考洲洋牡蠣養殖海域海-氣界面CO2交換通量的時空變化
2022-11-26 06:00:24彭鵬飛史榮君
海洋科學 2022年10期
關鍵詞:差異
彭鵬飛, 馬 媛, 史榮君, 王 迪, 許 欣, 顏 彬
考洲洋牡蠣養殖海域海-氣界面CO2交換通量的時空變化
彭鵬飛1, 馬 媛1, 史榮君2, 王 迪1, 許 欣1, 顏 彬1
(1. 國家海洋局南海調查技術中心 自然資源部海洋環境探測技術與應用重點實驗室, 廣東 廣州 510300; 2. 中國水產科學研究院南海水產研究所 廣東省漁業生態環境重點實驗室, 廣東 廣州 510300)
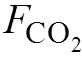
溶解無機碳;CO2; CO2交換通量; 牡蠣養殖; 考洲洋
化石燃料的使用導致大氣中以CO2為主的溫室氣體增加, 引起溫室效應加劇, 產生一系列的氣候問題[1-3]。海洋作為地球表面最大的碳庫, 其對碳的吸收能力將直接影響到全球碳循環[4], 具有巨大的碳匯潛力和負排放研發前景[5]。藍碳概念的提出, 確認了海洋在全球氣候變化和碳循環過程中至關重要的作用, 地球上大約有93 %的CO2會循環進入海洋, 它們或重新進入生物地化循環, 或被長期儲存起來[6]。1994—2007年, 海洋對CO2的年平均吸收率為2.6±0.3 PgC, 占這一時期全球人為CO2排放量的31%±4 %[7]。海洋吸收的CO2量不斷增加, 會使得表層海水的堿性下降, 引起海洋酸化, 產生另一重大環境問題[8]。因此, 一方面將CO2從大氣中移除, 減緩溫室效應, 海洋起到至關重要的作用, 另一方面, 將CO2從海水中移除, 緩解海洋酸化, 成為重要的研究方向。
大規模的貝藻類養殖能通過收獲從海洋中移出大量的碳, 明顯影響到淺海碳循環, 成為一個“可移出的碳匯”[9]。中國學者在世界上率先提出漁業碳匯和碳匯漁業的概念, 得到國際社會的廣泛關注[10-11], 貝類養殖是碳匯漁業的重要組成部分[12]。濾食性貝類具有高效的碳匯功能, 能顯著提高海域的碳匯潛力[13]。中國貝類海水養殖產生的碳沉積量為5.115×105t/a, 可減少大氣CO2增加量的0.010 9%[14]。海水養殖的濾食性貝類通過攝食活動大量去除海水中的顆粒有機碳, 并且通過形成貝殼能夠埋藏大量的碳, 伴隨著養殖貝類的收獲, 大量的碳能夠直接從海水中移出, 這勢必對養殖海區以及鄰近海域的碳循環產生重要的影響[9, 14]。國內學者對我國北方典型海灣桑溝灣和乳山宮家島以東水域、福建三沙灣鹽田港等涉及貝類養殖的海域碳循環進行了研究[15-17], 分析了養殖櫛孔扇貝對桑溝灣碳循環的貢獻[18]。
本研究根據2018—2019年在廣東考洲洋牡蠣養殖海域4個季節的調查數據, 估算該海域海-氣界面CO2交換通量, 分析其與環境因子的關系, 對研究淺海牡蠣養殖區的碳源匯潛力和海洋碳生物地球化學循環具有重要的科學意義。
1 材料與方法
1.1 調查區域概況
考洲洋是位于粵東沿岸的一個溺谷灣, 通過狹長的鹽洲水道與紅海灣相連, 水域面積約29.7 km2, 灘涂面積約13.73 km2, 是重要的近江牡蠣養殖水域之一, 出產的近江牡蠣稱為“赤岸蠔”, 是地方名優特產; 灣內入海河流主要是吉隆河, 全長22 km, 流域面積116 km2, 年徑流量為2.32×108m3[19-20]。考洲洋口門處的鹽洲島將口門分隔為兩個水道, 建有兩座橋梁與大陸相連, 牡蠣養殖區主要位于鹽洲島以西海域, 沿岸有較多對蝦高位養殖池。
1.2 站位布設與樣品采集
于2018年7月、11月和2019年1月、4月進行夏季、秋季、冬季和春季4個季節的調查。在牡蠣養殖區布設5個站位, 兩座橋梁處各布設1個站位作為非養殖區對照, 北側橋梁為大橋, 南側橋梁為小橋, 調查站位見圖1, 牡蠣養殖區每站采集表層海水(水面下0.5 m)1次, 橋梁處每站采集表層海水3個樣品作為重復。各種樣品的采集、保存、分析和測定均按《海洋監測規范》[21]進行。現場測試水深、水溫()、透明度, 使用Thermo Scientific Orion Star A211型pH計(相對精度±0.01)現場測試pH(NBS標度), 應用碘量法測定溶解氧(DO); 將500 mL水樣經0.45 μm醋酸纖維濾膜過濾后冷凍保存帶回實驗室, 經丙酮萃取后, 用分光光度法測試葉綠素(Chl); 用SYA2-2型實驗室鹽度計測試鹽度(); 水樣現場經WHATMAN囊式濾器過濾后帶回實驗室使用總溶解性無機碳分析儀(Apollo SciTech AS-C3)測試溶解無機碳(DIC)。
