圓齒野鴉椿嫁接愈合過程的顯微觀察及嫁接體酶活性的變化分析
2022-12-05 06:53:42萬瑞琪姜雪茹涂淑萍
經濟林研究 2022年3期
黃 航,劉 聰,萬瑞琪,姜雪茹,涂淑萍
(江西農業大學 林學院,江西 南昌 330045)
圓齒野鴉椿Euscaphis konishiiHayata 為省沽油科Staphyleaceae野鴉椿屬Euscaphis常綠小喬木,是我國特有的藥用與觀賞樹種[1]。該樹種的果皮于9月開始逐漸張開,露出鮮明艷麗的紅色內果皮,內果皮上附著黑色的種子,觀賞價值極高,觀果期跨越秋、冬和春(翌年)3 個季節,歷時7 個月之久。圓齒野鴉椿可叢植或列植,用于庭院、街道、風景名勝區的綠化與美化,以豐富秋、冬季園林景觀及生物多樣性,還可盆栽觀賞[2]。據報道,野鴉椿屬植物富含酚酸類、木脂素類和黃酮類等生理活性物質,具有抗腫瘤、抗菌、消炎等功效[3],其推廣應用前景十分廣闊。目前,主要采用播種繁殖技術繁育圓齒野鴉椿的苗木。但是,其種子發芽率低,且有深休眠特性[4],需經層積沙藏、熱水浸泡處理方可發芽[5];而且播種繁殖后代的性狀參差不齊,不能保持良種的優良性狀。扦插繁殖需選擇在3年以下的幼樹上剪穗扦插,而在成年樹上剪穗扦插,插穗成活率極低[6],且因受插穗的限制,繁殖系數也較低。組織培養污染率高,成本昂貴,技術要求嚴格[7]。而嫁接育苗可保持良種的優良性狀,能使其提前開花結果,增強其抗性,提高其品質,加快良種繁育速度[8-9]。因此,關于圓齒野鴉椿嫁接及愈合機理的研究,是提高良種繁育速度,實現良種化栽培的重要研究課題。前人關于嫁接愈合過程解剖學觀察[10]及酶活性的變化[11]等方面的研究報道已較多:黃蔓娜等[12]在對梨樹嫁接體愈合過程的觀察研究中將其分為隔離層產生期、愈傷組織產生期、愈傷組織分化與對接期、新輸導組織形成期4 個階段;駱建霞等[13]和Errea 等[14]的研究結果表明,砧、穗在嫁接愈合過程中的解剖結構在一定程度上能反映出嫁接愈合的親和性;此外,接口的愈合與相關酶活性密切相關,如過氧化物酶(POD)和過氧化氫酶(CAT)都是植物體內的保護酶,在一定程度上均影響著嫁接體接口的愈合[15]。而多酚氧化酶(PPO)是末端氧化酶之一,當植物受到傷害時,其易催化多酚物質形成醌,從而對植物起到保護作用;苯丙氨酸解氨酶(PAL)參與細胞分化與木質素的合成。可見,POD、CAT、PPO 和PAL 均為嫁接體愈合過程的重要評價指標[16]。