
荔枝P型ATP酶基因家族鑒定及其在采后貯藏過程中響應(yīng)拮抗菌N-1的表達(dá)模式分析
2022-12-16 02:57:52邵遠(yuǎn)志
熱帶作物學(xué)報(bào) 2022年11期
關(guān)鍵詞:分析
王 鑫,邵遠(yuǎn)志,湯 月,李 雯*
荔枝P型ATP酶基因家族鑒定及其在采后貯藏過程中響應(yīng)拮抗菌N-1的表達(dá)模式分析
王 鑫1,邵遠(yuǎn)志2,湯 月2,李 雯1*
1. 海南大學(xué)園藝學(xué)院海南省熱帶園藝作物品質(zhì)調(diào)控重點(diǎn)實(shí)驗(yàn)室,海南海口 570228;2. 海南大學(xué)生命科學(xué)學(xué)院,海南海口 570228
P型ATP酶是植物體內(nèi)一種重要的膜轉(zhuǎn)運(yùn)蛋白,在果實(shí)能量代謝和酸代謝中發(fā)揮重要作用,并廣泛參與植物離子運(yùn)輸、抗逆、細(xì)胞信號轉(zhuǎn)導(dǎo)等各項(xiàng)生命活動,然而在荔枝或其他無患子科植物中尚未對P型ATP酶基因家族進(jìn)行全面分析,本研究基于‘妃子笑’荔枝轉(zhuǎn)錄組數(shù)據(jù),通過對這些基因進(jìn)行生理生化分析、生物信息學(xué)分析和表達(dá)分析,共鑒定了28個(gè)P型ATP酶基因家族成員基因,探討P型ATP酶在荔枝采后貯藏過程中的潛在功能。結(jié)構(gòu)和系統(tǒng)發(fā)育分析表明該基因可以分為5個(gè)亞家族,同一亞族的基因在基因結(jié)構(gòu)和motif基序方面具有很大的相似性,其蛋白二級結(jié)構(gòu)主要以α-螺旋為主,亞細(xì)胞定位預(yù)測分析發(fā)現(xiàn)LcPAs多定位于線粒體(21/28)。通過轉(zhuǎn)錄組分析,在采后貯藏期間,荔枝P型ATP酶家族基因中上調(diào)表達(dá)的基因明顯高于下調(diào)表達(dá)基因,暗示其可能在荔枝采后品質(zhì)劣變過程中發(fā)揮著積極的抵御作用。