1.3 計算方法
海-氣界面CO2交換通量采用以下公式進行估算[22-24]。
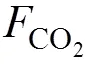
其中, 為海-氣界面CO2交換通量, 單位mmol/(m2·d), 若為負值, 則此區域為大氣CO2的匯, 若為正值, 則此區域為CO2的源; k是海-氣界面氣體傳輸速度(cm/h); as為CO2在海水中的溶解度系數(mol·kg–1·Pa–1), 是與溫度和鹽度相關的函數, 參照Weiss(1974)公式計算得到[25]; Δp(CO2)是海水與大氣中p(CO2)差值。本研究中, 大氣p(CO2)取值為41.61 Pa[15, 26], 海氣界面氣體傳輸速度k取大陸架海區的平均值10.3[16, 27]。
假定海水中CO2體系處于平衡, 如果海水的溫度、鹽度和壓力已經測定, 那么pH、總堿度(TA)、DIC和表層水二氧化碳分壓[(CO2)]就可以任意測定兩個, 然后計算出其余的參數[28]。本文利用pH值和DIC數據, 采用CO2SYS計算CO2體系的其他參數HCO3–、CO32―、CO2和(CO2)[29-31], 碳酸的第一級、第二級解離常數引自Lueker等[32], 總硼酸濃度引自Lee等[33], HF的解離常數引自Perez等[34]。
浮游植物初級生產力可用于估算浮游植物固碳強度[16]。依據海水中Chl質量濃度和透明度計算初級生產力[35]:
P=Chl a×××/2,(2)
式中,P是初級生產力(mg C·m–2·d–1),Chl a是葉綠素的濃度(mg·m–3),是同化系數, 取3.7 mg C/(mg Chl·h)[36],是透光層深度(m), 取透明度的3倍[37],是白天的時長(查詢中國天氣網日出日落時間計算)。
從表6可以看出,2012—2016年青島市總的就業人數不斷增加。2012—2016年青島市第一產業的就業比重呈現逐年遞減趨勢,但趨勢比較平緩;第二產業的就業人員比重同樣呈現逐年遞減趨勢,其中2013—2014年就業人員比重減少最多;第三產業的就業人員比重在逐年遞增,尤其2014年第三產業就業人員的比重增加最大,這是第二產業就業人員大量向第三產業轉移的結果。通過分析三次產業就業結構可以看出,2012年青島市的工業化發展水平屬于工業化后期,2013—2016年屬于工業化中期。
1.4 數據分析
所有數據均用平均值±標準差表示, 應用SPSS Statistics 17.0軟件對不同季節和不同區域的無機碳體系各分量的濃度、(CO2)和CO2海-氣界面交換通量等進行方差分析(ANOVA), 當<0.01時為差異極顯著, 當<0.05時為差異顯著, 數據的多重比較采用SPSS軟件中的LSD法(假定方差齊性)及Dunnett’s T3法(未假定方差齊性)進行分析, 數據相關分析采用Spearman秩相關系數。
2 結果與分析
2.1 海區水環境特征
海區水環境特征值見表1。考洲洋表層海水不同采樣區域的水溫、pH和DO差異不顯著, 鹽度有一定差異。水溫: 夏季>春季>秋季>冬季, 季節變化明顯(<0.05); pH: 春季>冬季>夏季>秋季, 春季和冬季差異不顯著(>0.05); DO: 冬季>春季>夏季>秋季, 冬季和春季、夏季和秋季差異不顯著(>0.05), 鹽度: 春季>冬季>秋季>夏季, 夏季鹽度顯著低于其余3個季節(<0.05)。鹽度在不同區域排序是小橋區>大橋區>牡蠣養殖區, 牡蠣養殖區鹽度顯著低于大橋區和小橋區(0.05)。
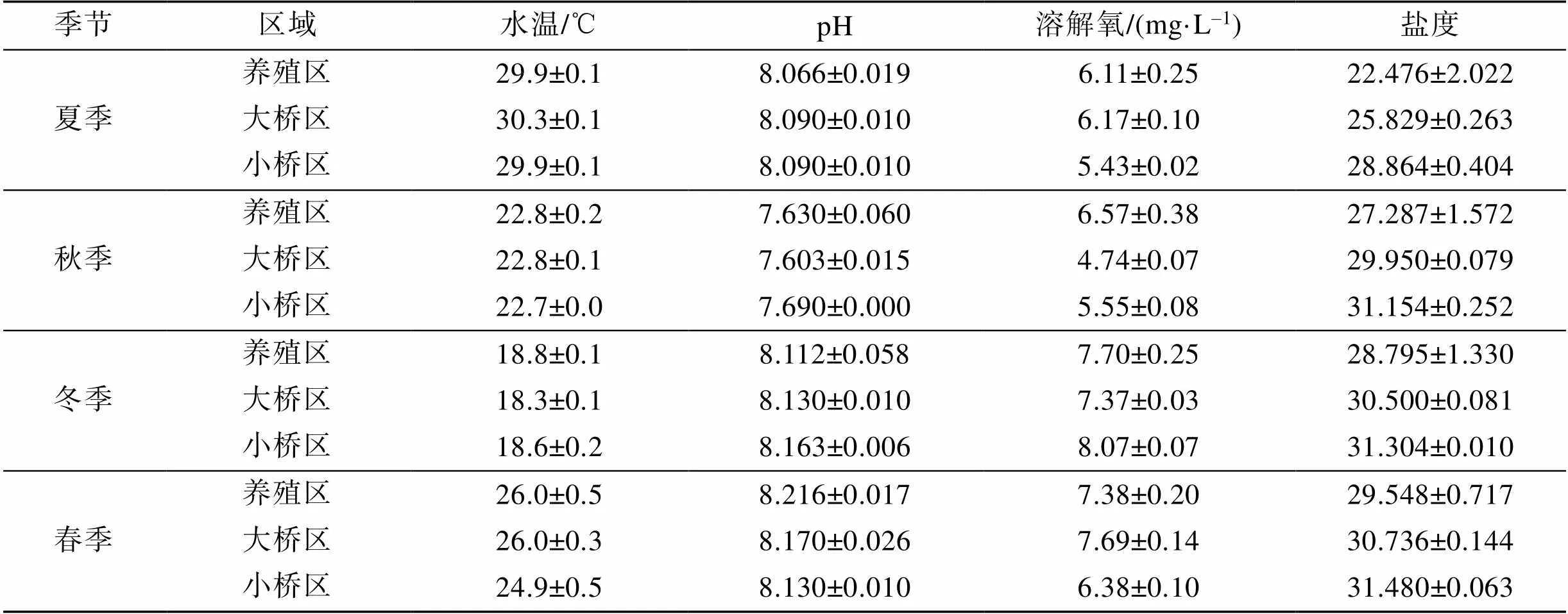
表1 不同季節表層海水基本參數
2.2 表層海水無機碳體系各分量的時空變化
