南疆棗園土壤產鐵載體細菌的群落結構
2022-12-05 06:53:52王利娜劉偉鋒魏喜喜劉豐鳴李建貴張國林
經濟林研究 2022年3期
趙 文,王利娜,劉偉鋒,魏喜喜,劉豐鳴,黃 瑤,孫 佳,萬 勝,李建貴,張國林
(1.新疆農業大學 a.草業與環境科學學院;b.林學與風景園林學院;c.實習林場,新疆 烏魯木齊 830052;2.新疆紅棗工程技術研究中心,新疆 烏魯木齊 830013;3.新疆大學 資源與環境科學學院,新疆 烏魯木齊 830046)
鐵是植物生長發育所必需的微量元素之一[1]。然而,在自然環境中,鐵主要以不溶性Fe3+(10-18mol/L,pH 7.0)的形式存在[2]。土壤本底中鐵的含量雖然較高,但在通氣性特別好的土壤中,特別是在堿性土中,可供植物吸收并利用的鐵非常少。世界上約1/3 的土壤為堿性,在南疆紅棗主要種植區內均為堿性土壤,植物對土壤中鐵的利用效率較低。因此,鐵是紅棗種植中常見的限制性因素之一,也是影響新疆紅棗產業發展的因素之一。對土壤中鐵的生物性能的改善已成為南疆棗園土壤養分和棗樹營養改善的研究重點。隨土壤鹽分含量增加,土壤中有效鐵含量進一步降低,僅憑自身作用吸收的鐵元素遠不能滿足植物的需要[3]。由于氧化作用和堿性土壤環境,鐵肥中的鐵元素易形成難降解的氫氧化鐵,不利于作物的吸收和利用。因此,當土壤中缺鐵或鐵元素有效性較低時,為植物提供一種新形式的鐵肥以促進植物的正常生長發育是很重要的[4]。
在低鐵環境下,細菌、真菌和植物的常見策略之一是分泌鐵載體。鐵載體(Siderophore)是一種低相對分子質量且對Fe3+具有高親和力的天然鐵螯合劑[5-8]。目前,已經有500 多種鐵載體被報道過,根據其配位結構的特點,可以分為羥基鹽、兒茶酚酸鹽、羧酸鹽等類型[9]。在鐵缺乏的環境中,微生物分泌合成的鐵載體使微生物能夠獲得鐵并維持其生長。一些植物根際促生菌(PGPR)也能合成鐵載體,并且能夠與病原菌競爭獲得與鐵載體螯合的鐵,抑制病原菌的生長[10-12]。在堿性土壤環境下,許多微生物為了克服鐵有效性低的問題,自行合成和分泌高鐵載體。釋放到土壤中的高鐵載體通過配位基團的螯合反應釋放不溶性鐵(如氧化鐵、氫氧化鐵),形成鐵載體與鐵的螯合物,提高了土壤中鐵的生物有效性,從而滿足微生物生長的需要[13]。在鐵脅迫下,大麥、黑麥和小麥等能夠分泌和吸收鐵載體的植物,比玉米、高粱和水稻具有更好的選擇優勢。但植物產生的鐵載體對Fe3+的親和力較低,由植物生成的鐵載體所絡合Fe3+的穩定性低于由微生物所生成的鐵載體,其在存在其他微生物所產生的高鐵載體環境中沒有競爭力。