目前,關于圓齒野鴉椿嫁接愈合過程的生理生化、解剖學等嫁接機理方面的研究尚未見諸報道。為給圓齒野鴉椿良種嫁接快繁育苗技術的研究提供理論參考依據,本研究以適應性強、分布范圍廣的同屬落葉樹種野鴉椿E.japonica作砧木,以觀賞價值高的常綠樹種圓齒野鴉椿作接穗,于2020年7月進行嵌芽接,對其接口愈合過程進行顯微觀察,并對嫁接體相關酶活性的變化情況進行研究,以探討圓齒野鴉椿嫁接成活的機理。
1 材料與方法
1.1 試驗材料及處理
以江西農業大學林學院花卉盆景基地(28°46′N,115°55′E)內2年生的盆栽野鴉椿實生苗為砧木,以8年生圓齒野鴉椿的當年生枝上健壯腋芽為接穗,2020年7月下旬在砧木離地約20 cm 處進行嵌芽接,于嫁接后不同時間剪取嫁接口1.5 cm 左右的莖段為試驗材料,對其進行解剖學觀察和相關酶活性的測定。嵌芽接的具體操作步驟為:先在接穗的芽上方處稍帶木質部向下斜切一刀,長約2.5 cm,再在芽下方角度為30°處斜切至第一切口底部,取下帶木質部芽片,在砧木光滑處自上而下稍帶木質部削一與接芽片大小相等的切面,隨后將芽片插入砧木切口并將兩者形成層對齊,用嫁接膜自下而上綁緊,僅將芽片上的葉柄露出即可。
1.2 試驗方法
1.2.1 圓齒野鴉椿嫁接愈合過程的解剖學觀察
于嫁接當天至嫁接后第8 天每天取樣1 次,嫁接后第8 ~22 天每隔2 d 取樣1 次,嫁接第22 天后每隔4 d 取樣1 次,至嫁接體成活為止。每次采集2 個嫁接體接口莖段,用70% 的FAA 固定液固定,置于4 ℃冰箱中保存以備用。將固定好的樣品取出,采用10%的乙二胺軟化處理45 d后,以流水沖洗2 h,以乙醇逐級脫水,每級脫水30 min,然后將脫水后的材料依次置于25%的二甲苯、50%的二甲苯、75%的二甲苯、純二甲苯、純二甲苯中進行透明處理,每次透明時間20 min,包埋,隨后將修整好的樣品置于Leica RA2235 旋轉切片機上切片,切片厚度為10 μm,展片,粘片,脫蠟,番紅固綠染色,封片,制成永久石蠟切片。最后,將切片置于德國產的ZEISS Scope.A1 顯微鏡下觀察并拍照。
1.2.2 圓齒野鴉椿嫁接愈合過程嫁接體相關酶活性的測定
于嫁接當天開始取樣,每隔4 d 取樣1 次,每次隨機選取3 ~4 個莖段,重復3 次。將樣品取下后立即置于液氮罐中速凍,帶回實驗室置于-80℃的超低溫冰箱中保存以備用。
