培養基中添加植物生長調節劑的蛹蟲草轉錄組學分析
2022-11-28 03:26:16王哲王升厚李夢雪柳葉飛王澤徐方旭
食品工業 2022年11期
王哲,王升厚,李夢雪,柳葉飛,王澤,徐方旭*
1.沈陽師范大學生命科學學院(沈陽 110034);2.沈陽師范大學實驗教學中心(沈陽 110034)
蛹蟲草(Cordyceps militaris)又稱北冬蟲夏草、北蟲草,2009年被批準為新資源食品[1-4]。蛹蟲草富含多種生物活性成分,如蟲草素、噴司他丁、腺苷、蟲草酸、蟲草多糖、超氧化物歧化酶、麥角甾醇及黃酮類物質等[5-9],具有抗腫瘤、免疫調節、抗炎抗菌及抗氧化等藥理作用[10-12]。蛹蟲草通過人工培育實現規模化生產,但在產業發展中仍存在亟待解決的技術性瓶頸問題,其中之一就是蛹蟲草在生長后期常出現“白毛病”,即學術界統稱的因感染“吃草菌”引起的真菌性病害,這種寄生性病原真菌會導致蛹蟲草產量嚴重下降[13-16]。
植物生長調節劑是一類與植物激素具有相似生理和生物學效應的物質,其主要成分有水楊酸、茉莉酸、多效唑和多胺。由于植物生長調節劑大多取自植物本身產生的天然產物,具有微量、高效、低毒等作用,同時還可調節作物的生長發育、提高生物轉化率、改善品質外,在作物抗病中具有顯著功效[17-20]。有研究表明,細胞壁是植物防御病原物侵染的第一道屏障。當病原物物侵染植物后,引起細胞壁的防御反應,參與細胞生命活動的某些信息傳遞,并發生防御反應,細胞壁在植物抗病過程中起著重要的能動作用[21-22]。除了細胞壁能夠抵御病原菌的侵染外,近年來,更多研究表明,一些抗病蛋白需要轉移到細胞核內才能啟動免疫反應,進而發揮抗病防御作用,而細胞核質轉運受體是實現這些抗病蛋白核質轉運必不可少的“載體”。因此,細胞核質轉運及轉運受體在抗病防御中發揮重要作用[23]。何茂華[24]研究表明,植物生長調節劑在誘導棉花抗黃萎病期間,其棉苗體內與抗病相關酶活性和一些次生代謝物質含量的變化與棉苗抗黃萎病性相關。肖翔等[25]研究表明,脫落酸通過拮抗水楊酸抗病信號通路負調控擬南芥對白粉菌的穿透后抗性。因此,了解植物對病原菌的防御機制及病原菌對植物的感染過程和分子機理尤為重要,有利于掌握設計植物的持久抗性所需的必要知識,為改善作物對不良環境的抵御能力提供方案和對策[26]。
關于植物生長調節劑誘導蛹蟲草抵抗寄生性病原真菌分子機制的研究鮮見報道。為揭示培養基中添加GN-01的蛹蟲草與對照組蛹蟲草鮮品的基因表達差異,試驗利用RNA-seq技術進行高通量測序和生物信息學分析,對GN-01抵抗蛹蟲草寄生性病原真菌的抗病機理在轉錄組學層面進行探究,為后期開展蛹蟲草真菌病害的防控提供理論依據。
1 材料與方法
1.1 材料與試劑
對照組(CK):栽培培養基為小麥40 g和清水60 g培養的蛹蟲草。
試驗組(DK):栽培培養基中加入稀釋3 000倍GN-01培養的蛹蟲草。
蛹蟲草PT07(沈陽師范大學特種菌業研究所);總RNA提取試劑RNAiso Plus[寶生物工程(大連)有限公司];脫氧核糖核酸酶I(北京凡知醫學科技有限公司);NEBNext Ultra RNA文庫制備試劑盒(美國NEB公司);Oligo(d T)磁珠(蘇州海貍公司);fragmentation buffer(上海恒斐生物科技有限公司);NEBNext Ultra RNA文庫制備試劑盒(美國NEB公司)。
1.2 儀器與設備
Illumina HiSeq 4000高通量測序平臺系統(上海派諾森生物科技有限公司);Agilent 2100生物分析儀(安捷倫科技有限公司)。
1.3 方法
1.3.1 總RNA提取和轉錄組測序
稱取對照組和試驗組蛹蟲草樣本,各200 mg,每組樣品選取3個生物學重復,轉錄組測序委托上海派諾森生物科技有限公司完成[27]。
1.3.2 差異基因篩選
將表達差異倍數|log2Fold Change|>1和顯著性P值<0.05作為篩選差異表達基因條件,滿足此篩選標準的基因即為差異表達基因。
1.3.3 差異基因的功能注釋和富集
