鹽脅迫下園林植物彩葉樹響應菌根共生的比較轉錄組分析
2022-12-13 01:14:10溫可馨
江蘇農業(yè)科學 2022年22期
陳 蓋, 溫可馨, 司 冰
(1.唐山工業(yè)職業(yè)技術學院,河北唐山 063299; 2.渭南職業(yè)技術學院,陜西渭南 714026)
土壤是植物生長發(fā)育的重要載體,然而全球范圍內超過7%的土地受到不同程度鹽脅迫的影響[1]。鹽脅迫是對作物生長發(fā)育及產量形成最不利的環(huán)境脅迫因子之一,其潛在的分子機制與多種生物途徑和過程有關,包括滲透調節(jié)、離子泵、氧化途徑以及營養(yǎng)障礙等代謝過程的改變[2];分子水平上,鹽脅迫可限制細胞分裂、DNA擴增,甚至誘導產生基因毒性[3]。此前的研究發(fā)現,植物體具有多種信號轉導通路協助植物適應鹽脅迫,例如鹽脅迫敏感系統(tǒng)(SOS)、絲裂原活化蛋白激酶(MAPK)級聯、磷脂酰肌醇(PI)和脫落酸(ABA)介導的信號轉導通路[4]。此外,為應對鹽脅迫,分子信號傳導可以激活其他轉錄因子(TF),TF可增強和激活基因調控信號,從而重新調節(jié)細胞離子、滲透和活性氧(ROS)的穩(wěn)態(tài)[5]。
叢枝菌根(arbuscular mycorrhizal,AM)真菌是一種重要的土壤有益微生物,它可與80%以上的陸生植物形成互惠互利的共生關系,這種共生關系可促進植物生長并改變植物的形態(tài)、生理和營養(yǎng)水平[6],從而提高對多種非生物脅迫因素的抵抗力,包括養(yǎng)分貧瘠、高溫、寒冷、干旱和鹽脅迫[7]。在鹽脅迫條件下,接種AM真菌的植物可以增強對離子穩(wěn)態(tài)、滲透平衡和抗氧化酶活性的調節(jié)[8],同時也可提高光合活性以改善養(yǎng)分活化、營養(yǎng)獲取和植物的水分吸收[7]。然而,以往對鹽脅迫環(huán)境改善機制的研究主要集中于形態(tài)學和生理學特征,很少對接種AM真菌的植物在基因組范圍內的基因表達進行探索,這限制了AM真菌在植物抗鹽性方面的應用認知[9],因此,須要對鹽脅迫條件下AM真菌改善機制進行進一步的分子水平分析。
彩葉樹是指在整個生長季節(jié),全部或部分葉色與自然綠色有明顯區(qū)別的木本植物類群,基本特征是具備一致的變色期、較長的觀賞期和整齊的落葉期[10]。目前,紫葉稠李、金葉女貞、雞爪槭、五角楓、金葉紅瑞木等10余種彩葉樹種廣泛引種栽植于我國各省市道路、街道兩側及公園中,是園林綠化的重要組成[11]。然而在實地栽種中,在各個地區(qū)均呈現出植株生長勢弱、適應性差、病蟲害嚴重等現象。許多彩葉樹在鹽漬土壤中生長不良,表現出葉片焦枯、葉色灰暗等現象[12]。