陽春砂花絲、花柱轉(zhuǎn)錄組測序及生物信息學(xué)分析
2022-12-13 01:14:12楊博涵湯麗云李子翔何國振
江蘇農(nóng)業(yè)科學(xué) 2022年22期
楊博涵, 徐 杰,2, 湯麗云, 蘇 景, 李子翔, 何國振
[1.廣州中醫(yī)藥大學(xué)中藥學(xué)院,廣東廣州 510006; 2.廣東一方制藥有限公司/廣東省中藥配方顆粒企業(yè)重點實驗室,廣東佛山 528244;3. 華南農(nóng)業(yè)大學(xué)生命科學(xué)學(xué)院,廣東廣州 510642; 4.陽春市農(nóng)業(yè)試驗場(陽春市春砂仁試驗場),廣東陽春 529600]
陽春砂(Amomumvillosum)是姜科(Zingiberaceae)豆蔻屬(Amomum)多年生常綠草本植物[1],其干燥成熟果實為四大南藥之首——砂仁的主流品種,具有理氣安胎、溫脾止瀉、化濕開胃的功效[2]。作為傳統(tǒng)的大宗藥材,陽春砂每年的需求數(shù)量大于300萬kg,具有重要的藥用價值及經(jīng)濟(jì)價值[3]。但陽春砂在生產(chǎn)上存在著嚴(yán)重的低產(chǎn)問題,其自然結(jié)實率僅有1.1%[4],研究者對此進(jìn)行了許多研究,認(rèn)為較低的自然結(jié)實率與陽春砂小花的特殊花器結(jié)構(gòu)有著密切的聯(lián)系。何國振等將陽春砂特殊的花器結(jié)構(gòu)稱為假合蕊柱,即雌雄蕊貼合在一起,但并未完全合生,在處于開花的狀態(tài)下,唇瓣包裹著假合蕊柱,并且呈現(xiàn)出半抱合的狀態(tài),與唇瓣間的距離極近,僅有1.3~2.0 mm,嚴(yán)重阻礙了相應(yīng)蟲媒的授粉[3]。為提高產(chǎn)量,農(nóng)戶在實際種植中主要采用人工授粉的方式進(jìn)行勞作,而人工授粉成本高、勞作強(qiáng)度大、根狀莖及花序易被嚴(yán)重踩踏損傷等原因是傳統(tǒng)人工授粉方式固有的弊端,嚴(yán)重抑制了農(nóng)戶的生產(chǎn)積極性。
細(xì)胞伸長或者細(xì)胞分裂所致的植物器官運動與內(nèi)源激素及相關(guān)抑制劑作用相關(guān),而細(xì)胞結(jié)構(gòu)及生長速率的不對稱是花器官運動的基礎(chǔ)。有文獻(xiàn)研究報道,臘梅(Chimonanthuspraecox)花絲的運動來源于其兩側(cè)表面細(xì)胞生長速率的差異[5]。姜科植物馬來良姜(Alpiniamutica)花柱運動部位兩側(cè)細(xì)胞層數(shù)的差異是其花柱卷曲運動的基礎(chǔ)[6],赤霉素(GA)、茉莉酸(JA)、吲哚-3-乙酸(IAA)等激素則有著調(diào)控植物雌雄蕊發(fā)育的作用[7-9]。何卓航等研究發(fā)現(xiàn),隨著陽春砂小花的生長,其花絲、花柱遠(yuǎn)、近軸側(cè)的細(xì)胞層數(shù)出現(xiàn)了差異,這種不對稱的結(jié)構(gòu)是雌雄蕊運動的基礎(chǔ),認(rèn)為陽春砂雌雄蕊的相向運動致使其假合蕊柱的形成[10];同時對花柱的近端以及遠(yuǎn)端軸側(cè)的IAA水平進(jìn)行測定,結(jié)果表明,生長時期在保持同樣的水平不同橫切部位中,IAA水平基本上呈現(xiàn)遠(yuǎn)端軸側(cè)小于近端軸側(cè)的趨勢,故花柱運動的原因可能是來自于花柱近、遠(yuǎn)軸側(cè)IAA水平的差異,該研究從生理層面初步闡述了陽春砂假合蕊柱的形成機(jī)制。然而,若要進(jìn)一步探明陽春砂假合蕊柱形成機(jī)制,則須對陽春砂花器官運動及發(fā)育的分子機(jī)制作深入研究。