屬于植物根際促生菌的細菌類群達20 多個屬,根際細菌和菌根真菌互作可以提高植物的生物量,促進植物生長,改善植物及其根際土壤的養分含量,影響根際土壤微生物環境[14]。植物促生菌產生與Fe3+形成穩定螯合物的鐵載體,可以被植物直接吸收利用[15],其中最具有應用前景的是假單胞菌屬Pseudomonas和芽孢桿菌屬Bacillus。土壤微生物群落結構被認為是土壤生態系統的預警和敏感性指標之一[16]。其在維持植物的生長方面起著極其重要的作用,并且可以調節土壤養分,維持生態系統的平衡[17]。土壤微生物的空間分布和物種組成與環境條件息息相關[18],其在較大程度上受到土壤環境條件,特別是土壤理化因子的影響[19]。
本研究中主要以南疆產鐵載體細菌群落分析為切入點,進行原位土壤取樣,采用由Schwny 和Neiland 于1987年提出的CAS(chrome azurol S)法來篩選鐵載體產生菌[20],并使用16S rDNA 高通量測序,對產鐵載體細菌群落進行分析,旨在為棗樹種植過程中微生物鐵肥的研制與應用提供參考,從而提高新疆紅棗的營養品質,促進紅棗產業的健康發展。
1 材料與方法
1.1 研究區概況
選擇新疆南疆地區紅棗主產區作為研究區。研究區地勢平坦,自然坡度較小,土地平整開闊,地下水位偏低。土壤為砂壤土,pH 為8.1 ~9.2,電導率為130.57 ~1 144.33 μS/cm。土壤中堿解氮質量分數為18.43 ~238.23 mg/kg,速效磷質量分數為11.27 ~324.50 mg/kg,速效鉀質量分數為198.62 ~387.93 mg/kg,有機質質量分數為3.2 ~19.2 g/kg。土壤中有效鐵質量分數為2.12 ~30.58 mg/kg,全鐵質量分數為12.4 ~34.0 g/kg,鐵轉化率為0.011%~0.184%。
1.2 土樣采集
根據張梅等[21]對南疆紅棗適生區的研究結果,選擇在新疆和田(策勒縣、墨玉縣)、喀什(岳普湖縣、麥蓋提縣、澤普縣)、阿克蘇(阿拉爾市、沙雅縣)、巴州(若羌縣、且末縣)4 個地區9 個縣(市)的22 個棗園采集土樣(表1)。
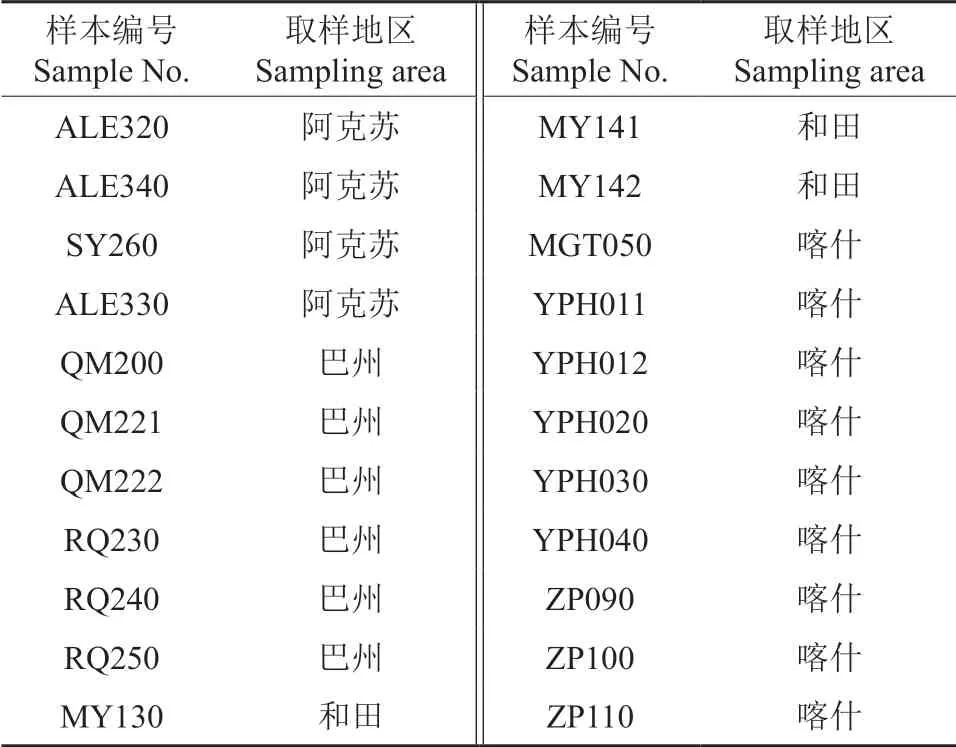
表1 南疆棗園土壤樣品來源Table 1 Source of soil samples from jujube orchards in southern Xinjiang
