學報 2022年10期
鐘曉霞, 劉志云,2, 周曉容,2, 黃金秀,2
(重慶市畜牧科學院1,重慶 402460) (農業部養豬科學重點實驗室2,重慶 402460)
菜籽粕是油菜籽榨油之后的副產物,其粗蛋白質量分數可達35%~45%,富含礦物質元素、維生素和氨基酸,是一種優質的植物蛋白資源[1]。但菜籽在熱榨或溶劑提取制油過程中,菜籽蛋白嚴重變性,導致消化吸收率低[2],制約了菜籽粕的深度利用,造成蛋白資源的浪費。研究表明,菜籽蛋白經蛋白酶作用,可得到許多分子鏈長度不等的、易溶解、易吸收的低分子小肽混合物[3],且具有抗氧化[4]、提高免疫力[5]、降血壓[6]、抗腫瘤[7]等功能活性。近年來,研究人員發現通過微生物發酵菜籽蛋白可獲得功能性菜籽多肽,而且與酶水解法制備相比,可以節省大量昂貴的酶制劑,降低生產成本[8],已成為菜籽粕高值化利用的新途徑之一。何榮海等[9]以枯草芽孢桿菌液態發酵菜籽粕得到具有抗氧化活性的菜籽肽,金晶[10]以枯草芽孢桿菌液態發酵菜籽粕得到具有ACE抑制活性的菜籽肽。
本研究實驗室從土壤中分離篩選到一株產堿性蛋白酶和中性蛋白酶的特基拉芽孢桿菌(Bacillustequilensis),能夠在菜粕平板上良好生長,具有分解菜籽蛋白的潛力。影響菜粕發酵獲得菜籽肽的因素包括發酵基質的pH值、料水比、KH2PO4添加量、葡萄糖添加量[11];發酵的溫度、時間、菌種接種量、轉速等[9]。由于菌株的特異性,不同菌株發揮作用所需條件不盡相同[12]。因此本研究擬以篩選到的特基拉芽孢桿菌為發酵菌株,以多肽得率為篩選條件,采用單因素和正交分析法對發酵基質的組成和發酵工藝進行優化,獲得最佳發酵基質組成和發酵工藝,提高菜籽多肽得率。以期為菜籽粕的深度利用提供參考。
1 材料與方法
1.1 材料
原料與試劑:菜籽粕,粗蛋白質量分數36.89%,含水量7.54%;2,2-二苯甲基苦味酰基苯肼基(DPPH)、2,4,6-三吡啶基三嗪(TPTZ)、2,2′-聯氮-雙-3-乙基苯并噻唑啉-6-磺酸(ATBS),分析純;Gly-Gly-Tyr-Arg,色譜純。
營養瓊脂培養基:牛肉膏3 g,蛋白胨10 g,NaCl 5 g,蒸餾水1 000 mL,瓊脂20 g,pH 7.2,121 ℃滅菌20 min,不加瓊脂作液體培養基使用。
1.2 實驗方法