考洲洋表層海水不同季節無機碳體系各分量濃度分布見圖2。牡蠣養殖區DIC、HCO3–、CO32–、CO2質量濃度的年均值分別是1 710.1±159.0、1 577.1±132.4、113.5±54.3、19.5±12.2 μmol/kg;(CO2)的年度變化范圍是(30.70~154.96) Pa, 年均值是(65.18±39.05) Pa。HCO3–是DIC的主要組成成分, 年均占比92.14%±1.98%, CO2占比最低, 年均值1.12%±0.71%。
按年度計算, 養殖區、大橋區和小橋區pH、CO32–、CO2和(CO2)差異均不顯著(>0.05); 牡蠣養殖區DIC、HCO3–質量濃度顯著低于大橋區和小橋區(<0.01), 后兩者的差異不顯著。
在養殖區, 季節對pH及無機碳體系各分量有影響(<0.01), 主效應檢驗各要素的2在0.8及以上, 其中pH、CO32–、CO2和(CO2)的2分別是0.970、0.945、0.957和0.957, 說明在養殖區, pH及無機碳體系各分量差異有80%以上來自于季節變化, 個別要素達90%以上。養殖區按季節排序, pH: 春季>冬季>夏季>秋季, 夏季和冬季的差異不顯著(>0.05); DIC: 冬季>春季>秋季>夏季, 冬季顯著高于其余3個季節(<0.05), 春季顯著高于夏季(<0.05), 秋季與春季、秋季與夏季的差異不顯著(>0.05); HCO3–: 冬季>春季>秋季>夏季, 秋季與其余3個季節的差異不顯著(>0.05), 其余季節之間的差異顯著(<0.05); CO32–: 春季>冬季>夏季>秋季, 春季與冬季顯著高于夏季與秋季(< 0.05), 夏季顯著高于秋季(<0.05); CO2: 秋季>夏季>冬季>春季, 秋季顯著高于其余3個季節(<0.01), 夏季顯著高于春季, 冬季和春季的差異不顯著(>0.05);(CO2): 秋季>夏季>冬季>春季, 春季和冬季的差異不顯著(>0.05), 其余季節的差異極顯著。
2.3 表層海水海-氣界面CO2交換通量的時空變化

圖2 不同季節表層海水pH、p(CO2)、無機碳體系各分量濃度
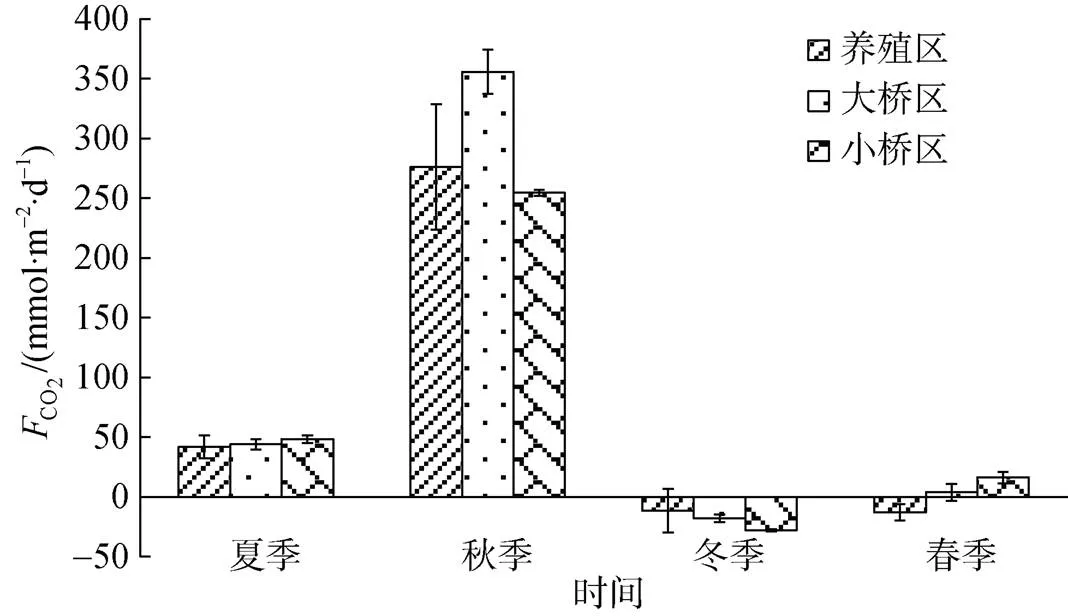
圖3 不同季節海-氣界面CO2交換通量
2.4 表層海水Chl a及初級生產力的時空變化
表層海水Chl及初級生產力(P)分布見圖4。牡蠣養殖區、大橋區和小橋區表層海水Chl質量濃度年度平均值分別是6.83±2.93、11.04±4.35、9.64± 5.65 μg/L, 養殖區的Chl質量濃度顯著低于大橋區(<0.01), 大橋區與小橋區差異不顯著(>0.05)。
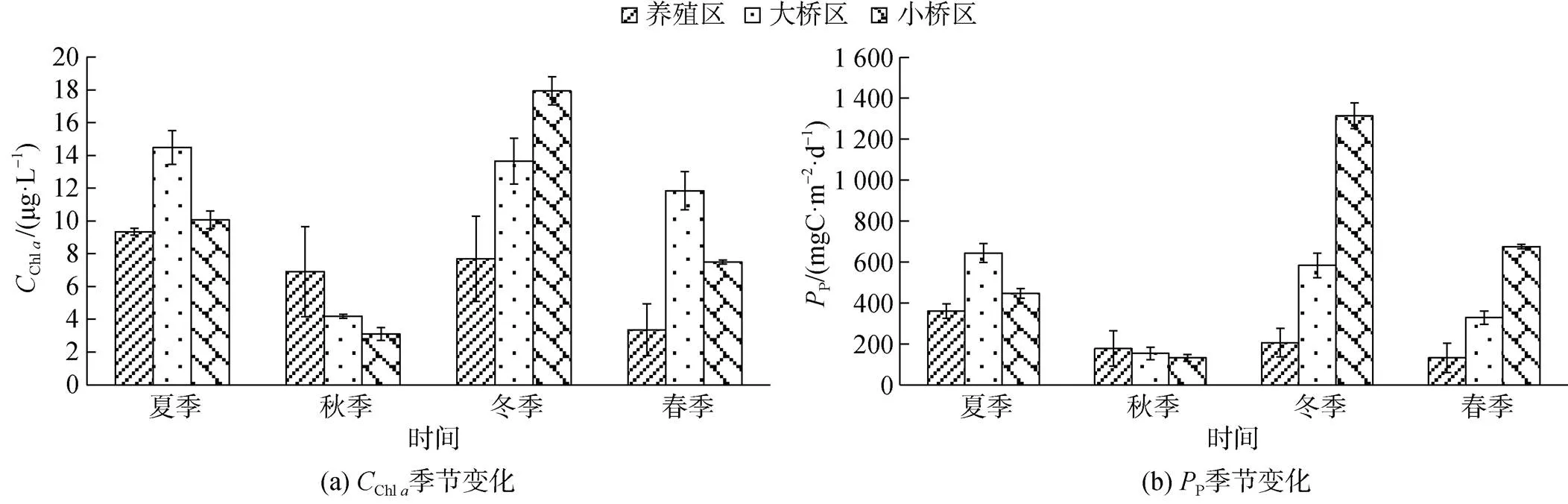
圖4 不同季節表層海水的Chl a濃度及PP
以季節和區域進行主效應檢驗, 季節和區域具有交互作用, Chl及P的2均在0.9以上, 說明季節和區域一起決定了這些要素90%以上的差異。
在夏季、冬季和春季, 牡蠣養殖區的P顯著低于大橋區和小橋區(<0.05); 在秋季, 3個區域的P差異不顯著(>0.05)。
養殖區表層海水Chl及P季節變化是夏季>冬季>秋季>春季。夏季的Chl質量濃度顯著高于春季(<0.01), 其余季節之間的差異均不顯著(>0.05)。夏季P顯著高于其余3個季節(<0.01), 其余3個季節的差異均不顯著(>0.05)。
2.5 海-氣界面CO2交換通量與水環境因子的相關性分析
海-氣界面CO2交換通量與水環境因子的相關系數見表2。從全年尺度來看, CO2交換通量與水溫呈現顯著的正相關, 與pH、DO、鹽度、Chl、DIC和P呈現顯著的負相關。從不同季節來看, CO2交換通量在4個季節與水溫的相關關系均不顯著; 夏季僅與DO呈顯著負相關, 秋季僅與pH呈顯著負相關, 冬季與pH、Chl和P呈現顯著的負相關, 春季與pH呈現顯著負相關, 與鹽度、Chl、DIC和P呈現顯著的正相關。
3 討論
3.1 環境因子對表層海水p(CO2)及CO2交換通量的影響
考洲洋是一個溺谷灣, 水交換較弱, 灣區海洋碳循環受灣外影響較弱, 夏季和秋季是養殖旺季, 冬季和春季大部分牡蠣已被收獲, 是養殖淡季。本研究中, 考洲洋表層海水的DIC濃度低于我國北方典型海灣桑溝灣和乳山宮家島以東水域[15-16], 高于福建三沙灣鹽田港[17], 這可能與考洲洋所在的地理區位和水文條件有關。