采用鉬酸銨比色法測定過氧化氫酶(CAT)活性,采用分光光度法測定過氧化物酶(POD)、多酚氧化酶(PPO)和苯丙氨酸解氨酶(PAL)的活性;所用試劑盒均由蘇州科銘生物技術有限公司生產。
1.3 數據處理
使用Excel 2010 軟件對數據進行處理并繪圖,利用Photoshop CS5 軟件對照片進行處理,采用SPSS 17.0 軟件對數據進行統計分析。
2 結果與分析
2.1 圓齒野鴉椿嫁接愈合過程的解剖學觀察結果
以野鴉椿為砧木嫁接圓齒野鴉椿后,對其嫁接體的愈合過程進行顯微觀察,結果表明,其愈合過程可劃分為3 個時期,即砧穗切口隔離層及愈傷組織形成期、砧穗結合部愈傷組織增殖與連接期、砧穗結合部愈傷組織分化形成維管組織期。
2.1.1 砧穗切口隔離層及愈傷組織形成期
砧穗切口隔離層及愈傷組織形成的過程如圖1所示。從圖1A 中可以看出,嫁接后第3 天在砧木的切面上可見一層染色較深的薄層,即為隔離層,而接穗的切面直到嫁接后第6 天才可觀察到隔離層(圖1C),說明砧木比接穗提早3 d 形成隔離層,早期形成的隔離層對傷口具有一定的保護作用。從圖1B 中可以看出,嫁接后第5 天,砧木切口周圍的部分形成層、木質部、木射線等部位的細胞開始脫分化形成愈傷組織。此時,橫切面上砧木木質部薄壁細胞、木射線脫分化的細胞徑向開始增厚并向外凸起,薄壁細胞明顯增加,無規則地堆積在一起,部分薄壁細胞體積較大,清晰可見內含較大的細胞核,隔離層清晰可見。從圖1D 中可以看出,嫁接后第7 天,砧木皮層、韌皮部薄壁細胞突破了隔離層,經脫分化形成愈傷組織,并于其四周大量分裂增殖,形成愈傷組織細胞團,多數細胞呈無序排列狀。而接穗切面于嫁接后第10 天在皮層和韌皮部處開始形成愈傷組織(圖1E)。可見,接穗較砧木晚5 d 形成愈傷組織。
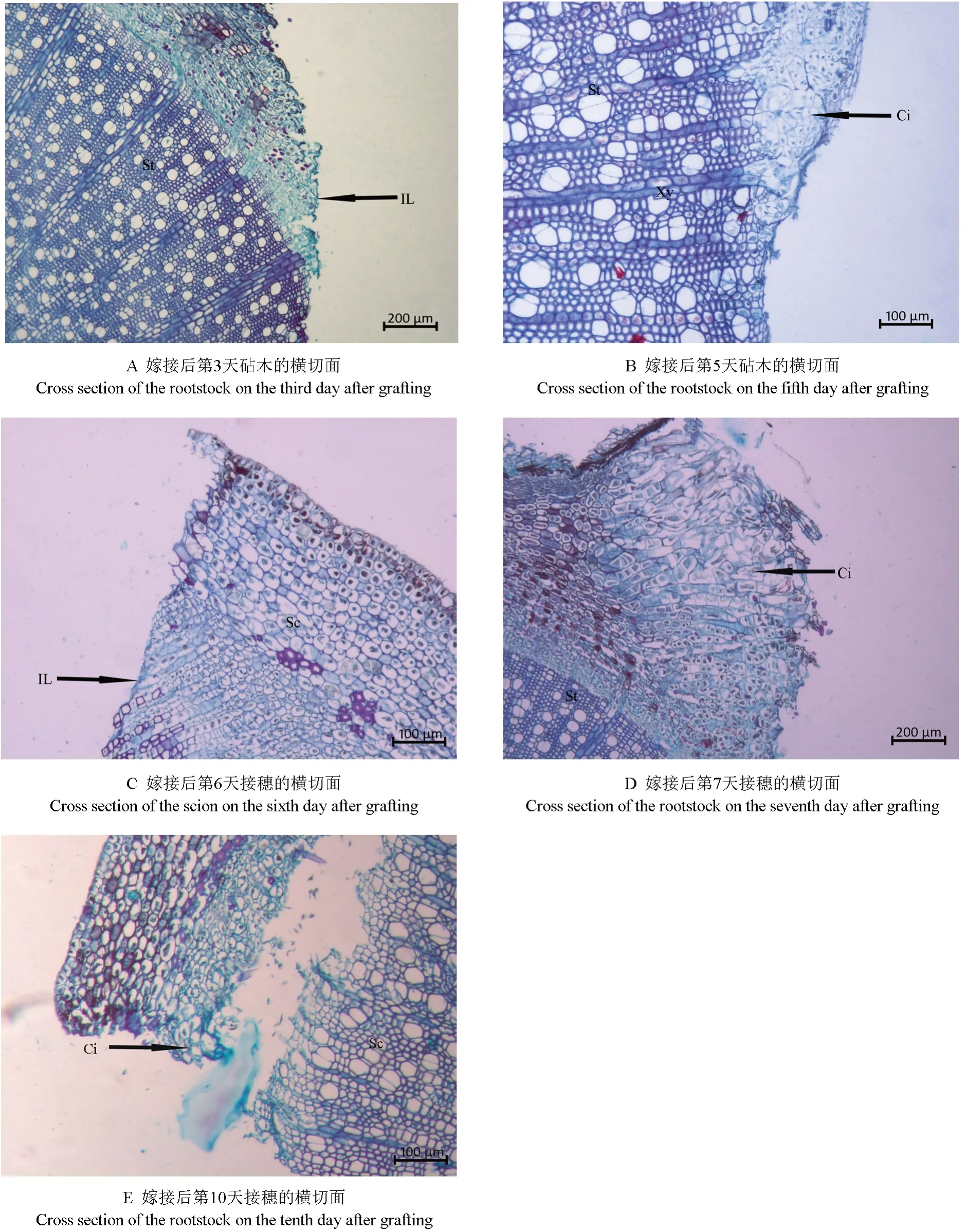
圖1 隔離層及愈傷組織形成過程的顯微觀察結果Fig.1 Observation on the microstructure of isolation layer and callus period
2.1.2 砧穗結合部愈傷組織增殖與連接期
砧穗結合部愈傷組織增殖與連接的過程如圖2所示。從圖2A 中可以看出,嫁接后第14 天,砧木和接穗間的空隙因愈傷組織的抱合和連接而被填滿,二者緊密結合形成愈傷組織橋,而隔離層已消失。此時,愈傷組織橋為砧穗間細胞流通的主要途徑。嫁接后第20 天,砧穗結合部構成形成層環,表明嫁接體已經愈合(圖2B)。