在前人的研究中,接種AM真菌可有效調節(jié)100 mmol/L鹽脅迫下彩葉樹的滲透脅迫、氧化損傷和離子應力,從而促進植株的生長發(fā)育[13],這些結果為進一步研究彩葉樹接種AM真菌的耐鹽性分子機制奠定了基礎。本研究中,借助Illumina HiSeqTM2500測序平臺分析彩葉樹雞爪槭(Acerpalmatum)葉片組織中的全基因轉錄變化,以探索AM真菌誘導耐鹽性的分子機制。研究結果可為更全面地了解AM真菌在耐鹽性中的作用提供理論支持,并啟動有效的基因工程策略。
1 材料和方法
1.1 供試時間與供試材料
試驗于2021年5—8月于渭南職業(yè)技術學院試驗場大棚中進行。供試彩葉樹品種為雞爪槭,購自楊凌益恒園林綠化有限公司。種子采用表面滅菌后在培養(yǎng)皿中的3層濕濾紙上,在 26 ℃ 環(huán)境下黑暗處理催芽4 d。
供試培養(yǎng)基質為體積比為2 ∶1的土壤與珍珠巖(直徑<3 mm),高壓蒸汽滅菌(121 ℃,1×105kPa,4 h)后,冷卻備用。土壤理化性質:pH值為7.11,有機質含量為18.32 g/kg,全氮含量為 0.95 g/kg,堿解氮含量為78.26 mg/kg,速效磷含量為21.03 mg/kg,速效鉀含量為110.64 mg/kg。供試NaCl為優(yōu)級純,購自上海玉博生物科技有限公司。
供試AM真菌為根內根孢囊霉(Rhizophagusintraradices),購自北京市農林科學院植物營養(yǎng)與資源環(huán)境研究所,直接用作接種菌劑,接種菌劑由土壤和孢子構成,孢子數>100個/g基質。
1.2 試驗設計
試驗采用完全隨機設計,設置4個處理:CK(不接種叢枝菌根真菌,不設置鹽脅迫處理)、AM(接種叢枝菌根真菌,不設置鹽脅迫處理)、SS(不接種叢枝菌根真菌,設置鹽脅迫)、AS(接種叢枝菌根真菌,設置鹽脅迫)。每個處理重復3次。
盆栽器具為桶形,盆高為17 cm,半徑為8 cm,每盆培養(yǎng)基質5 kg。將催芽的種子每盆1粒轉移至盆栽土中,加入200 mL育苗專用營養(yǎng)液,并保持60%土壤持水量。接種AM真菌的處理須將20 g AM真菌菌劑與滅菌的培養(yǎng)基質混合,不接種處理即采用經高溫滅菌處理的菌劑等量加入。培養(yǎng)30 d后在鹽脅迫處理中加入100 mL濃度為100 mmol/L的NaCl溶液,對照中加入等量蒸餾水。脅迫處理后30 d收獲植株,共培養(yǎng)60 d。
1.3 樣品采集及測定分析
1.3.1 生物量、菌絲定殖率、叢枝侵染率及泡囊豐富度測定 培養(yǎng)結束后,將植株全部取出,小心清洗以獲得完整根系,將根系切成1 cm的小段,采用臺盼藍染色,光學顯微鏡下用網格交叉記數法計算菌絲定殖率、叢枝定殖率及泡囊豐富度,具體方法參照Kormanik等描述的步驟[14]進行。生物量為鮮質量,采用電子天平稱量記錄。
1.3.2 轉錄組測定