在棗園中選取5 棵長勢一致且樹齡相近的棗樹,用小鏟子除掉棗樹周圍表層土壤和雜物,采用三點取樣法采集土壤樣品。取30 cm 左右深處的土壤,挖取棗樹根系及其上黏附的土壤。用無菌毛刷將根上黏附的細土掃落到無菌袋中,貼上標簽。將土樣用冰盒迅速帶回實驗室,置于-26 ℃冰箱中保存備用。
1.3 試劑和培養基制備
1)CAS 溶液。取65 mg CAS 溶于50 mL去離子水中,取72.9 g 十六烷基三甲基溴化銨(HDTMA)溶于40 mL 去離子水中,將2 種溶液與10 mL 1 mmol/L 的FeCl3溶液(溶劑為10 mmol/L 鹽酸)混合,定容至100 mL,混勻,pH調至6.8,121 ℃條件下滅菌20 min,常溫避光保存。
2)牛肉膏蛋白胨液體培養基。每升培養基中加入牛肉膏3 g、蛋白胨10 g、氯化鈉5 g、瓊脂18 g,加入適量去離子水,邊加熱邊充分攪拌溶解,最后補足所失水分。121 ℃條件下滅菌25 min。
3)固體培養基基礎。取葡萄糖100 g、蛋白胨20 g、七水硫酸鎂0.5 g、氯化鈣0.5 g、瓊脂粉20 g,溶于1 000 mL 去離子水中。121 ℃條件下滅菌20 min。
4)CAS 檢測培養基。取6.04 g 無水哌嗪溶于150 mL 去離子水中,加入10×MM9 鹽溶液(Na2HPO430 g、KH2PO41.5 g、NH4Cl 5 g 溶于500 mL 去離子水中)20 mL,混勻后調節pH 至6.8±0.2,于121 ℃條件下滅菌20 min。待冷卻至60 ℃時,取100 mL 加入剛滅菌的固體培養基基礎中,再加入保溫至60 ℃的CAS 檢測液100 mL,即為CAS 檢測培養基。
5)LB(Luria-Bertani)培養基。每升培養基中加入胰蛋白胨10 g、酵母提取物5 g、NaCl 10 g,調節pH 為7.0 左右,121 ℃條件下滅菌20 min。
1.4 產鐵載體菌分析
1.4.1 菌落篩選
稱取適量新鮮土樣,加入裝有滅菌生理鹽水的錐形瓶中,在恒溫搖床中振蕩30 min(28 ℃,170 r/min)。從振蕩好的菌懸液中,吸取0.1 mL懸濁液,加入牛肉膏蛋白胨液體培養基中,在恒溫搖床中培養24 h(37 ℃,130 r/min),對菌懸液中的細菌進行活化。