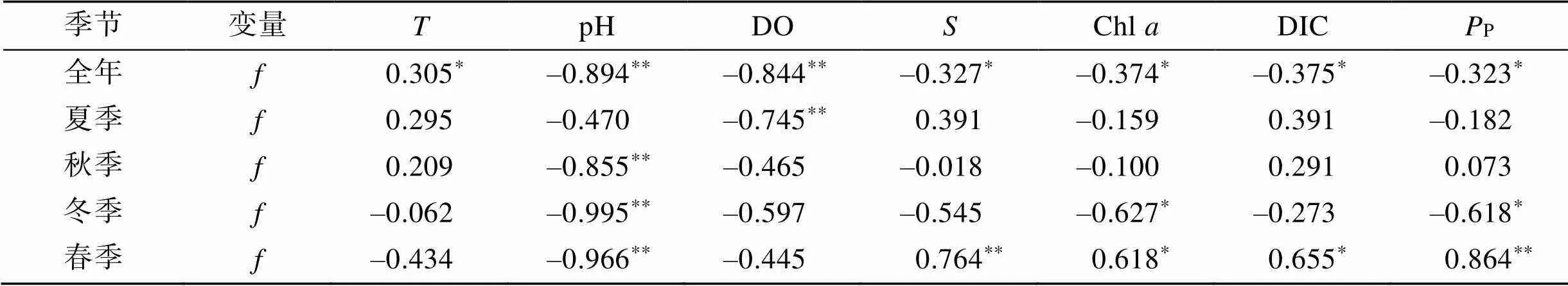
表2 CO2交換通量與水環境因子的Spearman相關系數
注: *表示相關性顯著(0.05), **表示相關性極顯著(0.01)。
表層海水(CO2)會受到物理、化學、生物的各種影響, 水溫和鹽度是重要的影響因素之一[25]。沿岸海水溫度主要受到太陽輻射和海洋大氣熱交換的影響, 海水鹽度主要受到淡水(包括降雨、徑流和地下水等)、外海高鹽水和蒸發作用的影響[28]。考洲洋水深較淺, 無大的徑流輸入, 受太陽輻射影響較大, 同一季節不同區域未形成顯著的水溫差, 太陽輻射的季節變化決定了水溫的主要差異; 夏季降雨較多, 沖淡水降低海水鹽度, 夏季的鹽度顯著低于其余3個季節; 養殖區位于灣頂, 受外海高鹽水的影響相對更小, 養殖區的鹽度顯著低于非養殖區。一般而言, 水溫對海水中(CO2)具有雙重影響, 海水中的碳酸鹽和其他弱酸鹽體系的平衡隨水溫的變化而變化, 海水中(CO2)隨水溫的升高而升高, 水溫升高又導致CO2溶解度降低, 引起海水中(CO2)降低。海水鹽度增加, 離子強度增大, CO2的溶解度降低, 海水(CO2)減小[38]。在全年尺度, 考洲洋表層海水(CO2)及CO2交換通量與水溫呈顯著正相關關系, 與鹽度呈顯著負相關關系, 但在單個季節均未呈現此規律。4個季節調查中, 表層海水(CO2)及CO2交換通量與水溫均無顯著的相關關系; 夏季、秋季和冬季表層海水(CO2)及CO2交換通量與鹽度均無顯著的相關關系, 春季反而呈顯著的正相關關系。可能的原因是水溫和鹽度的物理影響屬于客觀存在, 在(CO2)及CO2交換通量季節變化上起主導作用; 而在單個季節, 養殖牡蠣、浮游生物的生理生態過程與海域水溫、鹽度等物理過程耦合作用, 導致表層海水(CO2)及CO2交換通量的變化呈現出不同的變化。
浮游植物光合作用產生的固碳效應會降低海水中的(CO2)[39]。(CO2)與Chl成顯著負相關時, 可以推斷光合作用對影響表層水(CO2)起到決定性作用[15-16]。考洲洋4個季節調查中, 僅冬季表層海水(CO2)及CO2交換通量與Chl、初級生產力呈顯著負相關關系, 說明冬季浮游植物光合作用可能對影響表層水(CO2)起主導作用。在非養殖區, 表層海水(CO2)及CO2交換通量與Chl、初級生產力呈顯著負相關關系, 在養殖區的相關關系不顯著, 說明在非養殖區, 浮游植物光合作用可能對影響表層水(CO2)起主導作用。
3.2 養殖活動對表層海水p(CO2)及CO2交換通量的影響
養殖生態系統的復雜性使得(CO2)及CO2交換通量存在較大的不均勻性[40]。在淺海貝類養殖生態系統中, 浮游植物和養殖貝類是影響(CO2)的重要因素[16]。浮游植物的光合作用消耗CO2, 產生O2, 降低水體中的(CO2)[39]; 養殖貝類在形成貝殼的鈣化過程利用水體DIC的同時, 與呼吸過程協同釋放CO2, 增加水體中的(CO2)[41], 使得貝類在表現為生物碳匯的同時還表現為部分碳源[42]。貝類鈣化作用反應式如下: Ca2++2 HCO3–=CaCO3+CO2+H2O[14], 雖然每形成1 mol的碳酸鈣, 會釋放1 mol的CO2, 但可以吸收2 mol的HCO3–, 實際固定1mol碳[12, 41]。
牡蠣養殖活動通過鈣化作用和呼吸作用可直接影響養殖區的海洋碳循環。本研究中, 在養殖區, 養殖牡蠣鈣化作用大量利用海水中的HCO3–, 形成CaCO3軀殼, 導致水體中HCO3–降低, 表現為養殖區的HCO3–顯著低于非養殖區; 其中養殖旺季的鈣化作用強度大于養殖淡季, 表現為養殖區養殖旺季的HCO3–顯著低于養殖淡季。在非養殖區沒有養殖牡蠣消耗HCO3–, 養殖淡季和養殖旺季的HCO3–無顯著差異。不管是養殖旺季還是養殖淡季, 考洲洋海域養殖區和非養殖區表層海水的(CO2)及CO2交換通量無顯著差異, 與蔣增杰等[16]的研究一致, 養殖牡蠣鈣化、呼吸作用等生理因素釋放的CO2對表層海水(CO2)及CO2交換通量未產生顯著影響。