使用拮抗菌N-1處理荔枝可以有效抑制其果實(shí)采后褐變和病害的發(fā)生,結(jié)合RT-qPCR結(jié)果,發(fā)現(xiàn)拮抗菌N-1處理能誘導(dǎo)和在整個(gè)貯藏過程中的上調(diào)表達(dá),并維持了較高的ATP酶活性,進(jìn)一步說明P型ATP酶家族基因在抑制荔枝病變過程中發(fā)揮著重要作用。本研究為了解荔枝P型ATP酶基因家族基因的進(jìn)化和功能分析提供了參考,為深入了解拮抗菌N-1保鮮機(jī)理的分子調(diào)控機(jī)理提供新的證據(jù)。
荔枝;P型ATP酶基因家族;轉(zhuǎn)錄組;拮抗菌N-1
P型ATP酶是在植物整個(gè)生命過程中都發(fā)揮重要作用的膜轉(zhuǎn)運(yùn)蛋白,參與廣泛的細(xì)胞基本代謝過程。迄今為止,在闡明P型ATP酶的結(jié)構(gòu)、功能和調(diào)控特性方面都取得了極大的進(jìn)展。植物P型ATP酶在結(jié)構(gòu)上有一個(gè)亞基,8~12個(gè)跨膜(TM)片段,N端和C端暴露于細(xì)胞質(zhì),以及一個(gè)大型的中央細(xì)胞質(zhì)的結(jié)構(gòu)域,其上包含磷酸化和ATP結(jié)合位點(diǎn),P型ATP酶最重要的一個(gè)特征是形成一個(gè)磷酸化的中間產(chǎn)物,因此被命名為P型,P型泵的復(fù)雜性也反映了P型ATP酶的多樣性和這些轉(zhuǎn)運(yùn)體具有離子特異性[1]。目前已經(jīng)從擬南芥全基因組序列中鑒定出45個(gè)編碼P型ATP酶的基因[2],從向日葵基因組中鑒定出13個(gè)P型H+-ATP酶基因家族成員[3],從陸地棉中鑒定出98個(gè)P型ATP酶家族基因[4],從大豆基因組中鑒定了105個(gè)P型ATP酶基因[5],從水稻中鑒定出43個(gè)P型ATP酶基因家族成員等,通過蛋白序列和系統(tǒng)發(fā)育比較,一個(gè)共同的被子植物祖先擁有至少23個(gè)P型ATP酶基因家族成員的集合[6],說明它普遍存在于各種生物中,并廣泛參與植物離子運(yùn)輸、抗逆性、細(xì)胞信號轉(zhuǎn)導(dǎo)等各項(xiàng)生命活動。