1.2.1 特基拉芽孢桿菌液態發酵制備菜籽多肽的工藝優化
初始發酵基質:準確稱取2 g過60目菜籽餅(發酵底物),加入30 mL蒸餾水,121 ℃高溫高壓21 min。冷卻至室溫,備用。發酵條件為0.25 mL/g發酵底物的接種量,37 ℃,150 r/min發酵36 h。
1.2.1.1 發酵基質的優化
通過單因素實驗考察發酵料水比(1∶10、1∶15、1∶20、1∶25),KH2PO4添加量(0、1、3,5、7、9 mg/mL),葡萄糖添加量(0、1、3、5、7、9 mg/mL),發酵初始pH(4、5、6、7、8、9、10)對菜籽多肽得率的影響,考察某一單因素時,其他發酵條件不變。
1.2.1.2 發酵條件的優化
在發酵基質優化的基礎上,通過單因素實驗考察轉速(120、150、180、210、240 r/min),接種量(0.05、0.15、0.25、0.35、0.45 mL/g發酵底物),發酵溫度(33、35、37、39、41 ℃),發酵時間(12、24、36、48、60 h)對多肽得率的影響,考察某一單因素時,其他發酵條件不變。選取各因素的最優實驗范圍,采用正交分析法進行優化。
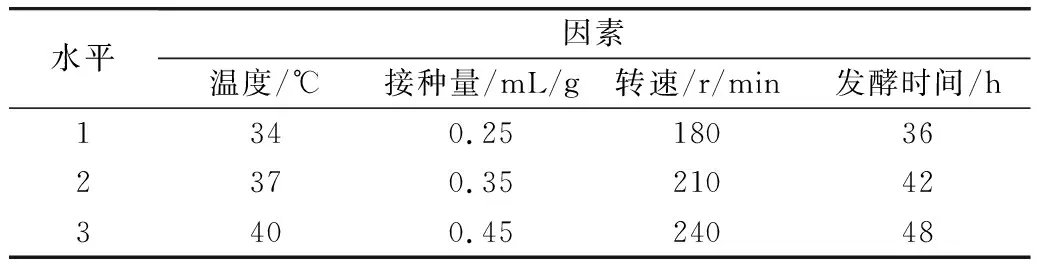
表1 L9(43)正交實驗設計
1.2.2 最優條件下發酵制備菜籽肽
在最優的發酵條件下開展驗證試驗,以未接種菌液在相同條件下發酵的樣品作為對照組,每組設置4個重復。發酵結束后檢測多肽得率、發酵上清液的氨基酸含量和體外氧化活性。
1.2.3 發酵液處理
將培養好的發酵產物取出,2~4 ℃條件下8 000 r/min離心10 min,上清液定容至50 mL,用0.45 μm微孔濾膜過濾上清液,除去不溶物和細菌,備用。
1.2.4 多肽得率的測定
根據所報道的文獻[13],配置試劑。多肽標準曲線的建立:將Gly-Gly-Tyr-Arg用5% TCA分別配制成0、0.312 5、0.625、1.25、2.5和5 mg/mL濃度的標準溶液,取0.9 mL不同稀釋梯度的溶液置于5 mL離心管中,加入雙縮脲試劑3.6 mL,混勻,靜置10 min,2 000 r/min離心10 min,于波長540 nm處測定OD值。在0~5 mg/mL的濃度范圍內,多肽的標準曲線為Y=19.042x+0.021 9(x為吸光值,Y為多肽濃度,mg/mL),R2=0.999 9,線性關系良好。