另一方面, 牡蠣養殖活動也可通過濾食作用降低浮游植物密度(攝食壓力)和通過代謝作用增加海水中的營養物質促進浮游植物增殖(促增殖作用), 影響浮游植物的光合作用間接影響海洋碳循環。養殖區和非養殖區Chl含量差異不顯著表明養殖牡蠣的攝食壓力尚未對浮游植物群落造成顯著影響[16]。本研究中, 在全年尺度, 養殖牡蠣攝食壓力總體可能大于其促增殖作用, 表現為養殖區的Chl和P顯著低于非養殖區。在養殖旺季, 養殖牡蠣攝食壓力與其促增殖作用可能相互抵消, 表現為養殖區和非養殖區之間的Chl和P差異不顯著。在養殖淡季, 養殖活動補充的營養物質減少, 而吉隆河和鹽洲水道給非養殖區補充的營養物質源源不斷, 營養物質在非養殖區的促增殖作用可能大于在養殖區, 表現為非養殖區的Chl和P顯著高于養殖區。在養殖區, 牡蠣養殖活動在養殖旺季的促增殖作用可能大于在養殖淡季, 表現為養殖旺季的Chl和P顯著高于養殖淡季, 但在非養殖區, 養殖旺季的Chl和P卻顯著低于養殖淡季。說明牡蠣的養殖活動已經對養殖區浮游植物群落造成復雜的影響, 但在不同的養殖階段其作用如何實現還需結合營養鹽的變化進一步研究。
考洲洋養殖海域養殖旺季為CO2的源, 秋季的源強最大; 養殖淡季整體為CO2的弱匯, 而北方桑溝灣貝類養殖區春季和乳山宮家島以東牡蠣養殖水域秋季均表現為CO2的匯區[15-16]。在夏季和秋季, 考洲洋牡蠣養殖區CO2源強和非養殖區的差異不顯著; 在冬季, 養殖區CO2源強整體上和非養殖區的差異不顯著, 但非養殖區為CO2的匯, 養殖區有的站位為匯, 有的站位為源, 這與浮游植物的光合作用影響有關, 表現為匯的地方Chl相對較高, 且附近的牡蠣均被收獲, 源的地方Chl相對較低, 且附近有少量牡蠣未被收獲; 在春季, 牡蠣養殖區為CO2的匯, 非養殖區總體上為源。
養殖牡蠣鈣化作用形成的CaCO3軀殼, 絕大部分通過收獲從海水中移除, 是一種碳的地球化學循環過程(循環周期長達數百萬年)[14], 以貝殼掩埋或功能化利用實施碳封存, 可成為持久的碳匯, 對移除、降低大氣中的CO2濃度, 緩解溫室效應具有一定的應用前景。此外, 貝類濾食水體中的顆粒有機碳, 一部分促進貝類軟體組織生長, 另一部分以糞和假糞的形式形成生物學沉積物, 具有更大的粒徑的更高的有機物, 沉降速率大于自然的懸浮顆粒物, 可加速碳從水環境移出[43]。
4 結論
1) 牡蠣養殖區表層海水中Chl、DIC、HCO3–和初級生產力顯著低于非養殖區, 養殖區和非養殖區的pH、DO、CO32–、(CO2)及CO2交換通量差異不顯著。養殖淡季表層海水中pH、DO、DIC、HCO3–和CO32–顯著大于養殖旺季, 養殖旺季的(CO2)和CO2交換通量顯著大于養殖淡季, 養殖淡季和養殖旺季的Chl和初級生產力差異不顯著。養殖區DIC、HCO3–呈現冬季>春季>秋季>夏季的季節變化特征, CO2、(CO2)、CO2交換通量呈現秋季>夏季>冬季>春季的季節變化特征。
2) 考洲洋海域水溫和鹽度的季節變化可能對表層海水(CO2)及CO2交換通量季節變化起主導作用。在冬季或非養殖區, 浮游植物光合作用可能對影響表層海水(CO2)及CO2交換通量起主導作用。養殖牡蠣的鈣化、呼吸作用等生理因素釋放的CO2對表層海水(CO2)及CO2交換通量并未產生顯著影響。
3) 考洲洋養殖海域養殖旺季為CO2的源, 秋季的源強最大, 養殖淡季整體為CO2的弱匯。養殖牡蠣呼吸作用和鈣化作用釋放CO2的過程使其在表現為生物碳匯的同時還表現為部分碳源, 鈣化作用形成的CaCO3軀殼是持久的碳匯, 對移除、降低大氣中的CO2濃度有積極的意義, 是一種有效的碳中和措施, 對于發展低碳經濟, 實現“碳中和”目標具有重要意義。
[1] SOLOMON S, PLATTNER G K, KNUTTI R, et al. Irreversible climate change due to carbon dioxide emissions[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(6): 1704-1709.