因此,本研究對陽春砂不同生長時期的花絲、花柱進(jìn)行轉(zhuǎn)錄組測序,通過數(shù)據(jù)拼接、組裝的方式建立陽春砂花絲、花柱的轉(zhuǎn)錄組數(shù)據(jù)庫,將所得的Unigene進(jìn)行功能注釋、分類以及簡單重復(fù)序列(SSR)分子標(biāo)記。同時,重點關(guān)注并篩選可能影響陽春砂雌蕊、雄蕊運動的激素合成與信號轉(zhuǎn)導(dǎo)途徑關(guān)鍵基因。以期進(jìn)一步解析陽春砂假合蕊柱產(chǎn)生的調(diào)控機(jī)制及分子機(jī)制,并為人為干預(yù)陽春砂的花器官結(jié)構(gòu),提高產(chǎn)量奠定理論基礎(chǔ)。
1 材料與方法
1.1 試驗材料
材料為大田栽培的不同生長時期陽春砂小花的花柱及花絲,陽春砂植株栽培于廣東省陽春市合水鎮(zhèn)那軟村陽春砂種植基地(22°17′N,112°01′E),于2017年5—7月在該基地開展大田試驗。參照陳紅的方法[11],按照長度的不同,將陽春砂小花的生長時期進(jìn)行了劃分(表1),并于陽春砂花期內(nèi),每天上午采摘小花,解剖并分離出不同時期小花的花絲與花柱,迅速置于液氮灌中,隨后轉(zhuǎn)移至超低溫冰箱(-80 ℃)進(jìn)行保存。
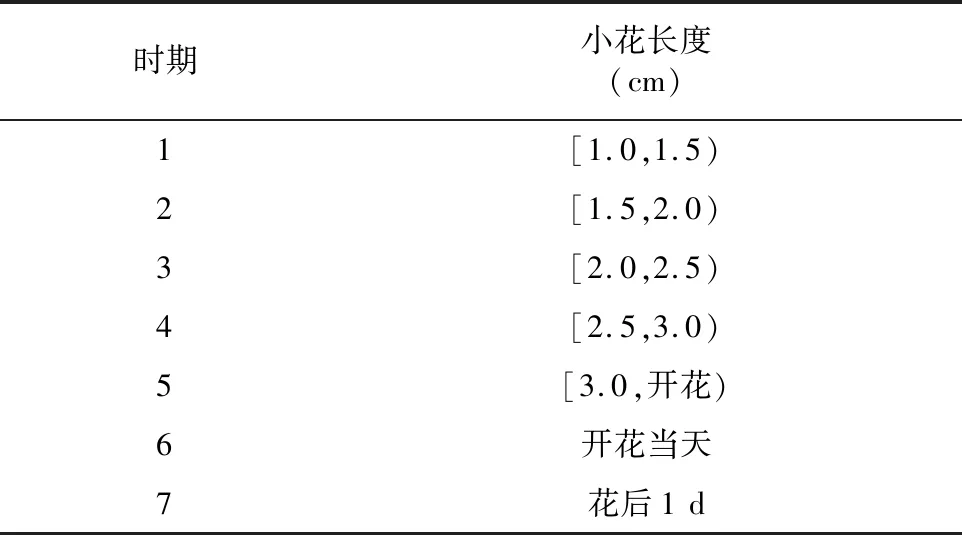
表1 陽春砂小花生長時期劃分
1.2 試驗方法
1.2.1 RNA提取 分別提取陽春砂小花在各生長時期的花柱和與花絲的總RNA,重復(fù)3次。利用NanoDrop 2000和Agilent 2100 Bioanalyzer測定RNA的濃度、純度與完整性,數(shù)據(jù)檢測合格后的RNA用于構(gòu)建轉(zhuǎn)錄組數(shù)據(jù)庫。此部分試驗與轉(zhuǎn)錄組的測序委托深圳華大基因科技服務(wù)有限公司完成。
1.2.2 cDNA文庫構(gòu)建及轉(zhuǎn)錄組序列組裝 使用Oligo(dT)磁珠富集質(zhì)檢合格且所有樣品的混合RNA。加入打斷試劑,以片段化的mRNA為模板合成1鏈、2鏈cDNA,配制反應(yīng)體系,使接頭與cDNA連接。PCR反應(yīng)及產(chǎn)物回收、擴(kuò)增。PCR產(chǎn)物變性,充分混勻,得到單鏈環(huán)形產(chǎn)物,隨后PCR產(chǎn)物變性,即得到文庫。利用Agilent 2100 Bioanalyzer和ABI StepOnePlus real-time PCR System對所得文庫進(jìn)行檢測,檢測合格后進(jìn)行轉(zhuǎn)錄組denovo測序。所得reads通過Trinity軟件進(jìn)行序列組裝,組裝序列的質(zhì)量通過BUSCO軟件進(jìn)行評估。