圖2 愈傷組織增殖與連接過程的顯微觀察結果Fig.2 Observation on the microstructure of callus proliferation and connection period
2.1.3 砧穗結合部愈傷組織分化形成維管組織期
砧穗結合部愈傷組織分化形成維管組織的過程如圖3所示。由圖3A 可以看出,嫁接后第26 天,嫁接體形成層的細胞分化出較多結構規整的木質部和韌皮部,較大口徑的導管已清晰可見,此時嫁接體已經形成了一個新的個體。嫁接后第40 天,新分化出的維管束擴大分化,維管束呈現出不規整的分布狀,形成維管束混合群體(圖3B)。
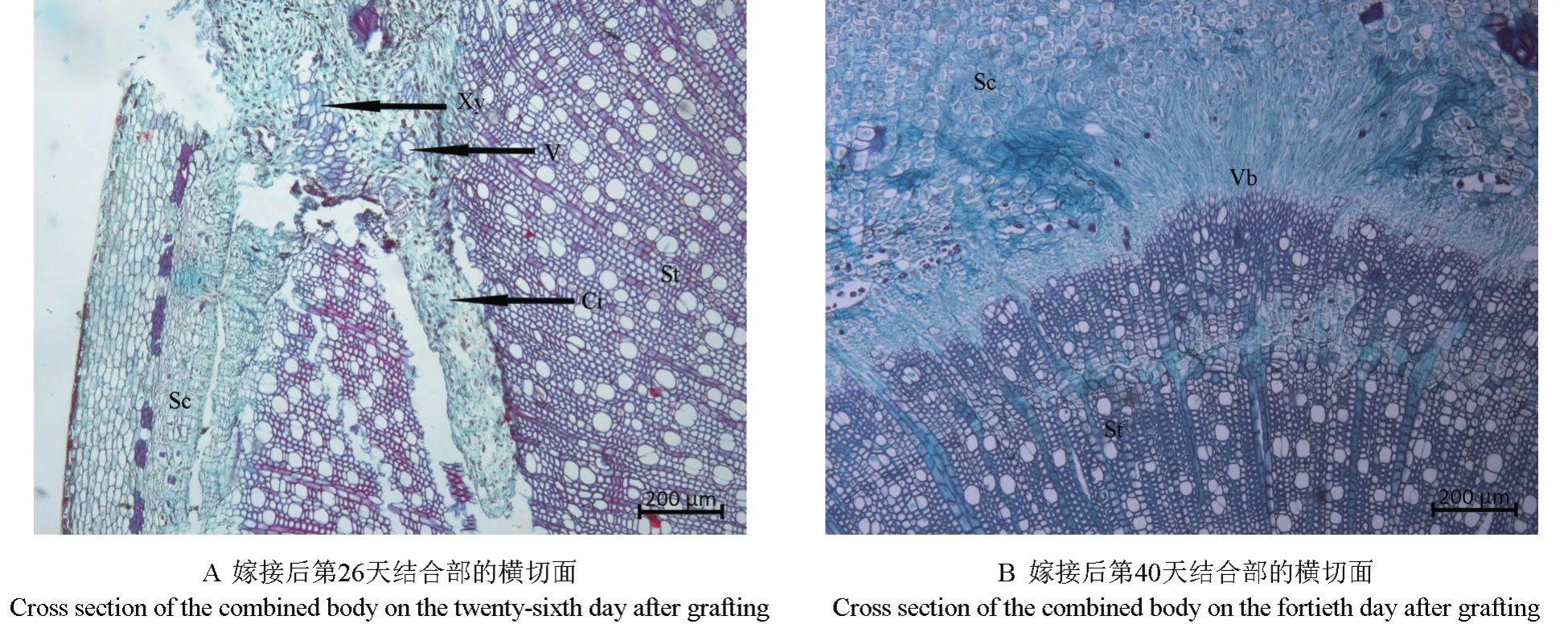
圖3 愈傷組織分化形成維管組織過程的顯微觀察結果Fig.3 Observation on the microstructure of callus during differentiation into vascular tissue
2.2 圓齒野鴉椿嫁接愈合過程中嫁接體酶活性的變化趨勢
2.2.1 過氧化氫酶和過氧化物酶活性的變化趨勢
嫁接口愈合過程中嫁接體內過氧化氫酶(CAT)和過氧化物酶(POD)活性的變化趨勢如圖4所示。嫁接當天至嫁接后第20 天,嫁接體內CAT 和POD 活性的變化趨勢基本一致。在嫁接后第10 天之前即砧穗切口隔離層和愈傷組織形成期,CAT 和POD 活性均顯著上升,其中,CAT 活性在嫁接第10 天達到峰值。在嫁接后第10 ~15 天即嫁接口愈傷組織細胞分裂增殖期,CAT 和POD 活性均顯著下降。在嫁接后第15 ~20 天即砧穗愈傷組織連接愈合期,CAT 和POD 活性均顯著上升。在嫁接后第20 ~25 天即砧穗結合部愈傷組織進行新生維管組織分化期,CAT 和POD 活性開始出現不一致的變化情況,即CAT 活性下降,而POD活性上升。

圖4 嫁接體內過氧化氫酶和過氧化物酶活性的變化趨勢Fig.4 The change trend of CAT and POD activities in graft
2.2.2 多酚氧化酶和苯丙氨酸解氨酶活性的變化趨勢