[2] 唐啟升, 陳鎮東, 余克服, 等. 海洋酸化及其與海洋生物及生態系統的關系[J]. 科學通報, 2013, 58(14): 1307-1314.
TANG Qisheng, CHEN Zhendong, YU Kefu, et al. The effects of ocean acidification on marine organisms and ecosystem[J]. Chinese Science Bulletin, 2013, 58(14): 1307-1314.
[3] PACHAURI R K, ALLEN M R, BARROS V R, et al. Climate change 2014: Synthesis report[R]. Contri-bu-tion of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change, IPCC, Geneva, 2015.
[4] SABINE C L, FEELY R A, GRUBER N, et al. The oceanic sink for anthropogenic CO2[J]. Science, 2004, 305(5682): 367-371.
[5] 焦念志. 研發海洋“負排放”技術支撐國家“碳中和”需求[J]. 中國科學院院刊, 2021, 36(2): 179-187.
JIAO Nianzhi. Developing ocean negative carbon emis-sion technology to support national carbon neutralization[J]. Bulletin of Chinese Academy of Sciences, 2021, 36(2): 179-187.
[6] NELLEMANN C, CORCORAN E, DUARTE C M, et al. A Rapid Response Assessment. Blue carbon: The role of healthy oceans in binding carbon[M]//United Nations Environment Programme, GRID-Arendal. Birkeland Trykkeri AS, Norway, 2009.
[7] GRUBER N, CLEMENT D, CARTER B R, et al. The oceanic sink for anthropogenic CO2from 1994 to 2007[J]. Science, 2019, 363(6432): 1193-1199.
[8] DONEY S C, FABRY V J, FEELY R A, et al. Ocean acidification: The other CO2problem[J]. Annual Review of Marine Science, 2009, 1(1): 169-192.
[9] 張繼紅, 方建光, 唐啟升. 中國淺海貝藻養殖對海洋碳循環的貢獻[J]. 地球科學進展, 2005, 20(3): 359- 365.
ZHANG Jihong, FANG Jianguang, TANG Qisheng. The contribution of shellfish and seaweed mariculture in China to the carbon cycle of coastal ecosystem[J]. Advances in Earth Science, 2005, 20(3): 359-365.
[10] 唐啟升. 碳匯漁業與又好又快發展現代漁業[J]. 江西水產科技, 2011(2): 9-11.
TANG Qisheng. Carbon sink fishery and good and fast development of modern fishery[J]. Jiangxi Fishery Science and Technology, 2011(2): 9-11.