1.2.3 轉(zhuǎn)錄組基因功能注釋及數(shù)據(jù)挖掘 利用生物信息學(xué)方法分析獲得的陽春砂Unigene,為獲得全方位的基因功能信息,對組裝所得的Unigene在七大功能數(shù)據(jù)庫中進(jìn)行注釋,包括NR、NT、 KOG/COG、GO、KEGG、SwissPro及Interpro。使用MIcroSAtellite (MISA)工具對Unigene進(jìn)行SSR位點的挖掘。并在KEGG代謝通路中重點關(guān)注植物激素生物合成與信號轉(zhuǎn)導(dǎo)Unigene的注釋情況。
2 結(jié)果與分析
2.1 轉(zhuǎn)錄組數(shù)據(jù)組裝
使用Illumina HiSeq-2000平臺對不同時期的陽春砂花絲及花柱的cDNA進(jìn)行測序。測序的結(jié)果顯示,從轉(zhuǎn)錄組中獲得了81.40 Mb的原始讀數(shù),用過濾軟件SOAPnuke去除低質(zhì)量的reads后共得到68.48 Mb Clean reads,最終獲得10.33 Gb的堿基總數(shù)。Q20和Q30的百分比分別為97.20%和93.16%(表2)。說明轉(zhuǎn)錄組的測序質(zhì)量較高,可以滿足后續(xù)的生物信息學(xué)分析。
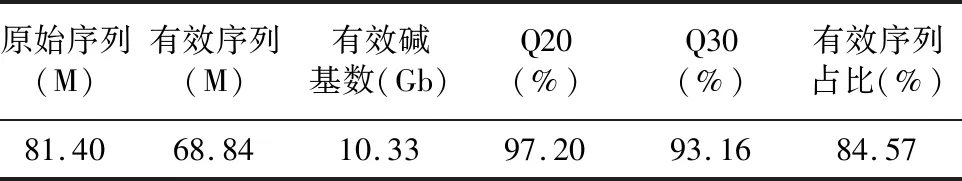
表2 測序數(shù)據(jù)質(zhì)量分析
對轉(zhuǎn)錄本中Clean reads進(jìn)行組裝,一共獲得138 590個轉(zhuǎn)錄本,包含了111 312 938個核苷酸的序列信息,這些片段長度的平均值為803 bp,N50為1 414 bp,GC含量為44.28%。對轉(zhuǎn)錄本進(jìn)一步聚類并去除冗余的序列之后得到94 584條Unigene,總長度為 92 501 015 bp。Unigene的N50、N70和GC含量分別為 1 582bp、1 002 bp和44.35%,均大于其對應(yīng)平均長度,說明組裝的結(jié)果良好(表3)。

表3 轉(zhuǎn)錄本和單基因簇統(tǒng)計分析
2.2 陽春砂轉(zhuǎn)錄組基因總體注釋情況
將所獲得的結(jié)果在七大功能數(shù)據(jù)庫進(jìn)行注釋,結(jié)果見表4。結(jié)果顯示,陽春砂共注釋到94 584條Unigene。其中,共有58 669條Unigene被NR數(shù)據(jù)庫注釋,占比最多,達(dá)到總Unigene的62.03%;NT數(shù)據(jù)庫有40 205條,占42.51%;SwissProt有40 318條,占總Unigene的42.63%;KEGG數(shù)據(jù)庫與KOG數(shù)據(jù)庫分別有44 000條和45 892條Unigene,各占46.52%與48.52%;Interpro有45 793條,占48.42%。GO數(shù)據(jù)庫注釋到的基因最少,僅有 12 050 條,占總數(shù)的12.74%;所得比對結(jié)果顯示,在七大數(shù)據(jù)庫中均能成功注釋的Unigene共有 5 691 條,占總Unigene條數(shù)的6.02%。
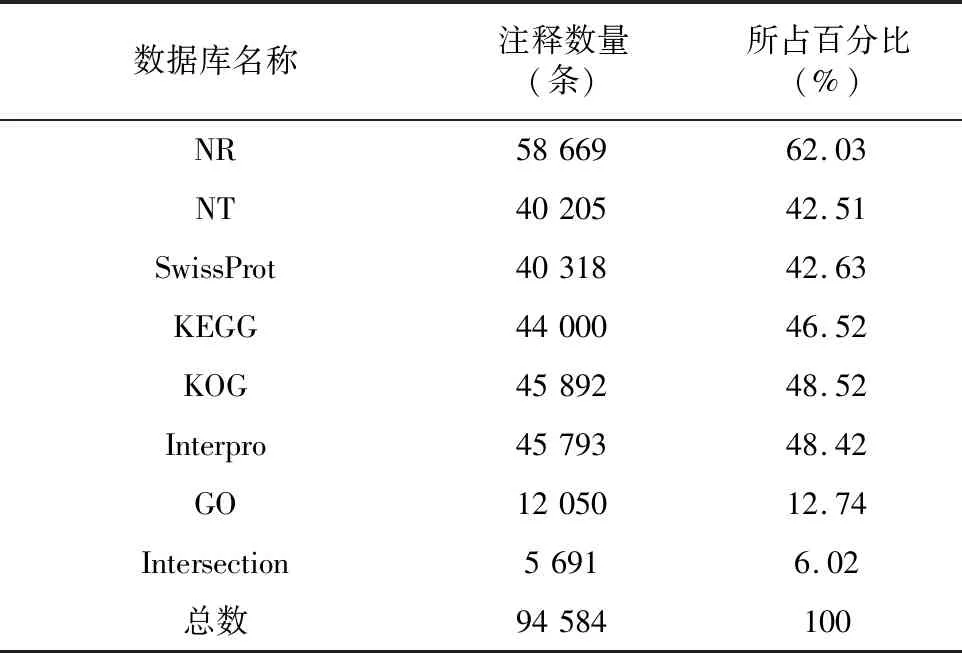
表4 轉(zhuǎn)錄組基因注釋情況統(tǒng)計