近年來大量研究表明P型ATP酶和植物的酸代謝密切相關(guān),從矮牽牛的遺傳分析中發(fā)現(xiàn)2種P型ATP酶PH1和PH5,這2種酶通過改變花瓣細(xì)胞液泡酸度從而改變花的顏色[7],通過進(jìn)一步研究發(fā)現(xiàn),這2個(gè)基因在柑橘類水果的酸調(diào)控中發(fā)揮重要作用[8]。此外,P型ATP酶還參與細(xì)胞內(nèi)PH穩(wěn)態(tài)的調(diào)控,為植物多胺(plant polyamines)PA前體向液泡運(yùn)輸提供動力[9],并在大豆鹽、干旱、寒冷、磷酸鹽饑餓等脅迫下被顯著誘導(dǎo)或抑制[5],在水稻和擬南芥的非生物脅迫中發(fā)揮重要作用[10]。
生防治作為一種安全健康的應(yīng)對果蔬保鮮問題的新途徑,是最有希望替代傳統(tǒng)化學(xué)保鮮劑的一種保鮮方法,其中拮抗菌生物防治具有安全、綠色和環(huán)保的優(yōu)點(diǎn),成為荔枝采后保鮮技術(shù)研究的熱點(diǎn)[11-12]。然而拮抗菌作為保鮮劑的作用機(jī)理十分復(fù)雜,目前可以確定的是果實(shí)能量代謝在其中起到了十分重要的作用。提高果實(shí)的能荷水平可以減緩桃[13]、梨[14]和枇杷[15]等果實(shí)病害的發(fā)生,陳夢茵[16]研究發(fā)現(xiàn)外源ATP處理能提高Ca2+-ATPase和H+-ATPase酶活性,維持果實(shí)較高的ATP含量,保護(hù)細(xì)胞膜結(jié)構(gòu),維持細(xì)胞的抗氧化系統(tǒng),使龍眼果實(shí)免受病菌帶來的傷害。因此,研究P型ATP酶基因家族,篩選出該家族中可能響應(yīng)拮抗菌作用信號的關(guān)鍵基因,可能有助于探明拮抗菌保鮮劑的作用機(jī)理,提高采后荔枝的果實(shí)品質(zhì)。
1 材料與方法
1.1 材料