取0.75 mL定容后的發酵液,加入0.75 mL 10%的三氯乙酸水溶液,旋渦混勻,靜置30 min,然后在10 000 r/min離心10 min。將上清液全部轉移到10 mL比色管中,用5%三氯乙酸定容至10 mL,搖勻,取0.9 mL上述溶液置于5 mL離心管中,加入雙縮脲試劑3.6 mL,混勻,靜置10 min,2 000 r/min離心10 min,取上清液于540 nm下檢測OD值。
多肽得率=(待測液中肽含量×發酵液定容體積)/發酵底物總質量×100%
1.2.5 菜籽多肽體外抗氧化活性研究1.2.5.1 DPPH清除率的測定
根據所報道的文獻[14, 15],用無水乙醇將DPPH配制成0.12 mmol/L的溶液,并避光保存。將待測樣品配制成不同梯度濃度的溶液;分別與DPPH溶液按1∶1的比例混合均勻,放置于常溫下避光反應30 min。最后,反應結束后在 517 nm下檢測樣品的吸光值。實驗中同時還需要設置空白組和對照組,每組設置3個平行。
DPPH清除率=(1-(AS-AC)/A0)×100%
式中:As表示樣品組(樣品和DPPH)吸光值;Ac表示對照組(樣品和無水乙醇)吸光值;A0表示空白組(DPPH和樣品溶解液)吸光值。
1.2.5.2 ABTS 自由基清除活性測定
根據所報道的文獻[16, 17],室溫下,在黑暗環境中用2.45 mmol/L的K2S2O8水溶液與7.0 mmol/L的 ABTS水溶液反應12~16 h產生ABTS自由基溶液。使用前,用pH 7.4的0.2 mol/L PBS 緩沖液稀釋 ABTS母液,使其在波長734 nm處的吸光值為 0.7±0.02。移取50 μL不同濃度的稀釋樣品與150 μL ABTS 溶液于96孔酶標板上混勻,室溫下避光反應6 min后,于波長734 nm處測定吸光值
ABTS自由基清除率=[1-(A1-A3)/(A2-A4)]×100%
式中:A1表示樣品組(樣品和ATBS);A2表示控制組(超純水);A3表示樣品空白組(樣品+PBS溶液);A4為空白組(超純水+PBS)。
1.2.5.3 鐵還原抗氧化能力(FRAP)測定