2.2.1 NR數(shù)據(jù)庫功能注釋 NR數(shù)據(jù)庫的注釋結(jié)果(圖1)顯示,匹配最多的物種為小果野芭蕉(Musaacuminatasubsp.Malaccensis),該物種注釋到的基因數(shù)量最多,共有45 257條,占比高達(dá)77.14%,證明陽春砂與該物種的同源性較高;其他物種依次為油棕(Elaeisguineensis)與海藻(Phoenixdactylifera),分別有3 338、2 435條Unigene被注釋,占比分別為5.69%、4.15%,陽春砂與這2種植物的同源性相對較低。而剩余的13.02%則分布于其他物種中。

2.2.2 KOG 數(shù)據(jù)庫功能注釋 在KOG數(shù)據(jù)庫中,共有45 892條Unigene被注釋到(圖2),共分為25個大類功能區(qū),包括一般功能預(yù)測、信號轉(zhuǎn)導(dǎo)機(jī)制、轉(zhuǎn)錄等功能。在為數(shù)眾多且不同的功能分類中,注釋到的基因的數(shù)量差異較為顯著,一般功能預(yù)測類基因數(shù)量最多,共有11 177條Unigene,占比為24.36%,其次是信號轉(zhuǎn)導(dǎo)機(jī)制,有8 271條Unigene被注釋,占比為18.02%;除此之外,負(fù)責(zé)轉(zhuǎn)錄功能的Unigene有5 470個,占比為11.92%;有1 172條Unigene注釋到負(fù)責(zé)次生代謝產(chǎn)物生物合成、運輸和代謝功能區(qū)中,占比為2.55%。
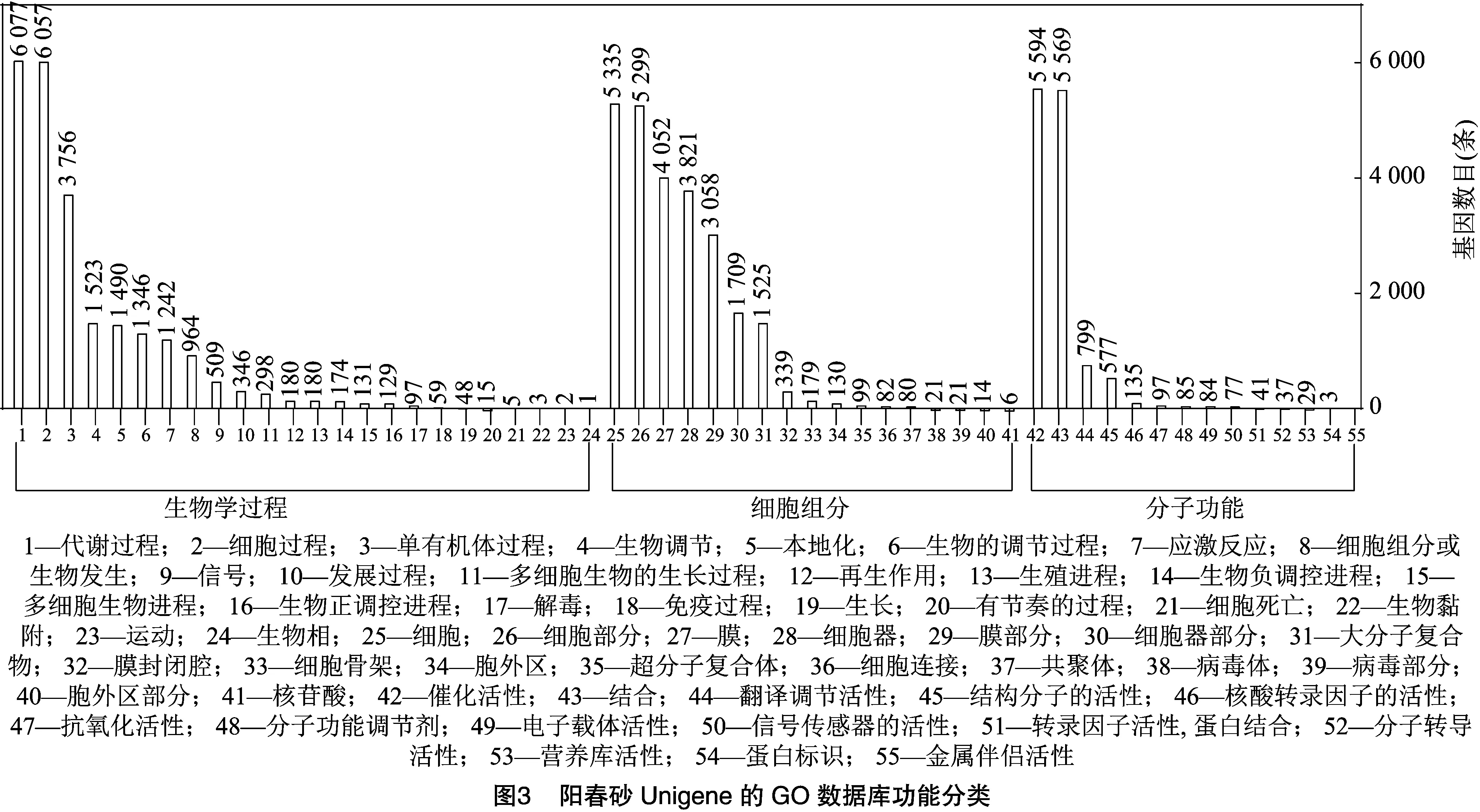
2.2.3 GO數(shù)據(jù)庫功能注釋 GO數(shù)據(jù)庫中的功能分類注釋結(jié)果見圖3,結(jié)果表明,陽春砂花絲及花柱中共有12 050條Unigene注釋到不同的功能節(jié)點上,共涉及到生物學(xué)過程、細(xì)胞組分和分子功能3個大類,55個亞類。歸入到細(xì)胞組分的17個亞類中,以細(xì)胞、細(xì)胞部分、膜和細(xì)胞器功能的Unigene數(shù)量最多,分別有5 335、5 229、4 052、3 821條。在分子功能的14個亞類中,催化活性和結(jié)合注釋數(shù)量最多,分別有5 594、5 569條。而涉及生物學(xué)過程的24個亞類中,以代謝過程(6 077條)和細(xì)胞進(jìn)程 (6 057 條)為主。結(jié)果表明,同一個Unigene可以注釋到多個功能結(jié)點上, 因此同一個功能分支的總注釋數(shù)大于注釋到該功能分支的總Unigene數(shù)。

2.2.4 KEGG數(shù)據(jù)庫功能注釋 對轉(zhuǎn)錄組測序獲得的Unigene進(jìn)行KEGG代謝通路富集分析后發(fā)現(xiàn),共有44 000條 Unigene得到注釋,涉及到137個通路。對富集顯著的前20條通路進(jìn)行分析(表5),其中,共有8 660條Unigene被代謝途徑通路所注釋,占比最多,達(dá)到注釋總數(shù)的19.68%。而次生代謝產(chǎn)物的生物合成通路共有4 191條Unigene被注釋到,占注釋總數(shù)的9.53%,另外,本研究主要關(guān)注的植物激素轉(zhuǎn)導(dǎo)通路一共有1 656條Unigene被注釋到,占比為3.76%。