供試?yán)笾ζ贩N為‘妃子笑’,采摘于海南省澄邁縣荔枝種植園。剔除病果、機(jī)械損傷果,留0.5 cm左右果柄、大小均一的荔枝果實(shí)為材料。將挑選后的荔枝均勻分為2組,一組于108CFU/mL拮抗菌N-1懸液中浸泡20 min,另一組用清水浸泡相同時(shí)間用作對照,隨后晾干。每30個(gè)果實(shí)裝一框,用80 cm×120 cm的保鮮膜輕裹,放置于25℃的智能人工氣候(RH:85%~90%)中貯藏,每2 d取樣一次。取30個(gè)果實(shí)用于觀察,并測定好果率和荔枝果皮的ATP酶活性[16],果皮使用液氮研磨,并使用天根RNA提取試劑盒(多糖多酚植物總RNA提取試劑盒DP441)提取荔枝果實(shí)總RNA,使用逆轉(zhuǎn)錄試劑盒(MonScriptTM RTIII All-in-One Mix With dsDNase)將RNA逆轉(zhuǎn)錄成cDNA保存?zhèn)溆谩?/p>
暹羅拮抗菌N-1從自然感病的熱帶水果中分離,隨后保存在–80℃環(huán)境中,該拮抗菌在之前的實(shí)驗(yàn)中已經(jīng)得到鑒定[17]。在使用之前,N-1拮抗菌在28℃條件下于PDA培養(yǎng)基中培養(yǎng)2 d,隨后轉(zhuǎn)移到NA培養(yǎng)基中培養(yǎng)36 h,最后稀釋拮抗菌懸浮液濃度為108CFU/mL,備用。
1.2 方法
1.2.1 荔枝P型ATP酶家族基因的收集與鑒定 利用Pfam數(shù)據(jù)庫(http://pfam.xfam.org/)獲取P型ATP酶家族基因的結(jié)構(gòu)域(PF00690、PF00122、PF13246)的比對文件構(gòu)建隱馬爾可夫模型(HMM)文件。然后,利用TB-tools軟件對荔枝蛋白數(shù)據(jù)庫進(jìn)行HMM搜索。此外,所有P型ATP酶蛋白序列均通過SMART(http://smart.emblhe- idelberg.de/)和NCBI(http://www.ncbi.nlm.n-ih. gov/cdd/)手動篩選結(jié)構(gòu)域,并去除缺乏P型ATP酶結(jié)構(gòu)域的蛋白序列[18]。