[11] 王清印. 碳匯漁業——概念、發展與展望[C]//海洋資源科學利用論壇論文集. 煙臺: 2011, p82-85.
WANG Qingyin. The concept, development and pro-spects of carbon sink fisheries[C]// Collection of Essays of Forum on Scientific Utilization of Marine Resource. Yantai, 2011: 82-85.
[12] 唐啟升, 劉慧. 海洋漁業碳匯及其擴增戰略[J]. 中國工程科學, 2016, 18(3): 68-73.
TANG Qisheng, LIU Hui. Strategy for carbon sink and its amplification in marine fisheries[J]. Strategic Study of CAE, 2016, 18(3): 68-73.
[13] TANG Q S, ZHANG J H, FANG J G. Shellfish and seaweed mariculture increase atmospheric CO2absorption by coastal ecosystems[J]. Marine Eco-logy Progress Series, 2011, 424: 97-104.
[14] 李海晏, 陳濤, 張海燕, 等. 中國貝類養殖對海洋碳循環的貢獻評估[J]. 海洋科學, 2014, 38(5): 39-45.
LI Haiyan, CHEN Tao, ZHANG Haiyan, et al. Evalua-tion of contribution of shellfish culture to ocean carbon cycle in China[J]. Marine Sciences, 2014, 38(5): 39-45.
[15] 劉毅, 張繼紅, 房景輝, 等. 桑溝灣春季海-氣界面CO2交換通量及其與養殖活動的關系分析[J]. 漁業科學進展, 2017, 38(6): 1-8.
LIU Yi, ZHANG Jihong, FANG Jinghu, et al. Analysis of the air-sea surface carbon dioxide flux and its intera-ction with aquaculture activities in Sanggou Bay[J]. Progress in Fishery Sciences, 2017, 38(6): 1-8.
[16] 蔣增杰, 方建光, 王巍, 等. 乳山宮家島以東牡蠣養殖水域秋季海-氣界面CO2交換通量研究[J]. 水產學報, 2012, 36(10): 1592-1598.
JIANG Zengjie, FANG Jianguang, WANG Wei, et al. Sea-air CO2flux in Crassostrea gigas aquaculture area of East Gongjia Island, Rushan, Shandong, in autu-mn[J]. Journal of Fisheries of China, 2012, 36(10): 1592-1598.
[17] 韋章良, 韓紅賓, 胡明, 等. 三沙灣鹽田港海水養殖海域海-氣界面CO2交換通量的時空變化[J]. 上海海洋大學學報, 2016, 25(1): 106-115.
WEI Zhangliang, HAN Hongbin, HU Ming, et al. Sea-sonal variation of sea-air CO2flux in mariculture area in Yantian Harbor, Sansha Bay[J]. Journal of Shanghai Ocean University, 2016, 25(1): 106-115.
[18] 張明亮, 鄒健, 毛玉澤, 等. 養殖櫛孔扇貝對桑溝灣碳循環的貢獻[J]. 漁業現代化, 2011, 38(4): 13-16.
ZHANG Mingliang, ZOU Jian, MAO Yuze, et al. Contribution of culturing scallop to carbon cycle in Sanggou Bay[J]. Fishery Modernization, 2011, 38(4): 13-16.
[19] 李純厚, 林欽, 蔡文貴, 等. 考洲洋浮游植物種類組成與數量分布特征[J]. 水產學報, 2006, 29(3): 379- 385.
LI Chunhou, LIN Qin, CAI Wengui, et al. Charac-te-ristics of species composition and quantity distribution of phytoplankton in Kaozhouyang Bay[J]. Journal of Fisheries of China, 2006, 29(3): 379-385.
[20] 田志群, 董永宏. 赤岸蠔(近江牡蠣)標準化育肥養成技術[J]. 海洋與漁業, 2016, 7: 56-58.
TIAN Zhiqun, DONG Yonghong. Chian oyster (Gould) standardized fattening cultivation technology[J]. Ocean and Fishery, 2016, 7: 56-58.
[21] 中華人民共和國國家質量監督檢驗檢疫總局. GB 17378.4-2007 海洋監測規范第4部分: 海水分析[S]. 北京: 中國標準出版社, 2008.
General Administration of Quality Supervision, Inspe-ction and Quarantine of the People’s Republic of China. GB 17378.4-2007 The specification for marine monitoring-Part 4: Seawater analysis[S]. Beijing: Standards Press of China, 2008.
[22] WANNINKHOF R. Relationship between wind speed and gas exchange over the ocean[J]. Journal of Geo-physica1 Research, 1999, 97(C5): 7373-7382.
[23] REHDER G, SUESS E. Methane andCO2in the Kuroshio and the South China Sea during maximum summer surface temperatures[J]. Marine Chemistry, 2001, 75(1/2): 89-108.
[24] TERNON J F, OUDOT C, DESSIER A, et al. A sea-sonal tropical sink for atmospheric CO2in the Atlantic Ocean: The role of the Amazon River discharge[J]. Marine Chemistry, 2000, 68(3): 183- 201.
[25] WEISS R F. Carbon dioxide in water and seawater: the solubility of a non-ideal gas[J]. Marine Chemistry, 1974, 2(3): 203-215.
[26] World Meteorological Organization.WMO greenhouse gas bulletin (GHG bulletin): The state of greenhouse gases in the atmosphere based on global observations through 2020[R]. World Meteorological Organization (WMO), 2021, 17: 1-10.
[27] SHIZUO, TSUNOGAI, SHUICHI, et al. Is there a “continental shelf pump” for the absorption of atmos-pheric CO2?[J]. Tellus B, 1999, 51(3): 701-712.
[28] 陳敏. 化學海洋學[M]. 北京: 海洋出版社, 2009.
CHEN Min. Chemical oceanography[M]. Beijing: Ocean Press, 2009.