1.3.2.1 RNA提取、cDNA文庫構建和高通量測序 使用RNA prep Pure Plant Kit[天根生化科技(北京)有限公司]分離12(4×3)張葉片樣品的總RNA。文庫構建和RNA-Seq分析由Biomarker Biotechnology Corporation(中國,北京)進行。首先,使用NEBNext? poly(A) mRNA Magnetic Isolation Module[紐英倫生物技術(北京)有限公司]從總RNA中富集提取poly(A) mRNA,將poly(A) mRNA片段化。這些中斷的片段被用作第一鏈和第二鏈cDNA 合成的模板。然后對得到的cDNA進行末端修復、磷酸化和連接到測序接頭;通過PCR擴增合適的產物以構建cDNA文庫。最后,使用Illumina HiSeqTM2500對12個純化的文庫進行高通量測序。
1.3.2.2 高通量測序RNA-Seq讀取映射和功能注釋 進行RNA-Seq分析后,通過修剪接頭并去除低質量的原始序列以獲得高質量的讀數序列。使用HISAT2軟件將讀數映射到雞爪槭參考基因組(GCA_001876935.1,Acerpalmatum2017)。使用StringTie軟件對映射的讀數進行組裝和定量分析,作為每千個堿基的轉錄每百萬個片段映射(FPKM)值。來自參考基因組的原始基因的非翻譯區(qū)(UTR),在連續(xù)映射讀取的基礎下,擴展到上游和下游以合成和優(yōu)化基因結構[15]。此外,通過比較基因組的原始注釋信息,進行一一對比找到以前未注釋的轉錄區(qū)域(即過濾長度大于50個氨基酸的短多肽產物和單個外顯子區(qū)域后的新基因)。對于新基因,對潛在的新轉錄本進行功能注釋,即將測序得到的轉錄本與公開可用的蛋白質數據庫(Nr、Swiss-Prot、GO、COGs、KOG、Pfam、egg-NOG和KEGG)進行比對,使用BLASTx算法獲取對比值。
1.3.2.3 差異表達基因(DEGs)分析 每個基因的表達水平由每千個堿基外顯子每百萬個片段映射值決定。使用DESeq軟件進行成對比較中的差異表達基因分析。通過計算所有基因的錯誤發(fā)現率(FDR)和倍數變化對數(log2C)。結合GO注釋結果進行功能描述,接著對DEGs進行KEGG Pathway富集分析,結合KEGG注釋結果獲取差異基因相關的生化代謝途徑及生物信號轉導途徑。當轉錄本符合FDR≤0.01和log2C的絕對值≥1 時,認為該轉錄本存在功能富集。轉錄組測序委托上海美吉生物醫(yī)藥科技有限公司完成。