吸取0.1 mL 活化后的菌懸液,均勻涂抹于CAS 檢測培養基上,置于28 ℃培養箱中培養48 h。使用接種針挑取鐵載體螯合圈半徑大于菌落直徑且長勢較好的菌落,放入盛有LB 液體培養基的錐形瓶中,在恒溫搖床中擴繁培養48 h(28 ℃,120 r/min)。吸取1 mL擴繁后的菌懸液于凍存管中,加入1 mL 無菌甘油(121 ℃滅菌25 min),搖勻后放入-26 ℃冰箱中保存,待測。
1.4.2 16S rDNA 測序
將產鐵載體細菌群落樣本送至杭州聯川生物技術股份有限公司,對序列進行PCR 擴增和16S rDNA 測序。所用引物序列見表2。

表2 南疆棗園土壤產鐵載體細菌PCR 擴增引物Table 2 Primers for PCR amplification of siderophore bacteria in jujube orchard soil of Southern Xinjiang
1.5 數據處理
使用Microsoft Office Professional Plus 2019軟件對數據進行計算和整理。基于16S rDNA 測序產生的原始數據,構建類OTU 表,獲得最終的特征表以及特征序列,采用Blast 軟件進行序列比對,用SILVA 數據庫對特征序列進行注釋,使用R 語言軟件對16S rDNA 測定數據進行分析和繪圖。采用Alpha 多樣性分析中的香農(Shannon)指數和辛普森(Simpson)指數進行種群多樣性分析[24],采用主成分分析(principal component analysis,PCA)、主坐標分析(principal coordinates analysis,PCoA)和非多維尺度分析(multidimensional scaling,NMDS)3 種方式進行種群差異分析。
2 結果與分析
2.1 南疆棗園土壤中產鐵載體細菌的群落結構
通過對經16S rDNA 測序生成的物種注釋信息進行處理,對南疆棗園土壤中產鐵載體菌群相對豐度處于前10 位的菌屬進行分析,其分布狀況如圖1所示[25]。由圖1可以看出,南疆棗園土壤中相對豐度排名在前10 位的產鐵載體細菌分屬變形菌門Proteobacteria 和厚壁菌門Firmicutes。其中,阿克蘇、和田和喀什地區棗園土壤中產鐵載體菌群以變形菌門為主,巴州地區以厚壁菌門為主。
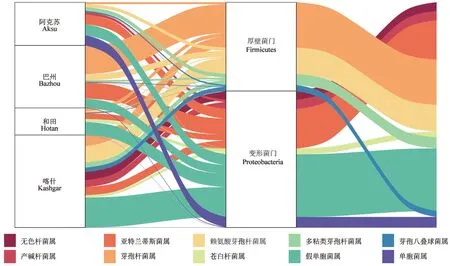
圖1 南疆棗園土壤中產鐵載體細菌(相對豐度前10 位)的菌屬分布Fig.1 Abundance of siderophores producing bacteria (top 10 abundance) in soil of jujube orchards in southern Xinjiang at phylum level and genus level