在所有的代謝通路中,共有23個與次生代謝相關(guān)的代謝通路,對其所有的Pathway進(jìn)行分析后(表6)發(fā)現(xiàn),一共有4 191個Unigene被次生代謝產(chǎn)物的生物合成途徑所注釋;而在本研究主要關(guān)注的植物激素合成途徑中,62個Unigene被油菜素內(nèi)酯生物合成途徑所注釋,另有59個Unigene被玉米素生物合成途徑所注釋。
2.2.5 SSR特征分析 轉(zhuǎn)錄組測序所得Unigene的SSR的檢測最終結(jié)果顯示,共有18 895個SSR被檢測到(圖4)。總共有14種多堿基重復(fù)SSR,其中, 三核苷酸重復(fù)的SSR 數(shù)目最多,共有6 313個,占SSR位點總數(shù)的33.41%;三核苷酸重復(fù)中AGG/CCT重復(fù)基元最多,共有1 434個;單核苷酸和二核苷酸重復(fù)分別有6 238個(33.01%)和5 128個(27.14%),二核苷酸中重復(fù)基元最多的是AG/CT,共有2 909個;六核苷酸重復(fù)數(shù)目共有546個(2.89%),四核苷酸重復(fù)330個(1.75%)和五核苷酸重復(fù)340個(1.80%)。
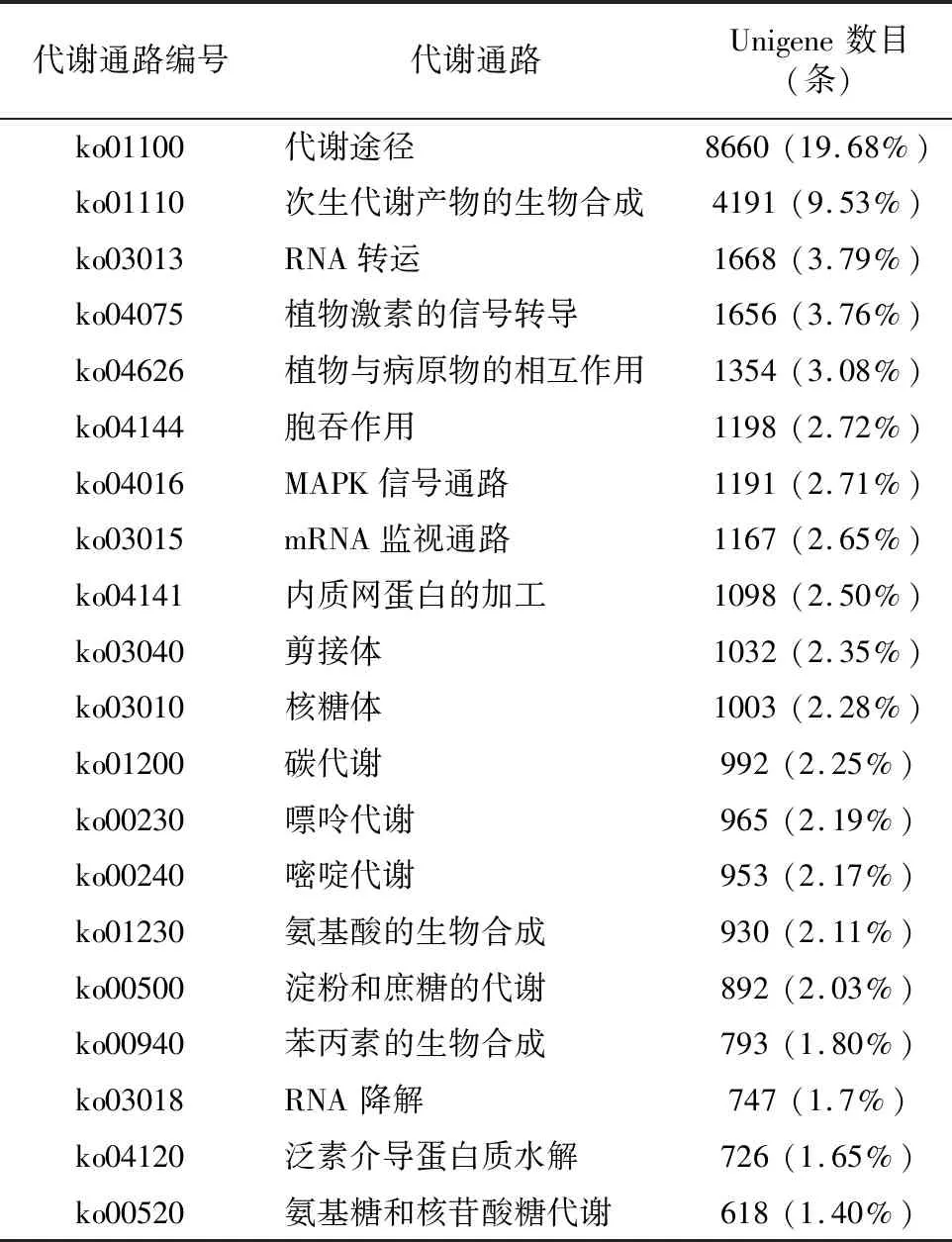
表5 次生代謝相關(guān)代謝通路的注釋情況

表6 次生代謝相關(guān)代謝通路的注釋情況

2.3 重點關(guān)注通路的Unigene 注釋情況
2.3.1 植物激素信號轉(zhuǎn)導(dǎo)途徑中Unigene注釋情況 在KEGG代謝通路中,共有1 760條Unigene注釋到植物激素的信號轉(zhuǎn)導(dǎo)途徑中(表7)。赤霉素的信號轉(zhuǎn)導(dǎo)途徑中注釋到的5個基因,對應(yīng)的Unigene最多,共有438個;其中,植物光敏色素互作因子(PIF)對應(yīng)的Unigene最多,達(dá)到318個;而水楊酸信號轉(zhuǎn)導(dǎo)途徑注釋到3個基因,包括非表達(dá)病程相關(guān)蛋白(NPR1)、TGA轉(zhuǎn)錄因子及病程相關(guān)蛋白1(PR1)基因,但對應(yīng)的Unigene最少,僅有74個;茉莉酸的信號轉(zhuǎn)導(dǎo)途徑中注釋到的茉莉酸ZIM結(jié)構(gòu)域蛋白(JAZ)、茉莉酸氨基酸合成酶(JAR1)、MYC2轉(zhuǎn)錄因子和茉莉酸受體蛋白(COI1)一共對應(yīng)了176條Unigene;乙烯和油菜素內(nèi)酯信號轉(zhuǎn)導(dǎo)途徑注釋到的基因最多,均為8個,分別注釋到139條及191條Unigene;其他植物激素中,生長素、細(xì)胞分裂素以及脫落酸的信號轉(zhuǎn)導(dǎo)途徑分別注釋到329、245、168條Unigene;統(tǒng)計結(jié)果中,各激素信號轉(zhuǎn)導(dǎo)途徑關(guān)鍵基因均有注釋,如細(xì)胞分裂素中的磷酸轉(zhuǎn)移蛋白(AHP)、生長素中的生長素載體(AUX1)、生長素/吲哚乙酸蛋白(Aux/IAA)轉(zhuǎn)錄家族以及脫落酸信號轉(zhuǎn)導(dǎo)途徑中的ABF轉(zhuǎn)錄因子等。綜上,研究較為深入的植物激素基本均在KEGG通路中有所注釋,且其信號轉(zhuǎn)導(dǎo)途徑的關(guān)鍵基因都有一定數(shù)量的Unigene被注釋到。