根據所報道的文獻[18],配置FRAP檢測試劑,標準曲線建立:將1 mL不同濃度的FeSO4溶液加入到10 mL比色管中,加入5.0 mL FRAP 工作液,37 ℃下避光反應20 min,在593 nm處測定反應混合物的吸光度。在0~1 mmol/L的濃度范圍內FeSO4還原力標曲回歸方程為:Y=3.549 1x+0.007(x為FeSO4濃度,mmol/L;Y為吸光度),R2=0.999 9,線性關系良好。
樣品測定:按照建立標準曲線的方法進行測定。結果表示:基于標準曲線的0~1 mmol/L的濃度范圍內,以1 mmol/L的FeSO4為標準,計算FRAR值。
1.2.5.4 還原力測定
根據所報道的文獻配制試劑[19],在5 mL離心管中依次加入0.2 mL不同濃度的稀釋樣品,在依次加入1 mL 0.2 mol/L PBS磷酸緩沖液(pH 6.6)和1 mL 1%鐵氰化鉀,迅速混勻,50 ℃水浴中反應20 min,冷卻并加1 mL 10% 三氯乙酸(TCA),混勻后以3 000 r/min離心10 min。取2.5 mL上清液,加0.5 mL 0.1%三氯化鐵,混勻,測定各樣品在700 nm波長處的吸光度。用蒸餾水作為無還原能力對照。
1.2.6 數據處理
用GraphPad Prism 8作圖,SPSS 20.0軟件處理數據,以平均數±標準差(Mean±SD)表示,用T-檢驗比較分析法進行顯著性檢驗(P<0.05)。
2 結果與分析
2.1 特基拉芽孢桿菌發酵制備菜籽多肽的優化
2.1.1 發酵基質的優化
對發酵基質的料液比、pH值、KH2PO4添加量、葡萄糖添加量進行優化,由圖1所示,隨著料液比的提高,多肽得率也隨之顯著提高,在料液比為1∶25時多肽得率較高;隨著發酵基質pH的提高,發酵上清液多肽得率先升高后略微降低,在pH 8時多肽得率較高;隨著KH2PO4添加量的提高,多肽得率持續降低,對菜籽多肽得率起到反向作用,故發酵基質中不添加KH2PO4;隨著葡萄糖的添加量的提高,多肽得率先下降后提高,但相較于不添加,多肽得率提高不顯著,考慮生產成本等問題,故在發酵基質中不添加葡萄糖。
最終以料水比為1∶25,初始pH為8,不添加KH2PO4緩沖鹽和葡萄糖的發酵基質進行發酵條件的優化。
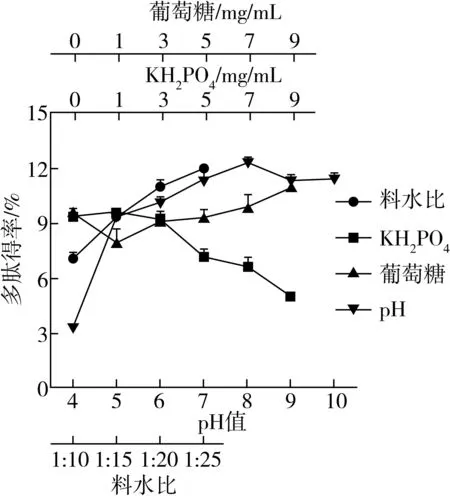
圖1 發酵基質對菜籽多肽得率的影響
2.2.2 發酵條件的優化2.2.2.1 發酵條件單因素優化