[29] LEWIS E, WALLACE D. Program developed for CO2system calculations[CP]. Carbon Dioxide Information Analysis Center, managed by Lockheed Martin Energy Research Corporation for the US Department of Energy Tennessee. 1998. https://www.researchgate.net/publication/ 240194526_CO2SYS-Program_developed_for_CO2_system_calculations.
[30] HEUVEN S V, PIERROT D, Rae J W B, et al. CO2SYS v 1.1, MATLAB program developed for CO2system calculations[CP]. 2011. https://www.researchgate.net/ publication/282354342_CO2SYS_v_11_MATLAB_program_developed_for_CO2_system_calculations.
[31] XU Y Y, PIERROT D, CAI W J. Ocean car-bonate system computation for anoxic waters using an updated CO2SYS program[J]. Marine Chemistry, 2017, 195(20): 90-93.
[32] LUEKER T J, DICKSON A G, KEELING C D. OceanCO2calculated from dissolved inorganic carbon, alkalinity, and equations for K1and K2: Validation based on laboratory measurements of CO2in gas and seawater at equilibrium[J]. Marine Chemistry, 2000, 70(1): 105-119.
[33] LEE K, KIM T W, BYRNE R H, et al. The universal ratio of boron to chlorinity for the North Pacific and North Atlantic oceans[J]. Geochimica Et Cosmo-chi-mica Acta, 2010, 74(6): 1801-1811.
[34] PEREZ F F, FRAGA F. Association constant of fluoride and hydrogen ions in seawater[J]. Marine Chemistry, 1987, 21(2): 161-168.
[35] CADEE G C, HEGEMAN J. Primary production in the Dutch Wadden Sea[J]. Netherlands Journal of Sea Research, 1974, 8: 240-259.
[36] RYTHER J H, YENTSCH C S. The estimation of phy-toplankton production in the ocean from chlorophyll and light data[J]. Limnology and Oceanography, 1957, 2(3): 281-286.
[37] YUKUYA Y, SHUN-EI I. Dynamic status of primary production in Lake Yunoko, a small eutrophic subalpine lake in central Japan[J]. The Botanical Magazine Tokyo, 1972, 85(3): 157-176.
[38] 朱連磊, 宋金明, 李學剛, 等. 東海中北部海域秋季表層海水中無機碳與海氣界面碳的遷移[J]. 海洋科學, 2012, 36(10): 26-32.
ZHU Lianlei, SONG Jinming, LI Xuegang, et al. Dissolved inorganic carbon and I air-sea carbon exchange flux in the central and northern East China Sea in autumn[J]. Marine Sciences, 2012, 36(10): 26-32.
[39] 張龍軍, 王婧婧, 張云, 等. 冬季北黃海表層海水CO2分布及其影響因素探討[J]. 中國海洋大學學報(自然科學版), 2008, 38(6): 955-9601.
ZHANG Longjun, WANG Jingjing, ZHANG Yun, et al. Distribution and controlling factors of surface seawater partial pressure of CO2in the Northern Yellow Sea during winter[J]. Periodical of Ocean University of China, 2008, 38(6): 955-9601.
[40] 張繼紅, 吳文廣, 任黎華, 等. 桑溝灣表層水CO2的季節變化及影響因素分析[J]. 漁業科學進展, 2013, 34(1): 57-64.
ZHANG Jihong, WU Wenguang, REN Lihua, et al. Seasonal variation ofCO2and its potential influe-n-cing factors in aquaculture areas of Sanggou Bay[J]. Progress in Fishery Sciences, 2013, 34(1): 57-64.
[41] CHAUVAUD L, THOMPSON J K, CLOERN J E, et al. Clams as CO2generators: Theexample in San Francisco Bay[J]. Limnology and Oceanography, 2003, 48(6): 2086-2092.
[42] CLAIRE G, FRANCK G, DOMINIQUE D. Secondary production, calcification and CO2fluxes in the cirri-pedesand[J]. Oecologia, 2008, 155(1): 133-142.
[43] 齊占會, 王珺, 黃洪輝, 等. 廣東省海水養殖貝藻類碳匯潛力評估[J]. 南方水產科學, 2012, 8(1): 30-35.
QI Zhanhui, WANG Jun, HUANG Honghui, et al. Potential assessment of carbon sink capacity by marine bivalves and seaweeds in Guangdong Province[J]. South China Fisheries Science, 2012, 8(1): 30-35.
Temporal and spatial variation in the sea–air CO2flux in the oyster aquaculture area of Kaozhou Bay
PENG Peng-fei1, MA Yuan1, SHI Rong-jun2, WANG Di1, XU Xin1, YAN Bin1
(1. South China Sea Marine Survey and Technology Center, State Oceanic Administration, Key Laboratory of Marine Environmental Survey Technology and Application, Ministry of Natural Resources, Guangzhou 510300, China; 2. South China Sea Fisheries Research Institute, Chinese Academy of Fisheries Sciences, Guangdong Provincial Key Laboratory of Fishery Ecology and Environment, Guangzhou 510300, China)
dissolved inorganic carbon;CO2; CO2flux; oyster aquaculture; Kaozhou Bay
Oct. 31, 2021
Q176; X55
A
1000-3096(2022)10-0140-10
10.11759/hykx20211031001
2021-10-31;
2022-01-14
廣東省促進經濟發展專項資金(GDME-2018E006); 廣東省漁業生態環境重點實驗室開放基金項目(FEEL-2017-6)
[Special Fund for Promoting Economic Development of Guang-dong Province, No. GDME-2018E006; Open Fund of Guangdong Provincial Key Laboratory of Fishery Ecology and Environment, No. FEEL-2017-6]
彭鵬飛(1985—), 男, 湖北天門人, 碩士, 主要從事海洋生態和環境科學研究, E-mail: pepefei@163.com
(本文編輯: 趙衛紅)
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