表7 轉(zhuǎn)錄組中植物激素信號轉(zhuǎn)導(dǎo)途徑Unigene注釋情況
2.3.2 植物激素合成途徑中Unigene注釋情況 轉(zhuǎn)錄組中激素合成途徑酶基因Unigene注釋情況(表8)顯示,赤霉素注釋的酶基因最多,包括內(nèi)根-貝殼杉烯合成酶(KS)、古巴焦磷酸合成酶(CPS)、內(nèi)根-貝殼杉烯氧化酶(KO)及GA3氧化酶(GA3ox)等8個合成通路中的關(guān)鍵酶基因,一共對應(yīng)67條Unigene;油菜素內(nèi)酯與水楊酸合成途徑中均有1個酶基因被注釋到,為異分支酸合酶(ICS)與油菜素內(nèi)酯-6-氧化酶2(CYP85A2),分別對應(yīng)9條和24條Unigene;茉莉酸合成途徑2個關(guān)鍵酶基因丙二烯氧化物合成酶(AOS)以及丙二烯氧化物環(huán)化酶(AOC)分別注釋到6條和7條Unigene,生長素一共注釋到42條Unigene,色氨酸轉(zhuǎn)氨酶(TAA1)及吲哚-3-丙酮酸單加氧酶(YUC)分別注釋到了其中22條和20條;乙烯合成途徑中的乙烯合成前體合成酶(ACS)與乙烯合成前體氧化酶(ACO)分別對應(yīng)9和14條Unigene;脫落酸及玉米素的生物合成途徑中均注釋到3個關(guān)鍵酶基因,分別有29條和45條Unigene。

表8 轉(zhuǎn)錄組中植物激素生物合成途徑Unigene 注釋情況
3 討論與結(jié)論
近年來,轉(zhuǎn)錄組測序技術(shù)發(fā)展迅速,同時伴隨著多個生物信息學(xué)分析平臺的加入,大量研究即便在沒有基因組數(shù)據(jù)支撐的前提下,通過轉(zhuǎn)錄組測序所得的結(jié)果也可在挖掘植物未知基因、明確相關(guān)生理功能的代謝途徑及基因調(diào)控機(jī)制等方面提供海量相關(guān)信息。無需參考基因組的數(shù)據(jù)便可對目標(biāo)物種進(jìn)行分子生物學(xué)方面的研究,已成為中草藥材基因信息挖掘的重要研究手段[12]。利用該技術(shù),前人已從多種植物中挖掘出有關(guān)花器官發(fā)育的基因,并分析了相關(guān)基因的調(diào)控機(jī)制。李梅等利用雌蕊缺失茶樹(Camelliasinensis)花的花、花蕾、花芽作為材料,基于轉(zhuǎn)錄組測序技術(shù),共發(fā)掘出了34個花器官發(fā)育差異表達(dá)的相關(guān)基因,包括KNOX家族基因、WUS類基因、花器官發(fā)育 ABCDE 模型的相關(guān)基因等,證明茶樹花即使是在雌蕊缺失的情況下,有關(guān)雌蕊發(fā)育的基因也是同樣存在[13]。而在長瓣兜蘭(Paphiopedilumdianthum)花蕾和花朵的轉(zhuǎn)錄組測序中發(fā)現(xiàn),相較于花蕾期,花朵時期中調(diào)控花朵發(fā)育相關(guān)的基因,如AG、C2H2-ZEP等基因的表達(dá)量明顯下調(diào),大量基因表達(dá)下調(diào)的原因可能是在花朵時期,花的各部分器官已完成分化,調(diào)控器官分化基因的相關(guān)任務(wù)已完成[14]所致。位明明等通過該技術(shù)初步明確了多個參與編碼橡膠樹(Heveabrasiliensis)花器官發(fā)育的基因家族,如Mads-box、MYB及AP2等基因家族[15]。