從門水平來看,4 個地區產鐵載體細菌整體上呈現出大致相同的特征。在占比最大的5 個菌屬中,芽孢桿菌屬、賴氨酸芽孢桿菌屬Lysinibacillus和多粘類芽孢桿菌屬Paenibacillus3 個菌屬為厚壁菌門,亞特蘭蒂斯菌屬Atlantibacter和假單胞菌屬2 個菌屬為變形菌門。總體上,假單胞菌屬在各地區的分布占比均相對較高,但各地區存在獨有的相對豐度較高的菌屬。巴州厚壁菌門中芽孢桿菌屬的相對豐度最高,變形菌門中亞特蘭蒂斯菌屬的相對豐度最高;喀什地區厚壁菌門中賴氨酸芽孢桿菌屬的相對豐度最高,變形菌門中假單胞菌屬的相對豐度最高;和田地區厚壁菌門中芽孢桿菌屬的相對豐度最高,變形菌門中假單胞菌屬的相對豐度最高;阿克蘇地區厚壁菌門中多粘類芽孢桿菌屬的相對豐度最高,變形菌門中假單胞菌屬的相對豐度最高。
總體上,占比最大的假單胞菌屬為變形菌門,占比次之的亞特蘭蒂斯菌屬同樣為變形菌門,與亞特蘭蒂斯菌屬占比相當的芽孢桿菌屬為厚壁菌門。
2.2 南疆棗園土壤中產鐵載體細菌群落分布的差異
根據獲得的特征值豐度,計算各樣品共有特征(feature)的數量,并通過Venn 圖直觀呈現。南疆4 個地區棗園土壤中產鐵載體細菌群落的分布特征如圖2所示。從圖2A 可以看出,4 個地區的產鐵載體細菌均有其特有的種群分布,且互有重疊。從圖2B 可以看出,總體上南疆棗園土壤中產鐵載體細菌分屬19 個門類,其中10 個門類為4 個地區所共有,巴州、和田和阿克蘇地區分別有1 個特有門類,阿克蘇與巴州有2 個共有門類、與喀什有1 個共有門類,喀什、和田和阿克蘇有1 個共有門類,喀什、阿克蘇和巴州有1 個共有門類,和田、喀什和巴州有2 個共有門類。從圖2C可以看出:4 個地區一共有230 個菌屬,其中4 個地區共有的菌屬達到了50 個;各地區均有其特有的菌屬,其中喀什特有的菌屬最多,達到了75 個,而其余地區所特有菌屬相對較少。
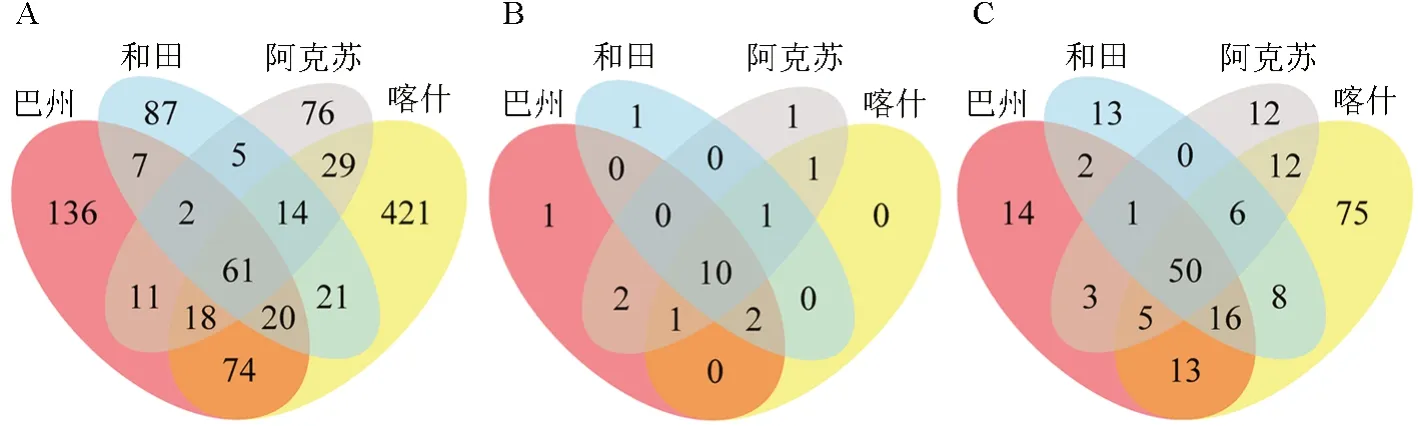
圖2 棗園土壤中產鐵載體細菌在南疆4 個地區的分布特征Fig.2 Distribution characteristics of siderophore bacteria in jujube orchard soil in four regions of southern Xinjiang