應用基因本體論聯合會建立的數據庫所建立的數據庫GO對所獲得差異基因的GO進行分類注釋,同時利用京都基因與基因組百科全書(KEGG)數據庫對差異基因進行KEGG通路的功能富集分析[17]。
2 結果與分析
2.1 質控數據分析
CK組和DK組蛹蟲草轉錄組測序原始數據讀序分別為45 086 267和44 293 419,經過對數據去除雜質及接頭序列后,相應的干凈讀序為42 704 425和41 868 819,其中各組中質量值≥20的堿基所占比例Q20均在97.39%以上,質量值≥30的堿基所占比例Q30均在93.58%以上,表明測序結果理想,可用于后續分析。
2.2 差異基因表達分析
對照組和試驗組共有9 618個差異基因,根據P值<0.05和|log2Fold Change|>1篩選顯著差異表達基因。從圖1可以看出,培養基中添加GN-01后,蛹蟲草子實體顯著差異基因共189個,其中上調基因83個、下調基因106個。
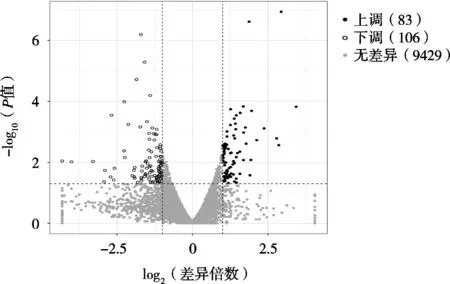
圖1 差異表達基因的火山圖
2.3 差異基因的GO富集分析
GO功能分類結果(圖2)表明:差異基因在細胞組分這類中,核小體、DNA復合包裝、蛋白質-DNA復合體3個條目上富集較顯著;在分子功能這類中,蛋白質異源二聚體活動、糖基鍵的水解酶活性2個條目上富集較顯著;在生物過程這類中,碳水化合物分解代謝過程和跨膜轉運2個條目富集較顯著。

圖2 GO功能注釋
2.4 差異基因的KEGG富集分析
從圖3可以看出:189個差異基因共富集到29條通路,其中,顯著富集到糖鞘脂生物合成-globo和isglobo系列、淀粉和蔗糖代謝、酮體的合成與降解、糖胺聚糖降解、生物素代謝這5條通路,說明蛹蟲草子實體中的差異表達基因在這5條通路上受GN-01的影響較大。
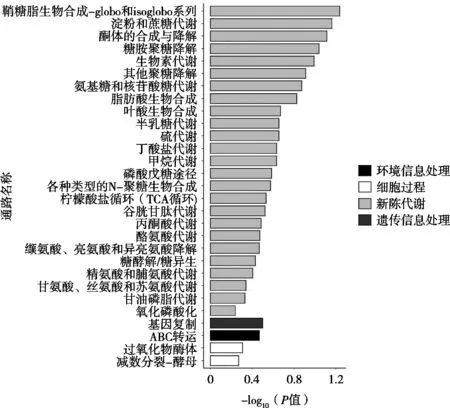
圖3 KEGG功能富集圖
2.5 關鍵KEGG通路與差異表達基因解析
栽培培養基中添加GN-01后蛹蟲草子實體中關鍵KEGG通路與差異表達基因詳見表1。其中:與次生代謝產物生物合成有關的基因為MGAM,FabG和GALM;P4HA主要參與精氨酸和脯氨酸的代謝;GSTK1主要參與植物過氧化物酶體路徑。由此可見,在蛹蟲草栽培培養基中添加GN-01,可有效激活次生代謝產物合成通路、脯氨酸代謝途徑和過氧化物酶體途徑,從而提高蛹蟲草在生長過程中的抗病能力。

表1 關鍵KEGG通路與差異表達基因
3 討論與結論
植物生長調節劑是人工合成或提取的具有天然植物激素生理活性的化合物,可用于調節或控制植物生長發育的某些過程。研究表明,植物生長調節劑通常可在植物體內傳導至作用部位,以很低的濃度促進植物生長和繁殖,其特點為效用高、用量小、毒性低,在全球農業生產中被廣泛應用,達到穩產增產、改善品質、縮短生長周期、延長保鮮期等作用[28-29]。試驗結果表明,在栽培培養基中添加GN-01后,蛹蟲草子實體中顯著差異基因189個,共富集到代謝通路29條。其中,差異基因MGAM,FabG和GALM主要參與次生代謝產物的生物合成,試驗發現,次生代謝產物廣泛參與植物生長、發育、防御等生理過程,可作為生化壁壘抵抗病原物的侵染過程,還可作為信號物質參與植物的抗病反應,在植物的抗病防御反應中發揮重要的作用[30]。差異基因P4HA主要參與精氨酸和脯氨酸的代謝,高水平的脯氨酸可通過維持細胞膜的完整性和增強抗氧化能力以提高植物的抗病能力。GSTK1主要參與植物過氧化物酶體路徑,過氧化物酶體的功能是可以使毒性物質失活,這種作用是過氧化氫酶利用過氧化氫氧化各種底物,如酚、甲酸、甲醛和乙醇等,氧化的結果使這些有毒性的物質變成無毒性的物質,從而提高植物的抗病性。由此可見,在蛹蟲草栽培培養基中添加GN-01,可有效激活次生代謝產物合成通路、脯氨酸代謝途徑和過氧化物酶體途徑,從而提高蛹蟲草在生長過程中的抗病能力。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