在優化發酵基質的基礎上,對菜籽粕的發酵條件進行優化,由圖2所示,隨著接種量的增加,多肽得率先增加后降低,接種量為0.35 mL/g發酵底物時,多肽得率最高;隨著溫度的增加,多肽得率先增加后降低,發酵溫度為37 ℃時,多肽得率最高;隨著轉速的增加,多肽得率增加,轉速為240 r/min時,多肽得率較高;隨著發酵時間的增加,多肽得率增加,發酵時間為48 h時,多肽得率較高。
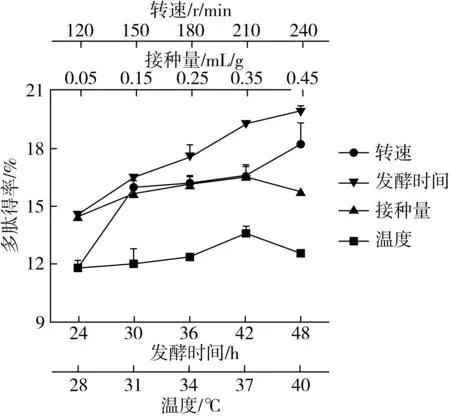
圖2 發酵條件對菜籽多肽得率的影響
2.2.2.2 發酵條件的正交優化
由表2可知,最佳發酵條件0.35 mL/g的接種量、37 ℃、240 r/min,發酵42 h。各因素對多肽得率的影響時間>溫度>轉速>接種量。
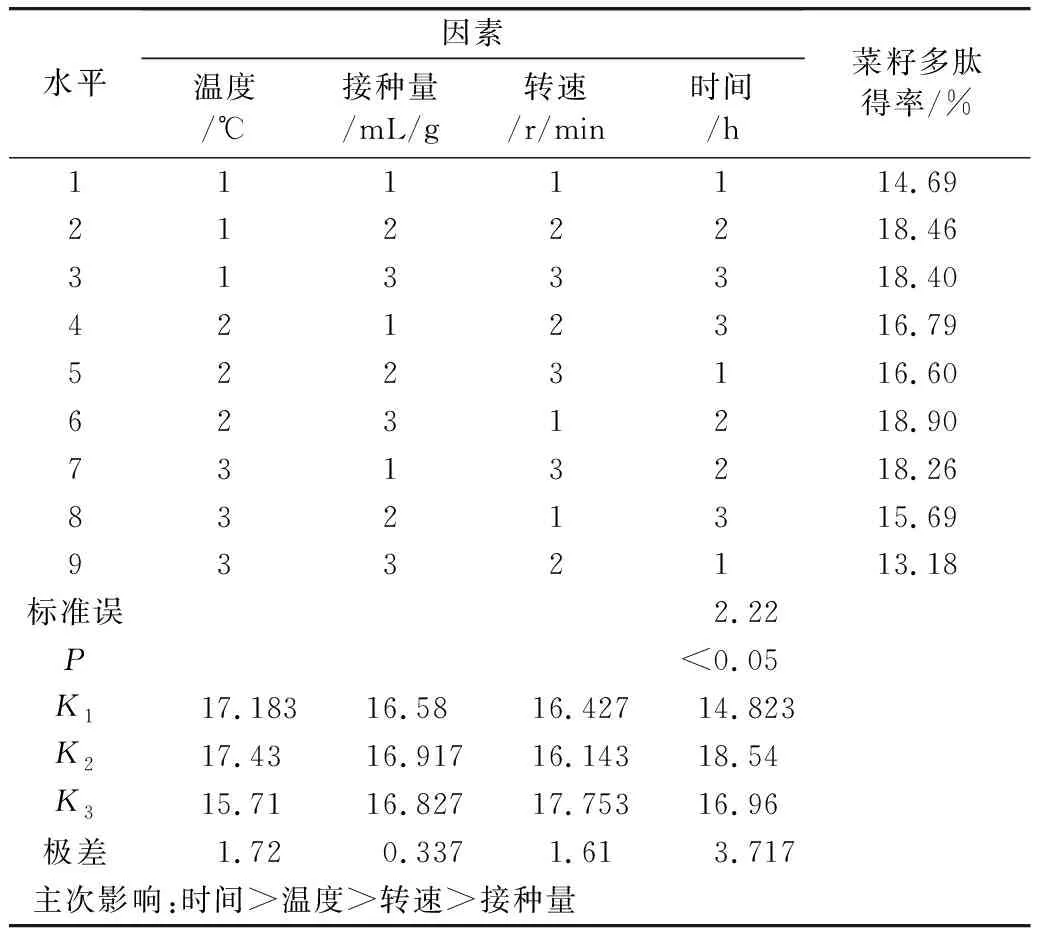
表2 發酵條件正交優化結果
2.2.3 最優條件下發酵制備菜籽肽
在最優發酵條件下進行4組平行實驗,對照組的多肽得率分別為2.9%、2.1%、2.1%、2.15%,平均值為2.3%;菜籽多肽得率分別為21.00%、22.15%、20.75%、21.5%,平均值為21.4%,較對照組提高了9.3倍。由表3可知,發酵上清液中氨基酸含量較對照組顯著提高9.55倍,必需氨基酸提高13.68倍,非必需氨基酸提高8.13倍,其中異亮氨酸、賴氨酸和組氨酸分別提高了18.24、20.46、30.98倍。
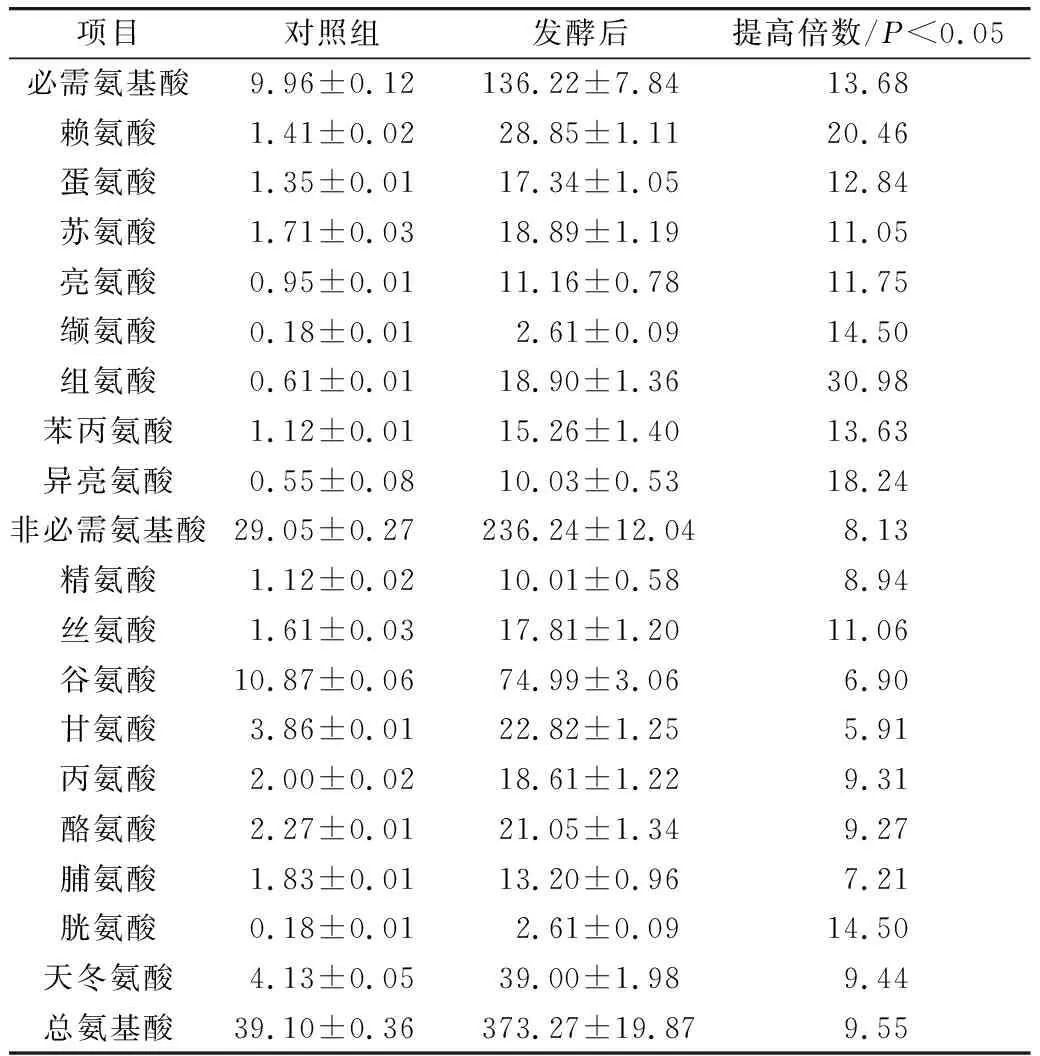
表3 發酵產物氨基酸含量/mg/100 mL
由圖3可知,在同一稀釋濃度下,發酵后上清液DPPH自由基和ATBS自由基清除活性、鐵總抗氧化能力、還原力均顯著高于對照組(P<0.05)。在試驗濃度范圍,隨著菜籽多肽濃度的增大,其抗氧化活性顯著增強,且具有一定的量效關系,經計算,菜籽多肽的50% DPPH自由基清除率為0.053 mg/mL,50% ATBS自由基清除率為0.087 mg/mL,發酵上清液稀釋20倍后的鐵總抗氧化活性為FRAR值為0.25±0.01。
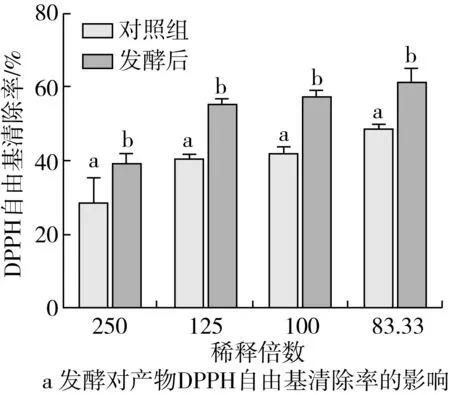
圖3 發酵對產物抗氧化活性的影響
3 討論
3.1 發酵基質對菜籽多肽得率的影響