結合樣品相對豐度,根據門水平和屬水平相對豐度最高的5 個群落組成,采用布雷柯蒂斯(Bray-Curtis)距離對樣品進行聚類分析。各樣本間門水平和屬水平相對豐度處前5 位種群的聚類情況如圖3所示。從圖3可以看出,同一地區土壤樣本間產鐵載體細菌種群的相似性并非均較高,非同一地區的部分樣本的產鐵載體細菌在群落組成上存在一些相似性。RQ240 與YPH020 間、ALE320 與RQ230 間、QM200 與ZP090 間相對豐度較高的5 個門的豐度水平基本一致;SY260 和YPH030 間相對豐度較高的5 個屬的豐度水平基本一致。各地區內,各樣本的產鐵載體細菌群落總體上具有相似性,但個別樣本群落具有特異性。巴州的QM221 和RQ250、喀什地區的ZP100,在門水平上幾乎全部為厚壁菌門,在屬水平上有各自的特點。QM221 以芽孢桿菌屬為主,賴氨酸芽孢桿菌屬次之;RQ250 幾乎全部為芽孢桿菌屬;ZP100 幾乎全部為賴氨酸芽孢桿菌屬。喀什地區的YPH012 和YPH011、和田地區的MY141、阿克蘇地區的SY260,在門水平上以變形菌門為主,在屬水平上,YPH012 和MY141 以假單胞菌屬為主,其中MY141 的假單胞菌屬豐度水平比YPH012 的較低,YPH011 和SY260 以其他為主,YPH011 存在一定比例的假單胞菌屬。
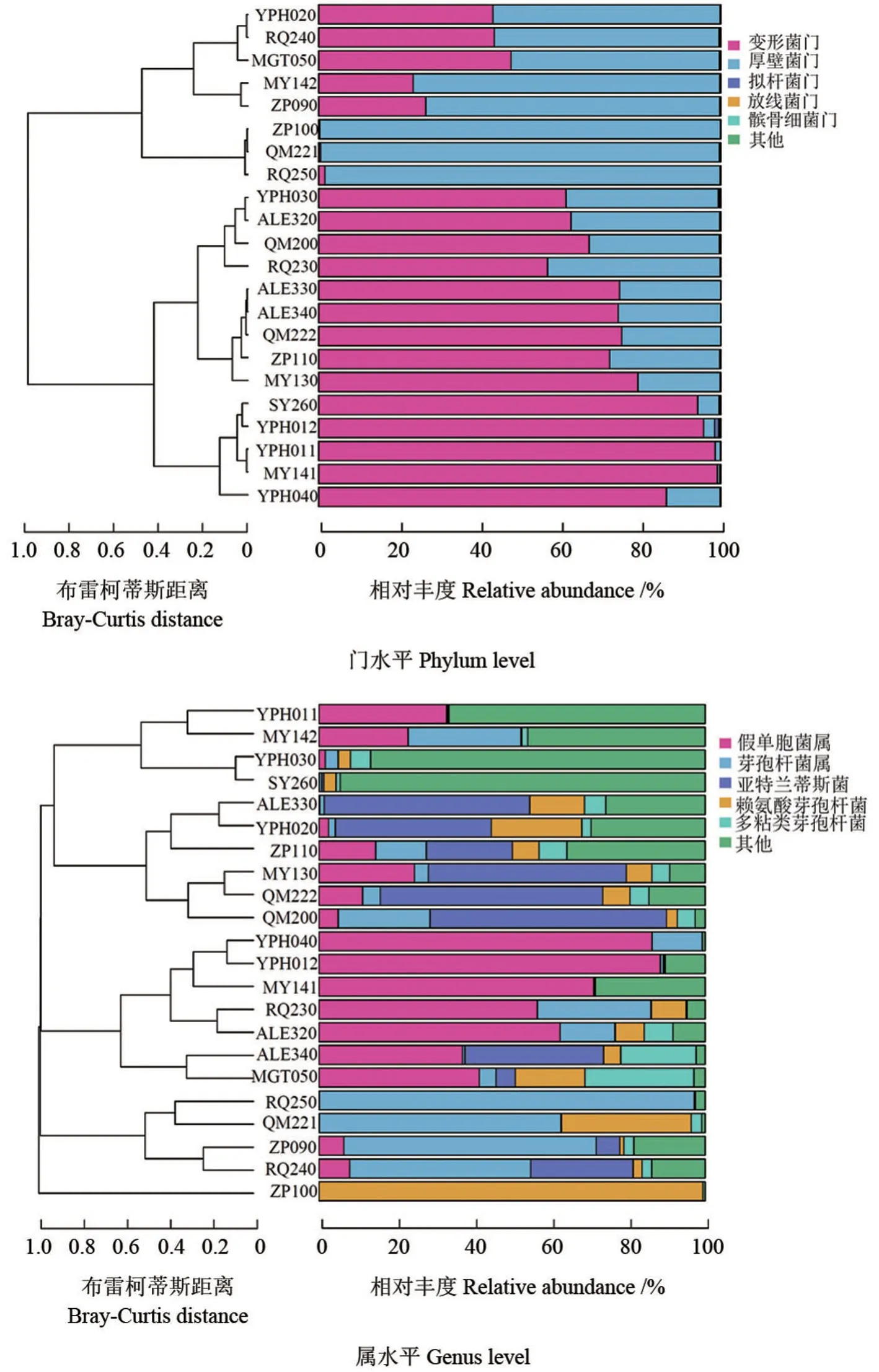
圖3 南疆棗園土壤樣本間產鐵載體細菌種群(相對豐度前5 位)的聚類Fig.3 Clustering of siderophore producing bacteria (top 5 relative abundance) among soil samples from jujube orchards in southern Xinjiang
南疆地區間棗園土壤中產鐵載體細菌種群(相對豐度前5 位)的聚類如圖4所示。從圖4可以看出:在門水平上,巴州、喀什、和田、阿克蘇4 個地區的變形菌門的相對豐度依序遞增;在屬水平上,巴州、喀什、和田、阿克蘇4 個地區的總體豐度水平較高的5 個菌屬的總豐度水平依序遞減,說明其他屬的部分菌類為變形菌門,但其豐度水平未排到前5 位。說明4 個地區的產鐵載體細菌群落在總體上具有相似性,但是也存在一定的多樣性。
同時,各地區在總體相對豐度較高的5 個門或屬的豐度水平上差異相對較大。雖然和田地區和喀什地區的產鐵載體細菌種群在門水平上較為相似,阿克蘇地區和喀什地區的產鐵載體細菌種群在屬水平上較為相似,但是從圖4可以看出,各種群的相對豐度有一定的差異。