嫁接口愈合過程中嫁接體內多酚氧化酶(PPO)和苯丙氨酸解氨酶(PAL)活性的變化趨勢如圖5所示。在嫁接口的愈合過程中,PPO 和PAL 活性的變化趨勢正好相反。在嫁接后第10 天之前即砧穗切口隔離層和愈傷組織形成期,PPO 活性顯著升高,而PAL 活性顯著降低;在嫁接后第10 ~15 天即砧穗結合部愈傷組織細胞分裂增殖期,PPO 活性顯著降低,而PAL 活性升高;在嫁接后第15 ~20 天即砧穗結合部愈傷組織連接愈合期,PPO 活性較前稍有降低,而PAL 活性顯著升高;在嫁接后第20 ~25 天即砧穗結合部愈傷組織分化形成維管組織期,PPO 活性升高而PAL活性降低。

圖5 嫁接體內多酚氧化酶和苯丙氨酸解氨酶活性的變化趨勢Fig.5 The change trend of PPO and PAL activities in graft
3 討 論
3.1 嫁接體接口愈合過程的解剖學觀察結果分析
嫁接成活過程實質上是砧、穗結合部位通過傷口產生愈傷組織、形成愈傷組織橋和形成層環,分化維管束組織,形成一個新個體的過程[17]。初慶剛等[10]將梨樹嫁接愈合過程劃分為愈傷組織的形成期和維管形成層的形成及活動期這2 個時期;兗攀等[18]將美國山核桃嫁接愈合過程劃分為砧穗隔離期、愈傷組織生長期、形成層環構形成期和新生維管組織分化期這4 個時期。不同物種嫁接愈合的組織學特征存在差異,而其基本結構及愈合過程卻大致相同。擬南芥嫁接后第3 天即分化出大量維管束結構[19];早實核桃芽接后第6 天砧穗維管束就已重新連接[20];黃瓜嫁接后第15 天維管束完全建成[21];梨樹嫁接后第20 天新的輸導組織就已形成[10];油茶芽苗砧嫁接后第35 天愈傷組織維管束才形成[22]。綜上所述,物種不同,嫁接體木質化程度不同,嫁接成活的快慢則不一致。顯微觀察結果表明,以‘野鴉椿’為砧木嫁接‘圓齒野鴉椿’后,嫁接體在嫁接后第26 天已形成了新的導管和篩管,使砧穗間能夠正常進行養分和水分的輸送,并形成了一個新的個體。
嫁接時,砧、穗切口細胞受到機械損傷,細胞的內含物與殘壁發生氧化反應,形成高分子物質,隔離層覆蓋整個嫁接面[22]。植物遭受創傷后易形成隔離層,嫁接初期隔離層能有效防止水分的蒸發,保護傷口不被病原體入侵,但是,隔離層太厚也會影響砧穗間愈傷組織的形成[11]。顯微觀察結果還表明,切口處開始出現隔離層與愈傷組織的時間,砧木比接穗分別早3 和5 d,砧木早于接穗恢復生理活動,故有利于嫁接體的愈合成活。這一觀察結果與楊志堅等[22]的研究結果一致。其中,砧木切口處隔離層與愈傷組織開始形成的時間分別為嫁接后第3 天和第5 天,與接穗切口處隔離層與愈傷組織開始形成的時間相比,分別早3 和5 d。隨著嫁接體的發育,嫁接第5 ~10 天,砧穗切口處部分細胞突破隔離層,形成愈傷組織,愈傷組織細胞在分化過程中不斷生長。