發酵基質的料水比、KH2PO4添加量、葡萄糖添加量、初始pH均能對菜籽多肽得率產生影響。料水比決定發酵培養基中的菜籽粕添加量和含水量,在本試驗中隨著料水比的提高,菜籽多肽得率也隨之提高,這與金晶[10]的研究結果相似。合理的料水比一方面有助于菌體吸收培養基的營養物質和氧傳遞,另一方面有利于微生物代謝產物阻遏削減,促進新陳代謝,從而促進菌體的生長繁殖[20]。
KH2PO4作為微生物培養基中常用的無機鹽之一,可以提供微生物生長所需的礦物元素,緩沖發酵基質pH的快速變化,但濃度過高也會抑制菌種的生長和產酶[11]。在本實驗中,添加KH2PO4降低了菜籽多肽得率,可能是因為KH2PO4本身呈弱酸性,對于發酵產物中的乙酸等酸性代謝物的積累未起到緩解作用,從而抑制了菌株生長。發酵基質初始pH值影響菌種對環境的適應速度、生長繁殖速度、細胞內酶的活性以及蛋白質的積累[21]。在本實驗中發酵基質呈弱堿性時菜籽多肽得率較高,適宜菌株和代謝產物的生成。
葡萄糖作為易于吸收的碳源,可以在菌種發酵初期提供大量碳源,維持其生長勢頭,從而產生大量的蛋白水解酶。在本實驗中隨著葡萄糖添加量的增加先降低后提高,可能是菜籽粕自身的碳源也能滿足菌株的前期生長,再加入過多的葡萄糖一方面會加速菌體呼吸,使培養基中的溶氧不能滿足菌體的需要,另一方面加速了一些酸性中間代謝物如乳酸、丙酮酸、乙酸等的累積,導致培養基pH值降低,從而抑制細菌的生長和水解酶類的合成。
3.2 發酵條件對菜籽多肽得率的影響
接種量的大小會直接影響發酵周期和產物的累積速度,在本實驗中隨著接種量的增加,多肽得率先增加后減少。隨著接種量的提高,可以縮短菌株的延滯期,加快酶的積累;但接種量過大,細菌繁殖迅速,培養液粘度增加,溶氧不足,細胞自溶加快,反而不利于產物合成[22]。溫度影響微生物生長及產酶,過高或過低都不利于發酵過程[23]。本實驗中,多肽得率的變化與酶促反應的動力學結果相似,即隨著溫度的升高,反應速度加快,導致細菌的生長繁殖加快;但隨著溫度的上升,酶失活的速度也越大[24]。
在搖瓶中進行優化時,轉速直接影響到培養基中的溶氧量,本試驗中隨著轉速的提高,多肽得率隨之提高。可能是較高的轉速,提高了培養基中的溶氧量和菌體與營養物質的充分接觸,有利于細菌快速繁殖增長及代謝產物的合成[25]。發酵時間影響微生物的生長和酶的作用,在本實驗中,隨著發酵時間的延長,多肽得率持續提高。發酵時間太短,菌的濃度低,產酶少,不能完全發揮,而發酵時間過長,大量代謝產物的積累抑制酶的活性,并可能將多肽進一步分解為游離氨基酸,導致多肽得率降低。