本研究通過RNA-Seq技術(shù)構(gòu)建了陽春砂不同時期花絲以及花柱總樣品的轉(zhuǎn)錄組數(shù)據(jù)庫,共獲得94 584條Unigene,Unigene的平均長度為842 bp,N50為1 582 bp,GC含量為44.35%。其中,共有 62 174 條Unigene被注釋到GO、KEGG、NR和KOG等七大數(shù)據(jù)庫,占總Unigene的 65.73%。注釋率較高,測序所得結(jié)果與拼接質(zhì)量較好。這些注釋的Unigene為陽春砂的代謝途徑、基因功能分類、花器官發(fā)育及雌雄蕊運動分析等方面提供參考依據(jù)。此外,一共有32 410條Unigene未被注釋到,占總Unigene的34.27%,推測這些Unigene可能為陽春砂中的非編碼 RNA 序列或現(xiàn)有基因數(shù)據(jù)庫尚未完善所致。在NR數(shù)據(jù)庫中共注釋到58 669條Unigene,其中77.14%的Unigene被注釋到姜科植物小果野芭蕉中,兩者比對所得同源基因數(shù)目較多的原因可能是親緣關(guān)系較近。轉(zhuǎn)錄組所得Unigene在GO功能分類中,注釋到生物學(xué)過程、細(xì)胞組分以及分子功能三大類,分別有 24、17、14個亞類,主要集中在代謝過程、催化活性、細(xì)胞部分和細(xì)胞過程等功能。而在KOG數(shù)據(jù)庫的分析結(jié)果中,共有 45 892 條Unigeine得到了注釋,在25個功能大類中,一般功能預(yù)測的Unigene最多,共有11 117條,信號轉(zhuǎn)導(dǎo)機(jī)制、轉(zhuǎn)錄以及翻譯后的修飾、蛋白質(zhì)轉(zhuǎn)換與代謝功能次之,核結(jié)構(gòu)與細(xì)胞運動最少,分別僅有346條和75條Unigene。通過KEGG數(shù)據(jù)庫分析發(fā)現(xiàn),被注釋到44 000條Unigene參與了23類共計137個KEGG代謝通路,這些通路主要集中在代謝途徑、次生代謝產(chǎn)物的生物合成、RNA轉(zhuǎn)運以及植物激素信號轉(zhuǎn)導(dǎo)等,可以通過所注釋的通路全面了解陽春砂花絲及花柱的代謝途徑信息。同時,在陽春砂花絲、花柱轉(zhuǎn)錄組中一共挖掘到18 895個SSR位點,在6種不同的核苷酸重復(fù)類型中均有分布,證明本研究轉(zhuǎn)錄組中位點的類型豐富,分布密度較大,具有良好的多態(tài)性潛能;在二核苷酸和三核苷酸中,重復(fù)最多的基序是分別是AG/CT和AGG/CTT,該結(jié)果與王煥的研究結(jié)果[16]一致。