1.2.2 系統(tǒng)進(jìn)化樹分析 利用MEGA-X軟件對荔枝、蘋果、獼猴桃、擬南芥、巴旦木、辣椒和蒺藜苜蓿共7個(gè)不同種屬物種的P型ATP酶基因家族全長蛋白序列進(jìn)行比對,對比結(jié)果在Jalview軟件中進(jìn)行美化分析,再使用MEGA-X軟件繪制進(jìn)化樹,并通過FigTree軟件進(jìn)行美化[19]。
1.2.3 保守基序和理化性質(zhì)分析 利用MEME (http://memesuite.org/t-ools/-meme)網(wǎng)站對荔枝P型ATP酶家族基因進(jìn)行motif分析,共設(shè)置15個(gè)motif,其他參數(shù)為默認(rèn)設(shè)置,隨后利用ExPAsy(http://web.expasy.org/com-pute_pi/)在線軟件預(yù)測P型ATP酶基因家族成員蛋白理化性質(zhì)信息,包括氨基酸數(shù)、分子量、等電點(diǎn)、總平均疏水指數(shù);通過Plant-mPLoc(http://www.csbio.sjtu.edu. cn/bioinf/-plant-multi/)在線軟件預(yù)測該基因家族蛋白的亞細(xì)胞定位,并利用GOV IV(http://npsa prabi.-ibcp.fr)在線軟件預(yù)測蛋白二級結(jié)構(gòu)[20]。
1.2.4 基于轉(zhuǎn)錄組的荔枝P型ATP酶家族基因表達(dá)分析 使用荔枝轉(zhuǎn)錄組(FPKM)值表示在各P型ATP酶家族基因的表達(dá)量,并用TBtools軟件繪制熱圖和維恩圖顯示該家族基因在荔枝貯藏過程中的表達(dá)水平。其中熱圖依據(jù)轉(zhuǎn)錄組數(shù)據(jù)FPKM值作圖,以分析荔枝貯藏前期(0、2d)和后期(6、8d)基因表達(dá)差異;維恩圖中差異基因篩選均以0 d為作為參照,以轉(zhuǎn)錄組數(shù)據(jù)-value和log2(FC)值為依據(jù)篩選差異基因。