在本實驗中,正交實驗的發酵時間較單因素實驗中的最佳時間有所縮短,說明各因素之間存在正向交互作用。合適的接種量,減少了菌株適應新環境的時間,適宜的溫度加速了菌株的生長繁殖;而較高的轉數提高了發酵基質中的溶氧量和氧的傳遞,為特基拉芽孢桿菌提供了更好的生長環境,也更有利于菌株產酶分解大蛋白分子成小蛋白,從而縮短了發酵時間,節約了時間成本。
3.3 最優發酵條件對菜籽多肽得率及其體外抗氧化活性的影響
通過對發酵基質和發酵條件的優化,特基拉芽孢桿菌液態發酵菜籽粕的多肽得率為21.4%,與何榮海等[9]利用枯草芽孢桿菌發酵制備菜籽多肽的得率為20.19%、金晶[10]利用枯草芽孢桿菌發酵制備菜籽多肽的得率為23.85%的研究結果相近。與對照組相比,用特基拉芽孢桿菌發酵菜粕后,發酵產物氨基酸含量大幅度提高。有研究表明,特基拉芽孢桿菌的基因能夠編碼可編碼蛋白酶、果膠酶、纖維素酶、淀粉酶等多種酶系[26],本實驗所用的特基拉芽孢桿菌能夠分泌中性蛋白酶和堿性蛋白酶,這些蛋白酶將不溶于水的菜籽大蛋白分解成可溶于水的小分子蛋白和肽,使得發酵產物中氨基酸和多肽含量大幅度提高,其中組氨酸含量提高了30.98倍。
與對照組相比,經特基拉芽孢桿菌發酵菜粕后,發酵產物的DPPH自由基清除率、ATBS自由基清除率、總抗氧化能力和還原力均顯著提高,與RONG H等[27]利用枯草芽孢桿菌固態發酵獲得菜籽肽的研究結果類似。其中50% DPPH自由基清除率所需菜籽多肽濃度低于何榮海等[9]的報道。發酵產物抗氧化活性的大幅度提高,可能與發酵產物氨基酸和多肽含量大幅提高有關。有研究表明,組氨酸、酪氨酸、甲硫氨酸和胱氨酸等具有抗氧化活性,特別是組氨酸由于其咪唑環的分解而表現出很強的自由基清除活性[28, 29]。
4 結論
特基拉芽孢桿菌液態發酵制備菜籽多肽的最佳發酵條件是發酵基質pH為8,料水比為1∶25,接種量為0.35 mL/g,發酵溫度為37 ℃,轉速為240 r/min,發酵時間為42 h。此條件下,菜籽多肽得率為21.4%,發酵上清液氨基酸含量大幅提高,體外抗氧化活性顯著提高。