1.3.3 實時定量PCR驗證差異基因準確性 將保存于-80 ℃的葉片樣品快速研磨,采用轉基因Ⅱ一體化引物試劑盒TransScript Ⅱ All-in-One First-Strand cDNA Synthesis SuperMix for qPCR(北京全式金生物技術股份有限公司)提取樣本總RNA,并將總RNA轉化為單鏈cRNA。采用Primer Premier 5.0軟件設計基因特異性引物(表1)。使用TransStart Top Green qPCR SuperMix(北京全式金生物技術股份有限公司)進行實時熒光定量PCR(qRT-PCR),并在Bio-Rad CFX96實時PCR檢測系統(tǒng)(Bio-Rad,USA)上1式3份進行PCR定量復孔檢測。
每個反應包含10 μL 2×TransStart Top Green qPCR SuperMix、2.0 μL cDNA樣本、0.4 μL基因特異性引物、7.6 μL ddH2O,最終體積為20 μL。利用比較的方法計算每個樣本的mRNA與內參基因AcerPM1的相對表達量,以2-ΔΔCT表示[16]。最后,通過 qRT-PCR和RNA-Seq分析比較SS、AS處理中基因表達的變化趨勢,以評估測序結果是否可靠。基因驗證委托上海美吉生物醫(yī)藥科技有限公司完成。

表1 qRT-PCR 引物序列信息
1.4 數據處理與統(tǒng)計分析
采用Excel 2013進行初步數據整理,采用SPSS 23.0軟件中的單因素方差分析進行統(tǒng)計檢測分析(α=0.05),所有圖形均采用Origin 8、R語言軟件繪制。
2 結果與分析
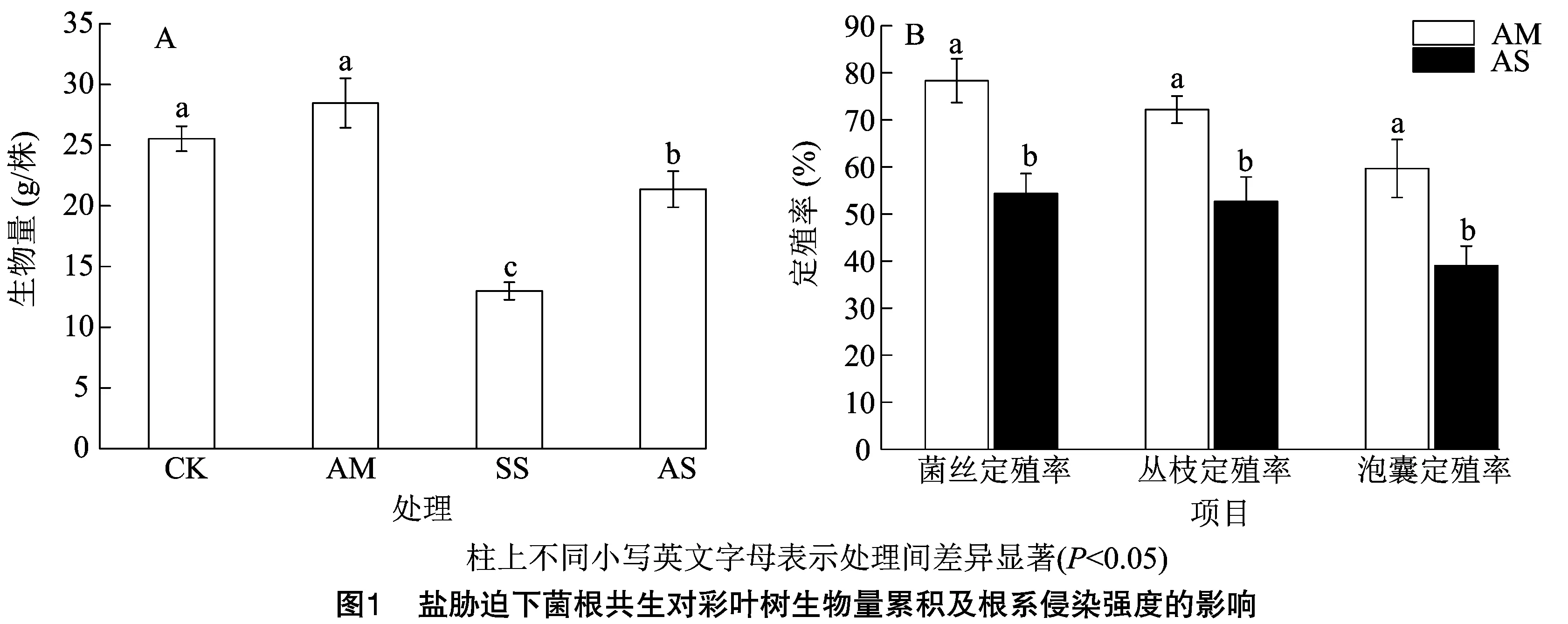
2.1 鹽脅迫下菌根共生對彩葉樹生物量累積及根系侵染強度的影響
由圖1-A可知,在生物量中,以AM處理生物量累積最高,SS處理生物量累積最低,各處理表現為AM>CK>AS>SS,其中AM與CK處理無顯著差異,二者皆顯著大于SS和AS處理,且SS處理顯著小于AS處理;與SS處理相比,CK、AM、AS處理分別顯著提高104.31%、119.17%、64.51%。由圖1-B可知,在沒有接種AM真菌的處理中沒有定殖情況,因此未列出;接種叢枝菌根真菌情況下,整體表現為菌絲定殖率大于叢枝定殖率,且以泡囊定殖率最低。在AM處理中,菌絲、叢枝及泡囊的定殖率分別為78.33%、72.16%、59.67%,鹽處理(AS)下,菌絲、叢枝及泡囊的定殖率分別為54.33%、52.67%及39.05%;且在任一定殖率指標中,皆以AM處理顯著大于AS處理。
2.2 鹽脅迫及菌根共生下彩葉樹轉錄本的原始數據分析
為了全面了解AM真菌對鹽脅迫下彩葉樹葉片轉錄組的影響,基于高通量RNA-Seq建立了12個cDNA文庫。去除低質量讀段、不確定堿基、過短序列以及可能污染的序列后,總共獲得了高質量序列數20 600 209~29 151 214,高質量堿基數 6 159 570 306~8 818 959 652,其中Q30>93.58%,鳥嘌呤和胞嘧啶(GC)所占的比例為47.58%~48.71%。此外,注釋比例表明映射到雞爪槭基因組(GCA_001876935.1,Acerpalmatum2017)的讀數比例皆較高,為81.54%~83.81%(表2)。上述結果表明該RNA-Seq測序的12組cDNA文庫可用作進一步分析的測序數據。