圖4 南疆地區間棗園土壤中產鐵載體細菌種群(相對豐度前5 位)的聚類Fig.4 Clustering of siderophore bacteria (top 5 relative abundance) in jujube orchard soil in southern Xinjiang
在門水平和屬水平上的聚類結果有所不同。在門水平上,各樣本總體上可以分為4 類,屬水平上可以分為5 類。聚類分析結果表明:大多數樣本在門水平上較為相近,但在屬水平上相距較遠,或在屬水平上較為相近,但在門水平上相距較遠;不同地區的樣本在空間上距離較遠,但其產鐵載體細菌群落結構有一定的相似性。
2.3 南疆棗園土壤中產鐵載體細菌群落的多樣性
南疆棗園土壤中產鐵載體細菌種群的多樣性如圖5所示。從圖5可以看出,除喀什地區的2個樣本、和田地區的1 個樣本和巴州地區的1 個樣本的產鐵載體細菌種群多樣性較低外,4 個地區的物種多樣性均值較為一致,且除阿克蘇地區外,物種多樣性指數較為平均,且足夠豐富。
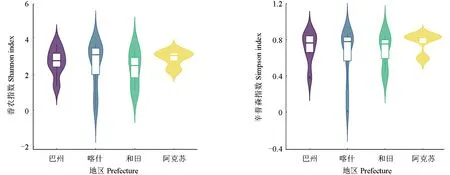
圖5 南疆棗園土壤中產鐵載體細菌種群的多樣性指數Fig.5 Diversity index of siderophore producing bacteria in soil of jujube garden in southern Xinjiang
對南疆各地區間產鐵載體細菌的種群分布進行對比,結果如圖6所示。從圖6可以看出,采用PCA、PCoA 和NMDS 這3 種分析方法得出的結果較為一致,4 個地區間產鐵載體細菌群落分布的差異不大,有較強的相似性,其中阿克蘇地區各樣點間群落分布的差異最大,和田地區各樣點間群落分布的差異最小,喀什地區各樣點間群落分布的差異不大。
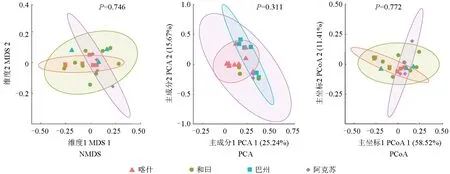
圖6 南疆地區間棗園土壤中產鐵載體細菌種群分布的差異性Fig.6 Differences in the distribution of siderophore producing bacteria in jujube orchards in southern Xinjiang
3 結論與討論
綜合以上分析結果可知,在所測定的22 份土壤樣本的產鐵載體細菌群落中,占比最大的為變形菌門的假單胞菌屬和厚壁菌門的芽孢桿菌屬,南疆各地區產鐵載體細菌群落雖然總體上呈現較為相似的構成,但仍具有多樣性,各地區保持了其獨特的群落構成。