植物激素是植物通過自身代謝所產(chǎn)生的一些有機(jī)信號分子,可在低濃度下產(chǎn)生明顯的生理效應(yīng),并在合成部位發(fā)揮功能[17-18]。而花器官的運動受到激素調(diào)節(jié),Luo等研究發(fā)現(xiàn),外源施加IAA后,會明顯影響姜科植物花柱的彎曲程度[19]。有些植物激素也會調(diào)控姜科植物藍(lán)豬耳二長雄蕊的翻轉(zhuǎn)與伸長[20]。此外,其他植物激素同樣也在雌雄蕊發(fā)育中起著重要的調(diào)控作用。JA可調(diào)控花藥的開裂,JA生物合成通路的相關(guān)基因DAD1與OPR3在生長素感知缺陷的擬南芥突變體中的表達(dá)量提高,可提前使得花藥產(chǎn)生開裂,說明此過程是一種負(fù)向調(diào)節(jié),即生長素可以通過介導(dǎo)JA的生物合成來調(diào)節(jié)花藥的發(fā)育[21-22]。在模式植物擬南芥的其他研究中,茉莉酸與赤霉素信號轉(zhuǎn)導(dǎo)相關(guān)基因也在擬南芥雄蕊的發(fā)育過程中起到了重要的調(diào)控作用[8,23-24]。在本研究重點關(guān)注的植物激素合成與信號轉(zhuǎn)導(dǎo)途徑中一共有2012條Unigene被注釋,囊括了赤霉素、生長素、茉莉酸及脫落酸等9種重要的植物激素,為進(jìn)一步從分子層面闡述陽春砂花器官的發(fā)育及運動提供了大量候選基因。
筆者所在課題組于2012年提出通過生理手段改變陽春砂的花器結(jié)構(gòu),以適應(yīng)更多種類的昆蟲傳粉而提高產(chǎn)量的研究思路[25]。近10年來,已經(jīng)在陽春砂的花芽分化規(guī)律[26]、花器結(jié)構(gòu)特征[3]、假合蕊柱的形成過程[10]、某些生殖生物學(xué)特性[27]、落果規(guī)律[28]等多方面進(jìn)行了研究,明晰了陽春砂生產(chǎn)過程中由于特殊花器官結(jié)構(gòu)導(dǎo)致傳粉昆蟲種類受限和低產(chǎn)的原因。因此,下一步的研究方向需對本研究中所獲有關(guān)陽春砂花器官的發(fā)育及運動的關(guān)鍵候選基因進(jìn)行更深層次的功能分析與驗證,并對陽春砂假合蕊柱形成與相關(guān)基因表達(dá)模式的關(guān)系進(jìn)行分析與探討,不僅可為深入開展陽春砂花器官發(fā)育與運動相關(guān)基因的克隆及相互作用的模式分析提供數(shù)據(jù)基礎(chǔ),也為進(jìn)一步深入揭示陽春砂假合蕊柱的分子調(diào)控機(jī)制提供了理論依據(jù),以期形成人為干預(yù)的效果進(jìn)而改變陽春砂花器官的形態(tài),拓寬陽春砂唇瓣與假合蕊柱之間的距離,為陽春砂的授粉蟲媒打造一個相對有利的授粉環(huán)境,從而提高自然結(jié)實率,達(dá)到增加農(nóng)民收入的目的。
猜你喜歡
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
財經(jīng)(2017年2期)2017-03-10 14:35:35
爆笑show(2016年7期)2017-02-09 09:36:13
財經(jīng)(2016年15期)2016-06-03 07:38:02
財經(jīng)(2016年3期)2016-03-07 07:44:46
財經(jīng)(2016年6期)2016-02-24 07:41:51
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