2.3 鹽脅迫及菌根共生下彩葉樹原始轉錄本的功能注釋及分類
根據RNA-Seq測序結果,基于與參比基因組(GCA_001876935.1,Acerpalmatum2017)進行對比總共鑒定出6 019個原始轉錄本,且都均勻分布在染色體上。功能注釋見圖2,在上述數據庫中總共注釋得到4 672個新基因。可注釋到COG、GO、KEGG、KOGPfam、Swiss-Prot、egg-NOG及Nr的新基因數分別為626、1751、1181、1920、1 660、2 190、3 057 及4 644個,分別占總的新基因數的13.40%、37.48%、25.28%、41.10%、35.53%、46.88%、65.43%及99.40%。
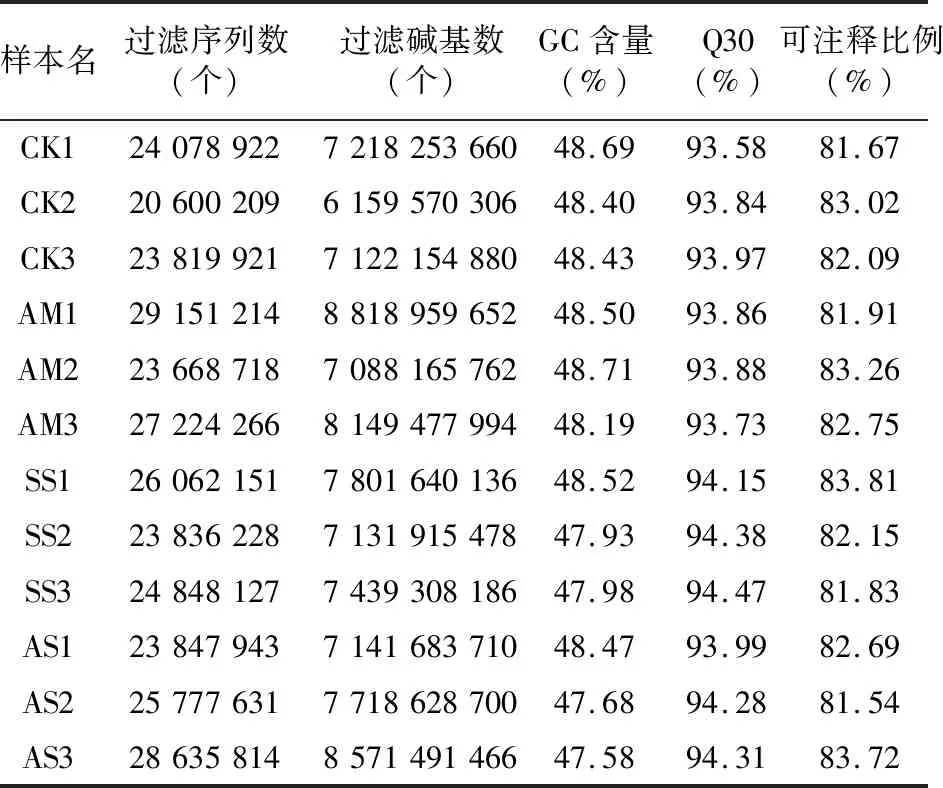
表2 鹽脅迫下菌根共生下彩葉樹轉錄本的原始數據分析

2.4 鹽脅迫及菌根共生下彩葉樹轉錄本的差異基因(DEGs)分析
由圖3-A可知,當進行鹽脅迫處理且無論接種叢枝菌根與否的對比組中(AM/SS、CK/SS),差異基因的下調數量整體上是上調數量的2倍。對于接種AM真菌處理中,在沒有和有鹽脅迫對比下(CK/AM、SS/AS),上調的DEGs數量則幾乎是下調的DEGs數量的2 倍。這些結果表明,雞爪槭中更多的DEGs響應于鹽脅迫而發(fā)生下調,而更多的DEGs響應于接種AM真菌而上調。有趣的是,本研究發(fā)現41個基因在比較CK/SS處理時下調,但在比較SS/AS處理時卻上調。此外,在比較SS/AS處理時,有286個基因上調,但在比較CK/SS處理時,這些基因的表達水平沒有差異。因此,可以推斷具有這些表達特征的DEGs可能是鹽脅迫條件下響應菌根共生的關鍵基因。
為了進一步探索來自4個不同比較處理的DEGs之間的關系,構建了不同比較處理間DEGs的Venn圖。由圖3-B可知,僅在接種與不接種AM真菌條件下(CK/SS、AM/AS),共有798個DEGs,其中未接種的植物特有1 630個DEGs。這些結果表明,在接種和未接種AM真菌的植物對鹽脅迫的分子反應存在明顯差異。此外,為了識別鹽脅迫下AM真菌特異性調控的DEGs,SS/AS與CK/AM僅共有1個共同DEGs,這表明大多數DEGs在SS和AS處理的比較可能與AM真菌調節(jié)的鹽脅迫有關。因此SS/AS中的這些DEGs(455個)可用于后續(xù)基因功能表征以分析由AM真菌誘導耐鹽性的可能分子機制。

2.5 鹽脅迫及菌根共生下彩葉樹差異基因(DEGs)的功能注釋