1.2.5 實(shí)時(shí)熒光定量PCR分析 根據(jù)荔枝P型ATP酶家族基因的ORF序列,在網(wǎng)站(https:// primer3.ut.ee/)上設(shè)計(jì)引物序列,并在DNAMAN軟件上驗(yàn)證引物的可靠性,設(shè)計(jì)好的引物交由中美泰和生物技術(shù)有限公司合成。使用Mona公司試劑盒(MonAmpTM ChemoHS qPCR Mix)進(jìn)行RT-qPCR擴(kuò)增,荔枝P型ATP酶家族基因的相對表達(dá)水平采用2–DDCT法進(jìn)行標(biāo)準(zhǔn)化分析[21],并用GraphPad Prism 8軟件構(gòu)建基因表達(dá)柱狀圖。
1.3 數(shù)據(jù)處理
數(shù)據(jù)采用SAS軟件進(jìn)行統(tǒng)計(jì)分析。
2 結(jié)果與分析
2.1 對荔枝P型ATP酶基因家族的鑒定
通過對荔枝轉(zhuǎn)錄組數(shù)據(jù)進(jìn)行篩選,并結(jié)合在線工具SMART(http://smart.emb-lheidelberg.de/)和NCBI(http://www.ncb-i.nlm.nih.gov/cdd/)的結(jié)構(gòu)域分析,共鑒定出28個(gè)P型ATP酶基因家族成員,并對所有的家族成員進(jìn)行系統(tǒng)編號,將其命名為~。
荔枝28個(gè)P型ATP酶基因家族成員的氨基酸長度范圍在203(LcPA28)~1061(LcPA25)之間不等,相對分子質(zhì)量范圍為50 926.22~ 259 788.49 Da,等電點(diǎn)范圍為4.75~5.14,所有蛋白均為疏水性蛋白,脂肪系數(shù)范圍為17.29~ 29.66,α-螺旋占比最大。亞細(xì)胞定位預(yù)測發(fā)現(xiàn)荔枝大多數(shù)P型ATP酶家族基因定位于線粒體(21/28),其次定位于細(xì)胞膜(5/28)、葉綠體(LcPA15)和細(xì)胞質(zhì)(LcPA22)中(表1)。
2.2 系統(tǒng)進(jìn)化樹分析