嫁接后第10 ~20 天,為砧、穗切口處愈傷組織細胞分裂增殖期;嫁接后第14 ~20 天,為砧、穗愈傷組織連接愈合期;嫁接后第14 天,砧、穗間的空隙被愈傷組織填滿,嫁接體形成愈傷組織橋,隔離層消失;嫁接后第20 天,砧穗結合部構成形成層環,嫁接體完成愈合過程。兗攀等[18]觀察發現,愈傷組織分化形成愈傷組織橋和形成層環,并隨即產生新的輸導組織(如韌皮部、木質部),砧、穗便接合成一個整體。兗攀等[18]的觀察結果與本研究結果一致。
嫁接愈合過程中,嫁接體的顯微結構在一定程度上可以反映砧、穗嫁接的親和性[13]。嫁接后第21 ~26 天,為嫁接體愈傷組織分化形成新生維管組織期。嫁接后第26 天,砧穗結合部形成層的細胞已經分化出新的導管和篩管,標志著嫁接體已成活,并已形成一個完全獨立的新個體。從外觀上看,此時接芽萌發,砧穗緊密連接,這與砧穗結合部愈傷組織分化形成維管組織期的顯微觀察結果相吻合,表明野鴉椿與圓齒野鴉椿均有較強的嫁接親和性。因此,嫁接后第26 天之前是圓齒野鴉椿嫁接成活的關鍵時期。
王淑英等[23]在觀察葡萄嫁接愈合過程時發現,形成層的細胞是愈傷組織產生的唯一來源。但是,楊志堅等[22]在觀察油茶芽苗砧嫁接口愈合過程時發現,木本植物愈傷組織的來源不僅限于形成層的細胞,還包括髓區、木射線、皮層的細胞。周乃富等[24]的研究結果表明,核桃嫁接愈合方式不僅存在由形成層細胞主導的砧穗愈合方式,還存在由木射線細胞主導的新方式。顯微觀察發現,嫁接初期砧穗傷口處皮層、韌皮部薄壁細胞和木射線及形成層的細胞均可產生愈傷組織,但是,形成層的細胞在嫁接愈合過程中自始至終占主導地位。因此,在圓齒野鴉椿嫁接過程中,創造有利于愈傷組織生長和新的形成層產生的條件,是提高嫁接成活率的重要途徑。
3.2 嫁接體接口部位抗氧化酶活性的變化趨勢
過氧化物酶(POD)和過氧化氫酶(CAT)是植物體內重要的抗氧化酶類,具有清除接口處過量活性氧(ROS)、促進木質素合成的作用,有利于嫁接體接口的愈合[15]。有關研究結果表明,與非親和性的黃瓜、甜瓜嫁接苗相比,親和性的黃瓜、甜瓜嫁接苗接口處均有較高的抗氧化酶(SOD、POD、APX)活性和較低的活性氧含量[25]。胡艷青等[11]的研究結果表明,嫁接初期,CAT 活性升高,有利于清除氧自由基,起到防止過氧化傷害和清除植物體內過剩的H202的作用;嫁接體成活后,CAT 活性隨之下降。