2.5.1 鹽脅迫及菌根共生下彩葉樹DEGs的GO功能注釋與分類 為了解釋AM真菌在鹽度脅迫中的改善機制,研究了在SS/AS比較中的455個DEGs的特征。對于GO分析,455個DEGs中有247個被分配到至少1個GO分類中,其主要涉及生物過程、分子功能和細胞成分(圖4);DEGs顯著富集(KS值≤0.05)在32個過程中產生富集(表3)。這些富集的GO term在鹽度脅迫的適應性反應中的生物過程高度相關,包括蛋白質生物合成相關過程(GO:0006383、GO:0006452、GO:0006449、GO:0045901、GO:0045905、GO:0006414和GO:0006415)、ATP生物合成過程(GO:0006754)、蛋白質谷胱甘肽化(GO:0010731)、細胞分化調控(GO:0045595)、氮同化相關過程(GO:0010167、GO:0009309、GO:0009084)、呼吸電子傳遞鏈(GO:0022904)以及類胡蘿卜素代謝過程(GO:0043288)。
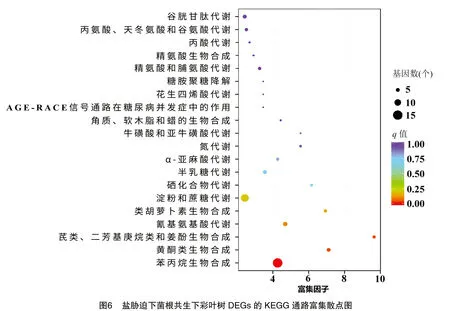
2.5.2 鹽脅迫及菌根共生下彩葉樹DEGs的KEGG功能注釋與富集分析 基于京都基因與基因組百科全書(KEGG)分析中,455個DEGs中有98個被分配了KEGG ID并被分類為58個途徑(圖5)。在圖6中突出顯示了前 20 條富集途徑(基于q值),其中,DEGs顯著(q值≤0.05)富集在與次級代謝物生物合成相關的3條途徑中,包括苯丙烷、黃酮類、二苯乙烯類、二芳基庚烷類和姜酚的通路代謝(圖5、圖6),這些通路可能與耐鹽性有關。
2.6 鹽脅迫及菌根共生下彩葉樹DEGs的qRT-PCR驗證
為了驗證RNA-Seq數據的可靠性,在SS/AS的455個DEGs中隨機挑選了6個DEGs進行qRT-PCR分析。通過qRT-PCR分析發(fā)現,Gene23660、Gene26140、New_Gene4379、Gene20176、Gene20188、New_Gene23129的log2FC值分別為9.66、3.26、3.39、2.71、4.64、2.54(圖7),這6個基因的表達趨勢與轉錄本的RNA-Seq分析結果高度一致。這些結果表明DEGs分析是準確可靠的。

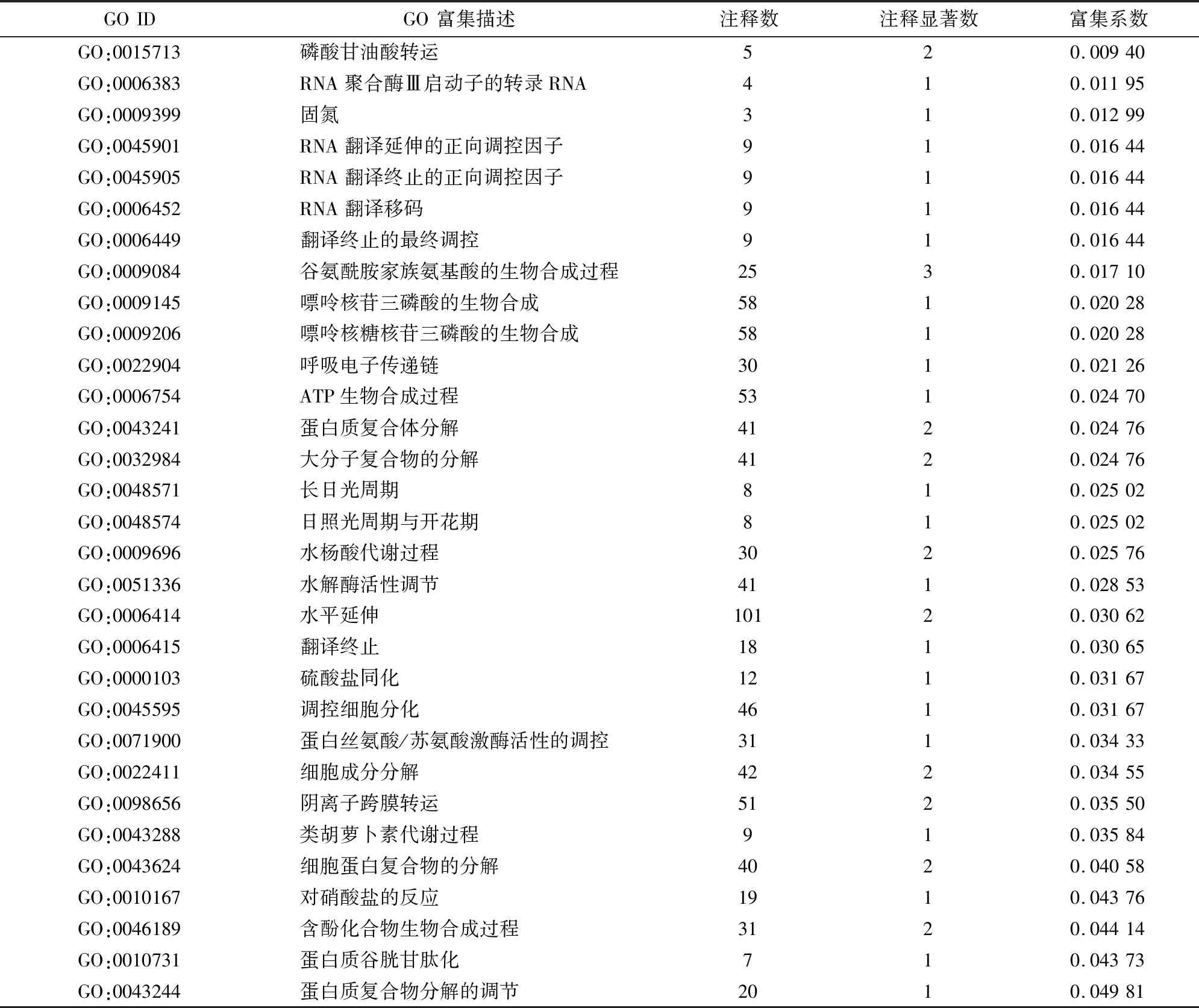
表3 鹽脅迫下菌根共生下彩葉樹DEGs的GO分類

3 討論與結論
叢枝菌根真菌在土壤中廣泛存在,是重要的功能性微生物組成部分[7]。關于AM真菌改善鹽脅迫的研究已經在煙草、黃瓜、玉米以及小麥等多種植物中得到證實[17-19]。為了更好地了解AM真菌對植物耐鹽性的潛在分子機制,在鹽脅迫下給彩葉樹雞爪槭接種AM真菌,并進行比較轉錄組分析,建立了12個測序文庫,確定了6 019個潛在基因,注釋得到4 672個新基因。此外,在鹽脅迫條件下由AM真菌誘導的455個差異基因顯著富集在幾個GO和KEGG通路中,這些通路涉及耐鹽機制,包括植物細胞內部環(huán)境的改善、氮代謝相關過程以及可能的光保護機制。這些途徑進一步證實AM真菌可調控離子滲透和氧化應激的影響,并改善鹽度脅迫下的氮代謝和光保護機制。