通過聚類分析,將荔枝中28個(gè)LcPA蛋白分成5個(gè)亞組,Group Ⅱ、Group Ⅲ和Group Ⅴ均有3個(gè)成員,分別為、和,、和,、和,Group Ⅳ僅有1個(gè)成員,其余18個(gè)成員均屬于Group I(圖1)。
2.3 荔枝P型ATP酶基因家族結(jié)構(gòu)分析
利用MEME分析了荔枝P型ATP酶基因家族蛋白序列的15個(gè)保守motifs(圖2),這些基序的氨基酸組成如表2所示。本研究發(fā)現(xiàn),所有LcPA蛋白均含有motif 2,表明其在荔枝P型ATP酶基因家族中高度保守,此外,高度保守的motif 4同樣存在于除外的所有LcPA蛋白中。在所有15個(gè)motif基序中,值從高到低排在前6的依次是motif 1、motif 4、motif 5、motif 7、motif 6、motif 2,均位于N末端結(jié)構(gòu)域。
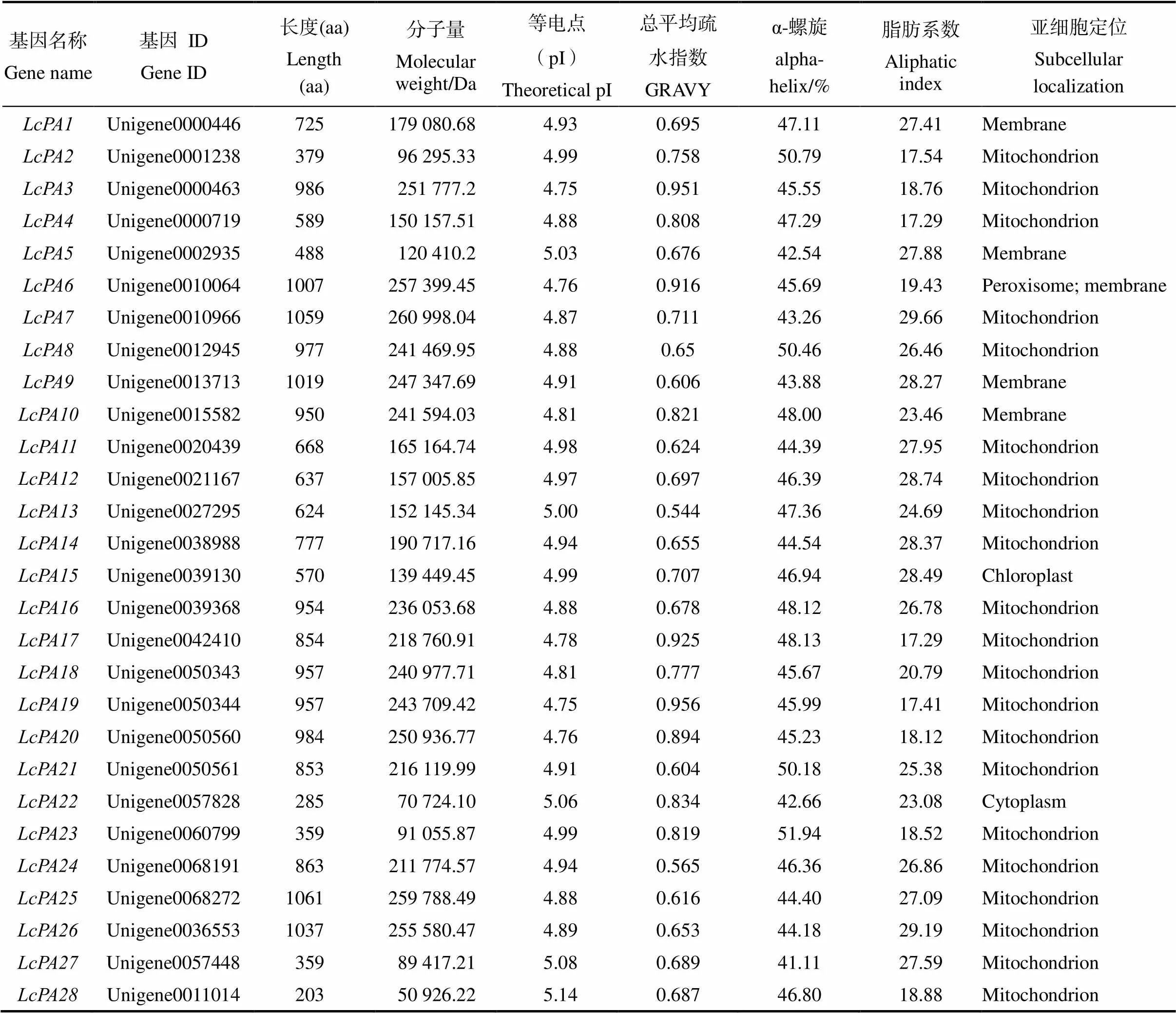
表1 荔枝P型ATP酶基因家族成員信息
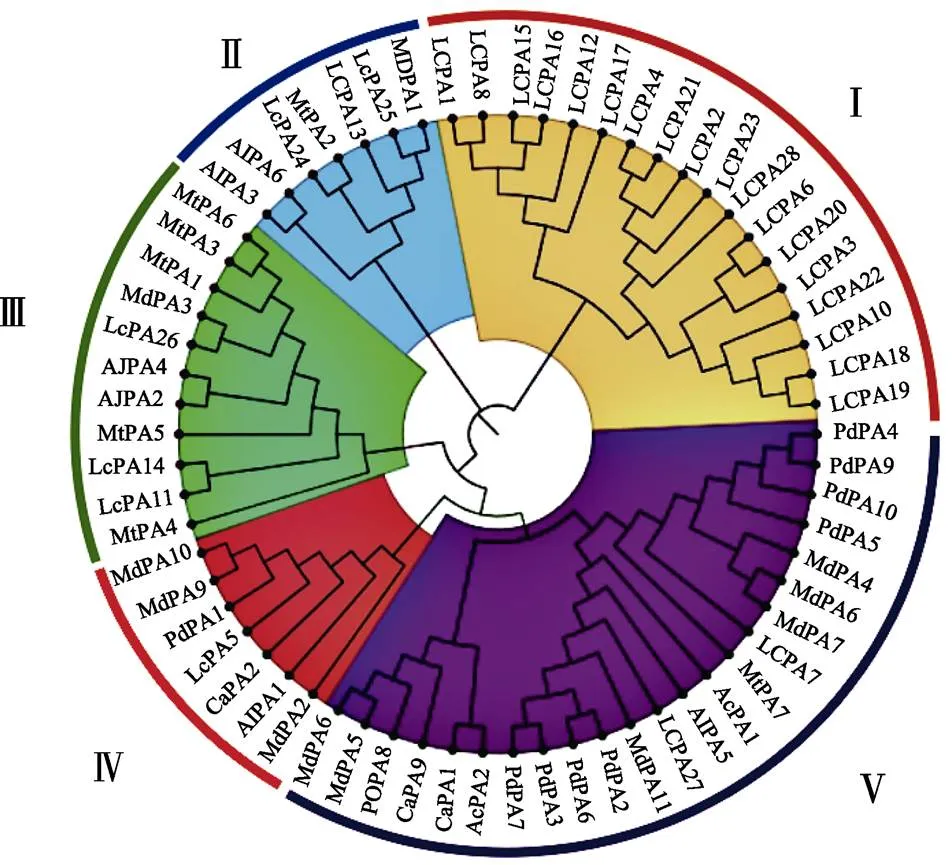
圖1 P型ATP酶基因家族系統(tǒng)進(jìn)化樹分析
2.4 荔枝P型ATP酶家族基因采后貯藏過程中的表達(dá)分析
根據(jù)荔枝P型ATP酶家族基因在荔枝采后貯藏過程中的表達(dá)水平繪制成熱圖(圖3A)和維恩圖(圖3B)。結(jié)果表明,在整個(gè)貯藏過程中出現(xiàn)明顯上調(diào)或下調(diào)表達(dá)的基因占到64.3%,且貯藏后期差異表達(dá)的基因數(shù)量明顯高于前期,上調(diào)表達(dá)基因的數(shù)量顯著高于下調(diào)表達(dá)基因。上調(diào)表達(dá)基因的大量富集暗示著該基因家族基因可能在‘妃子笑’荔枝品質(zhì)劣變過程中起到積極的抵御作用。
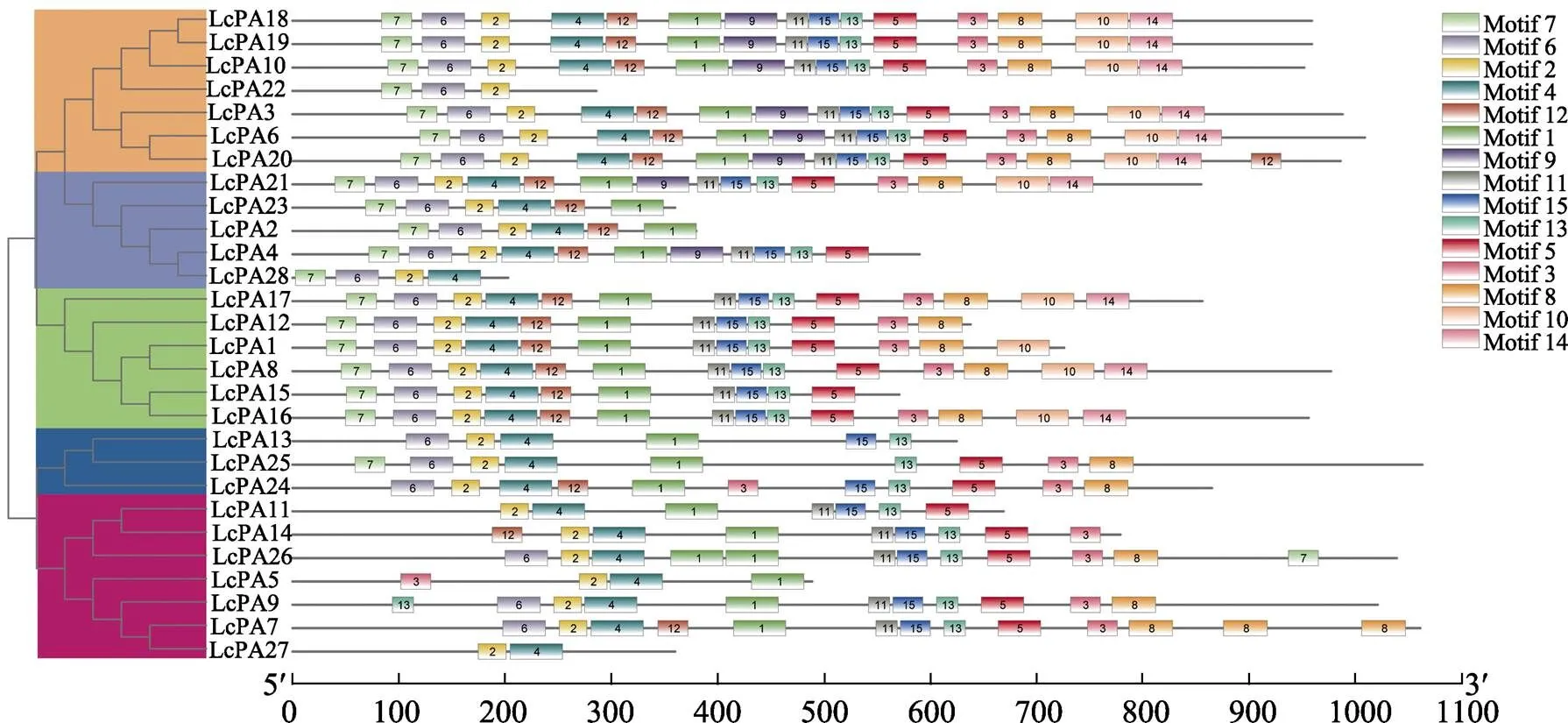
圖2 荔枝P型ATP酶基因家族motif分布

表2 荔枝P型ATP酶基因家族結(jié)構(gòu)域序列標(biāo)識