鐵是最早被發現的植物必需微量元素之一,參與植物的多種生理過程。在新疆的堿性土壤環境中,鐵的利用效率不高,因此很有必要篩選產鐵載體微生物。通過對22 份土壤樣本中產鐵載體細菌群落的16S rDNA 測序,可以看出南疆4 個地區棗園土壤中產鐵載體細菌主要為變形菌門和厚壁菌門2 類,在屬水平上假單胞菌屬占比最大,這與Manninen 等[26]的研究結果相符。在屬水平上,還有變形菌門的亞特蘭蒂斯菌屬和厚壁菌門的芽孢桿菌屬。芽孢桿菌屬也是產鐵載體細菌的主要種群,Zhang 等[27]從小麥根際提取出多種芽孢桿菌屬的產鐵載體細菌。
通過對比可以發現,南疆4 個地區棗園土壤中產鐵載體細菌群落雖然有較強的相似性與共通性,但也保持了其特有的群落,說明不同地區土壤中產鐵載體細菌具有差異性。巴州棗園土壤中產鐵載體細菌群落與其他地區有所不同,可能是因為巴州地區的土壤情況與其他3 個地區有較為明顯的差別,且可能與巴州地區紅棗種植的管理模式有關。而且同一地區樣本的相似性并非均較高,非同一地區的部分樣本在群落組成上存在一些相似性。
大多數研究結果表明,產鐵載體細菌大多為變形菌門的假單胞菌屬和厚壁菌門的芽孢桿菌屬,因此在氣候和土壤養分條件等不同的地區,產鐵載體細菌群落可能有相似性。南疆棗園土壤中產鐵載體細菌群落的多樣性分析結果證實了上述觀點。南疆棗園土壤中產鐵載體細菌種群的多樣性分析結果表明,4 個地區的物種多樣性均值較為一致,除阿克蘇地區外,其他地區產鐵載體細菌種群的多樣性均較為豐富且均勻。通過對種群的差異性分析可以看出:阿克蘇地區各樣點間群落分布的差異最大,可能是阿拉爾市棗園與其他棗園的管理水平存在差異所導致;和田地區各樣點間群落分布的差異最小,可能是因為和田地區棗園土質較為相似,且均為干旱滴灌。雖然喀什地區的特有菌屬最多,但其樣點間群落分布的差異不大,說明同地區管理水平相近的棗園土壤中產鐵載體細菌群落較為相似。棗園的管理模式會對棗園土壤中產鐵載體細菌群落產生影響,這與吳菲等[28]的研究結果一致,人類活動的干擾會導致土壤微生物和土壤環境因子發生顯著變化。
本研究中采集了南疆紅棗主要種植區的土壤樣本進行分析,但采樣點不夠豐富,樣點分布不夠平均,使得分析結果具有一定的局限性。在后續研究中,可以擴大采樣范圍,增加采樣點位,進行更加全面深入的研究。另外,假單胞菌屬具有易于培養和產鐵載體種類豐富的特性,可以用來研制微生物鐵肥。