3.1 鹽脅迫下菌根共生涉及的細胞改善作用
首先,在鹽脅迫條件下,抗氧化酶的分泌與非酶化合物的合成在清除過量ROS以維持氧化平衡和減少非生物脅迫對細胞中的影響方面起著重要作用,如過氧化物酶(POD)、谷胱甘肽過氧化物酶(GPX)、單脫氫抗壞血酸還原酶(MDHAR)、腺苷高半胱氨酸酶(AHCY)、谷胱甘S-轉移酶(GST)、抗壞血酸(AsA)、谷胱甘肽(GSH)和硫氧還蛋白(TRX)等[18,20-22]。本研究發(fā)現,許多編碼POD、GST、MDHAR、AHCY和TRX的DEGs發(fā)生上調表達并參與了多種抗氧化系統(tǒng),包括抗壞血酸-GSH循環(huán)和過氧還蛋白/硫氧還蛋白(PrxR/Trx)途徑。

前人研究表明,AM真菌可通過調節(jié)脯氨酸代謝酶活性從而改變葉脯氨酸代謝,這對調控宿主滲透壓至關重要[23]。本研究發(fā)現,2個編碼K+通道的基因上調AKT基因的表達,這些基因參與K+在葉部中的易位[2,24]。敏感系統(tǒng)信號通路通過調節(jié)質膜和液泡中的Na+轉運在離子穩(wěn)態(tài)中發(fā)揮重要作用,這涉及胞質Ca2+信號運輸、質膜和液泡Na+/H+逆向調節(jié)以及H+-ATPase建立的H+梯度驅動過程等[25]。本研究中,參與SOS途徑的相關基因發(fā)生上調,包括編碼Ca2+結合蛋白(SOS3)和CBL基因家族相互作用蛋白激酶(SOS2)的DEGs。此外,CK/SS比較中發(fā)現編碼V型H+轉運三磷酸腺苷(ATP)酶的基因表達水平增加,編碼質膜ATP酶的基因在SS/AS中也發(fā)生上調,這對于在液泡膜和質膜上建立電化學H+梯度至關重要。這表明接種AM真菌通過在更全面、更平衡的滲透調控網絡中調整過量Na+傳輸來促進離子穩(wěn)態(tài)。此外,過量的Na+通常會導致鹽度脅迫下糖生植物中K+缺乏,因此,AKT基因的上調表達也可能在植物細胞中保持較高的K+/Na+發(fā)揮作用,從而適應鹽脅迫[26]。
3.2 鹽脅迫下菌根共生對氮代謝相關過程的調控作用
氮(N)是限制植物生長和發(fā)育的常量營養(yǎng)素,研究表明氮吸收、運輸、還原和同化以及氨基酸代謝等代謝過程都受到鹽度脅迫的影響[27]。就植物對鹽漬土的適應而言,增強N代謝可能是AM真菌改善宿主發(fā)育的重要功能之一[17]。谷氨酰胺合成酶(GS)、谷氨酸脫氫酶(GDH)和谷氨酸合成酶(GOGAT)是參與N同化的關鍵酶[28]。在本研究中,編碼GS和GDH的DEGs在鹽度脅迫下發(fā)生下調表達;接種AM真菌后,編碼GS、GDH和GOGAT以及NRT1/PTR家族中編碼蛋白質的DEGs表達發(fā)生上調。