試驗結果表明,嫁接后第0 ~10 天,以野鴉椿為砧木、以圓齒野鴉椿為接穗進行嵌芽接,其嫁接體的CAT 活性顯著升高,而此時即為解剖學上砧穗切口隔離層與愈傷組織的形成時期,說明嫁接體在初期受到創傷后,嫁接苗的抗氧化能力得到提升。嫁接后第20 天,即砧穗結合部愈傷組織分化形成新生維管組織時期,CAT 活性開始降低,此時嫁接體已經愈合,形成層細胞分化出較多結構規整的木質部和韌皮部,新的輸導組織已經形成,結合部 CAT 活性呈下降趨勢,這與上述研究結果基本一致。但是,曲云峰[26]研究發現,CAT 不是杏嫁接成活的關鍵酶,具體原因有待進一步研究。
盧善發[27]的研究結果表明,POD 是氧化吲哚丁酸的關鍵酶,嫁接初期POD 活性升高,有利于維持激素的平衡;王瑞敏等[28]研究了金葉銀杏的扦插生根機制,結果表明,高活性的POD 能夠促進插穗愈傷組織的形成;張紅梅等[29]研究發現,嫁接愈合過程中,POD 活性呈升—降—升的變化趨勢,POD 活性高有利于維管組織的木質化。試驗結果還表明,嫁接初期,嫁接體內的POD 活性顯著升高,并于嫁接后第10 天出現第1 個高峰值,此時即為愈傷組織形成時期,解剖學觀察結果顯示,嫁接第5 天后,砧穗愈傷組織大量分化,由此可以推測,嫁接后第5 ~10 天是愈傷組織形成和生長的高峰期;隨著嫁接愈傷組織的分裂增殖,POD 活性顯著降低;嫁接后第15 ~25 天,嫁接體內的POD 活性又顯著升高;嫁接后第26 天,嫁接體接合部位的POD 活性最高,此時正是愈傷組織分化形成維管組織時期,POD 的活性高,表明其參與了維管組織的分化。綜上所述,嫁接體結合部的POD 和CAT 活性均高,則有利于嫁接口的愈合,這與上述試驗結果類似。
3.3 嫁接體接口部位苯丙烷代謝相關酶活性的變化趨勢
苯丙氨酸解氨酶(PAL)和多酚氧化酶(PPO)為苯丙烷代謝相關酶,在嫁接體愈傷組織細胞形成、增殖及分化過程中發揮了重要作用[30]。
Chen 等[31]研究發現,嫁接接口處PPO 活性升高,有利于嫁接體的愈合與生長;López-Gómez等[32]的研究結果表明,PPO 主要作用于嫁接初期,表現為嫁接初期PPO 活性上升。試驗結果表明,嫁接后第0 ~10 天即隔離層與愈傷組織形成時期,PPO 活性快速升高,形成了足夠厚的隔離層,可以保護嫁接體免受感染,這與上述研究結果一致。但是,亦有研究結果表明,PPO 活性升高加速了接口處木質化的進程,從而使嫁接難以成活[33]。嫁接后第10 天,嫁接體內PPO 活性迅速下降并趨于平穩,而此時正是愈傷組織增殖與連接期,隔離層的消失有利于嫁接口的愈合,這一結果與張淑紅等[34]的研究結果類似。