這表明彩葉樹雞爪槭的耐鹽性增強可能與接種AM真菌提高N代謝效率有關。一些研究表明,AM真菌還會導致編碼與次級代謝相關的酶的基因表達水平增加,例如酚類化合物、黃酮類化合物和木質素的代謝,這些次級代謝涉及抗氧化、防御系統(tǒng)和耐鹽調控[29]。本研究發(fā)現,455個已鑒定的DEGs在參與次級代謝產物生物合成的苯丙烷、黃酮類、二苯乙烯類、二芳基庚烷類和姜酚的等生物途徑中存在顯著富集,這與前人的研究結果[29]基本一致。這些結果表明,接種AM真菌可激活氮素相關次級代謝基因從而可能在增強耐鹽性中起重要作用。
3.3 鹽脅迫下菌根共生可能涉及的光保護機制
光合作用是受鹽脅迫影響的主要過程之一,可能會導致過多的光消耗,從而導致光抑制甚至光損傷[30]。然而,植物已經進化出多種機制來保護自己免受光損傷,包括通過平衡光能的吸收和利用以及細胞環(huán)境的修復來避免光抑制[2]。本研究中,結合轉錄組數據結果,在鹽脅迫下彩葉樹雞爪槭可能通過下調編碼光捕獲葉綠素a、葉綠素b結合蛋白(LHCⅡ)基因的表達水平從而降低光抑制,葉綠素是植物體中最豐富的光收集器[31]。因為LHCⅡ從PSⅡ遷移到PSⅠ的狀態(tài)轉變是由編碼絲氨酸/蘇氨酸蛋白激酶基因(STN7)的表達水平上調啟動的,從而平衡光系統(tǒng)之間的激發(fā)能量分布[29]。
本研究中,另一個顯著的變化是參與類胡蘿卜素代謝過程(GO:0043288)的DEGs水平發(fā)生了改變,類胡蘿卜素具有ROS清除、光保護和膜穩(wěn)定性功能,類胡蘿卜素代謝DEGs上調可有助于提高耐鹽性[32]。光反應中,多余的光能可以從葉綠體輸出并通過線粒體呼吸鏈消散[33],這涉及替代呼吸途徑能量守恒電子傳遞(ETC)途徑的重要旁路,該途徑涉及Ⅱ型煙酰胺腺嘌呤二核苷酸磷酸(NADPH)脫氫酶(NDs)和抗氰化物替代氧化酶(AOX)[27-28],從而在光保護中起著關鍵作用。本研究中,在鹽脅迫下編碼AOX的基因被下調,同時隨著接種AM真菌編碼NDB2(一種Ⅱ型NDs)的基因被上調。這些結果表明,接種AM真菌的彩葉樹雞爪槭采用多種策略來保護自己免受光損傷從而克服鹽脅迫。
綜上,本研究中,基于彩葉樹雞爪槭基因組分析總共鑒定出6 019個原始轉錄本,通過注釋到COG、GO、KEGG、KOGPfam、Swiss-Prot、egg-NOG及Nr的新基因數共得到4 672個新基因,并在鹽脅迫處理(SS)和鹽脅迫下AM真菌接種處理(AS)的比較轉錄中鑒定了455個差異基因。進一步研究表明,在455個DEGs中,一些被鑒定為耐鹽基因,因為它們參與植物細胞內部環(huán)境的改善、氮代謝相關過程和可能的光保護機制。本研究為后續(xù)研究功能分析的耐鹽候選基因提供了理論依據。