劉婧冉等[16]的研究結果表明,PAL 參與嫁接愈合過程中細胞的分化,有利于嫁接體組織的發育和功能的重建。馮金玲等[35]對油茶嫁接的試驗結果表明,在嫁接初期即愈傷組織形成時期,PAL活性呈下降趨勢;而在嫁接愈合完成后,PAL 活性卻上升。嫁接試驗中發現,在嫁接初期即愈傷組織形成期,PAL 活性降低;而在愈傷組織細胞分裂增殖與連接期,PAL 活性升高。這一結果表明,此時砧、穗間細胞分化旺盛,有利于木質部的形成,這與上述研究結果類似,但與黃蔓娜等[12]在對梨樹進行嫁接試驗時發現的嫁接后第18 天砧、穗形成層連接后其PAL 活性快速增加的結果不同。
試驗結果表明,PPO 和PAL 酶活性在嫁接體愈合過程中的變化趨勢正好相反。嫁接初期,PPO活性升高,而PAL 活性降低,這有利于嫁接體接口愈傷組織的形成。
目前,關于圓齒野鴉椿嫁接愈合過程的生理生化機制、解剖學等嫁接機理方面的研究報道較為鮮見。本研究對圓齒野鴉椿嫁接愈合過程進行了解剖學觀察,并分析了嫁接體的生理生化特征,但因受場地、苗量等因素的影響,未設置自根苗與嫁接苗的酶活性的比較試驗,故試驗結果存在一定的局限性;對嫁接愈合過程的細胞學觀察,還需結合嫁接接口愈合分子機制、基因組學和蛋白組學等方面的研究結果來分析,以優化并完善圓齒野鴉椿嫁接繁殖技術體系。
4 結 論
本研究以野鴉椿實生苗為砧木、以圓齒野鴉椿當年生枝上健壯的腋芽為接穗進行嵌芽接試驗,觀察了砧穗愈合過程的解剖學形態特征,測定并分析了嫁接后不同時期其結合部的相關酶活性的變化情況,得出了如下結論。
1)嫁接體的愈合過程可劃分為3 個時期,即砧穗切口隔離層及愈傷組織形成期(嫁接后第0 ~10 天)、砧穗結合部愈傷組織增殖與連接期(嫁接后第11 ~20 天)、砧穗結合部愈傷組織分化形成維管組織期(嫁接后第21 ~ 26 天)。
2)嫁接體在嫁接后第26 天形成了新的導管和篩管,砧穗間細胞能夠正常接觸和流通,輸送養分和水分,形成了一個新的個體,說明野鴉椿與圓齒野鴉椿均有較強的嫁接親和性。
3)嫁接愈合過程中,嫁接體內酶活性的變化趨勢為:嫁接初期,POD、CAT 和PPO 活性均升高而PAL 活性降低,這有利于嫁接體接口的愈合;而嫁接體愈合后,PAL 活性降低而PPO 和POD 活性均升高,這可促進嫁接體維管組織的形成。