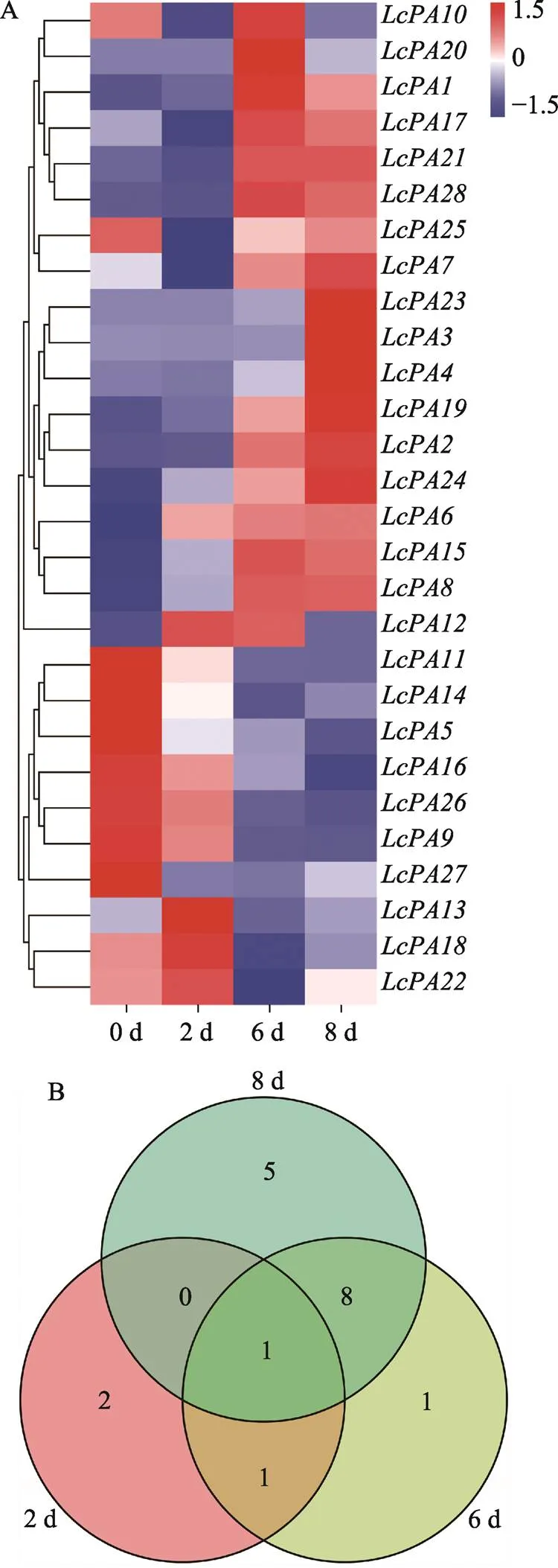
圖3 荔枝P型ATP酶基因家族的表達(dá)分析
2.5 拮抗菌N-1處理下果實(shí)外觀,好果率和ATP酶活性變化
隨著貯藏時(shí)間的增加,果實(shí)的好果率呈現(xiàn)明顯的下降趨勢,但在整個(gè)貯藏過程中,拮抗菌處理后的荔枝果實(shí)的好果率始終高于對照果實(shí)(圖4A,圖4B),從圖4A中可以看出,處理組荔枝果實(shí)的外觀,褐變及病變情況均好于對照組,說明拮抗菌N-1處理可以有效的抑制荔枝果實(shí)的褐變和病變。結(jié)合圖4C可知,拮抗菌處理的荔枝果實(shí)ATP酶活性在整個(gè)貯藏過程中顯著高于對照果實(shí),說明拮抗菌可能通過提高ATP酶活性維持荔枝的高能荷狀態(tài),增強(qiáng)荔枝的抗病性,減少荔枝營養(yǎng)物質(zhì)的損失,最終達(dá)到延緩荔枝果實(shí)衰老腐敗的目的。
2.6 部分LcPA家族基因在拮抗菌N-1處理下的表達(dá)
從LcPA基因家族在采后貯藏過程中的表達(dá)分析中初步篩選出在‘妃子笑’荔枝貯藏過程中可能起到關(guān)鍵作用的家族基因,并對這些基因在拮抗菌N-1處理后的表達(dá)情況進(jìn)行實(shí)時(shí)熒光定量PCR分析(圖5)。結(jié)果表明,拮抗菌N-1顯著誘導(dǎo)了基因和在整個(gè)貯藏過程中的上調(diào)表達(dá),誘導(dǎo)基因、在荔枝貯藏第2天上調(diào)表達(dá),并誘導(dǎo)基因、和在貯藏后期的上調(diào)表達(dá),總體而言,拮抗菌N-1處理能誘導(dǎo)荔枝P型ATP酶基因家族基因的上調(diào)表達(dá),這與ATP酶活性的變化一致。以上結(jié)果說明,荔枝LcPA家族基因在拮抗菌N-1抑制荔枝病變的過程中發(fā)揮著積極的作用,且響應(yīng)外源刺激信號的時(shí)期和作用也有所不同。
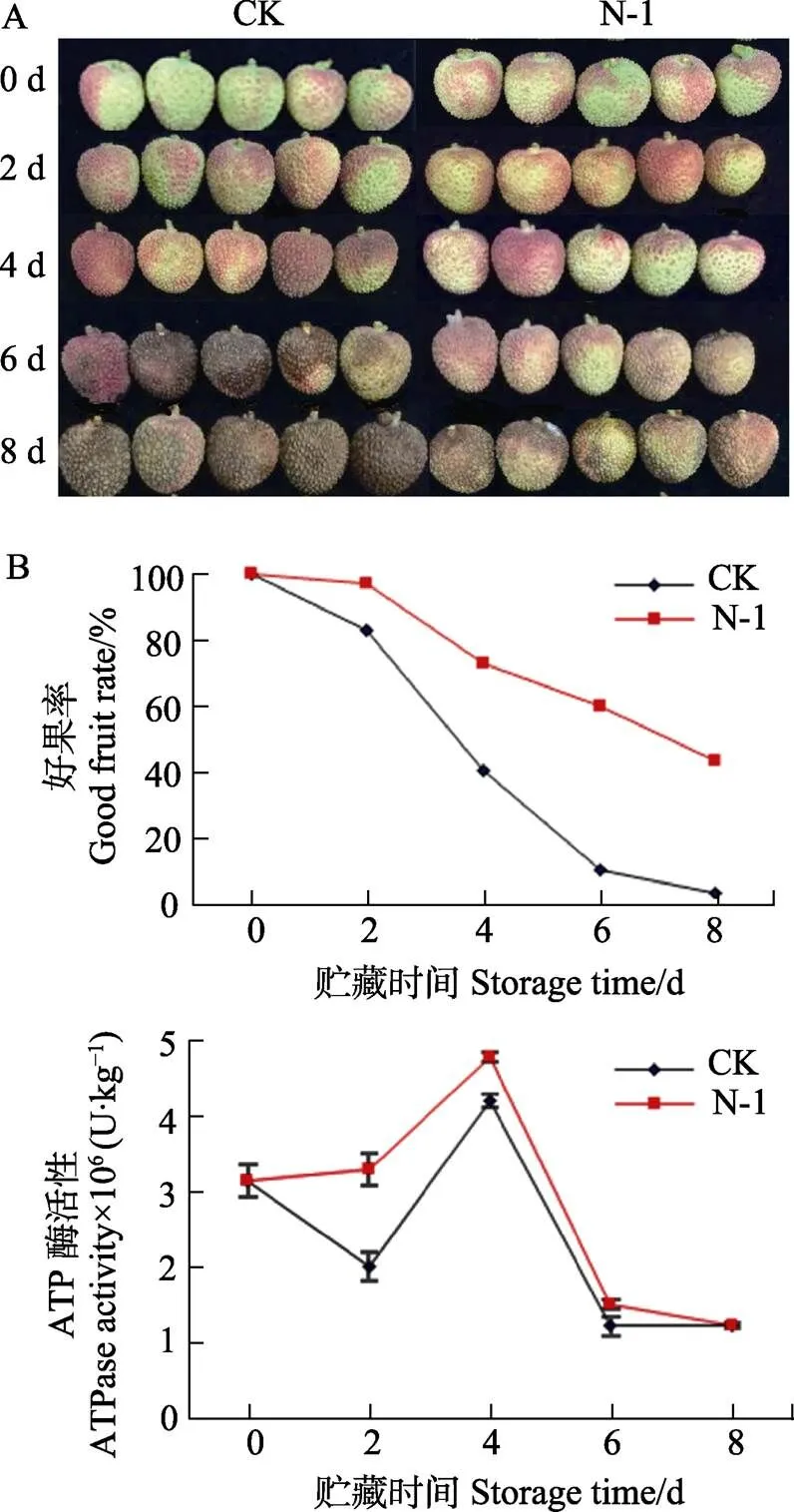
圖4 拮抗菌N-1處理下果實(shí)好果率和ATP酶活性變化
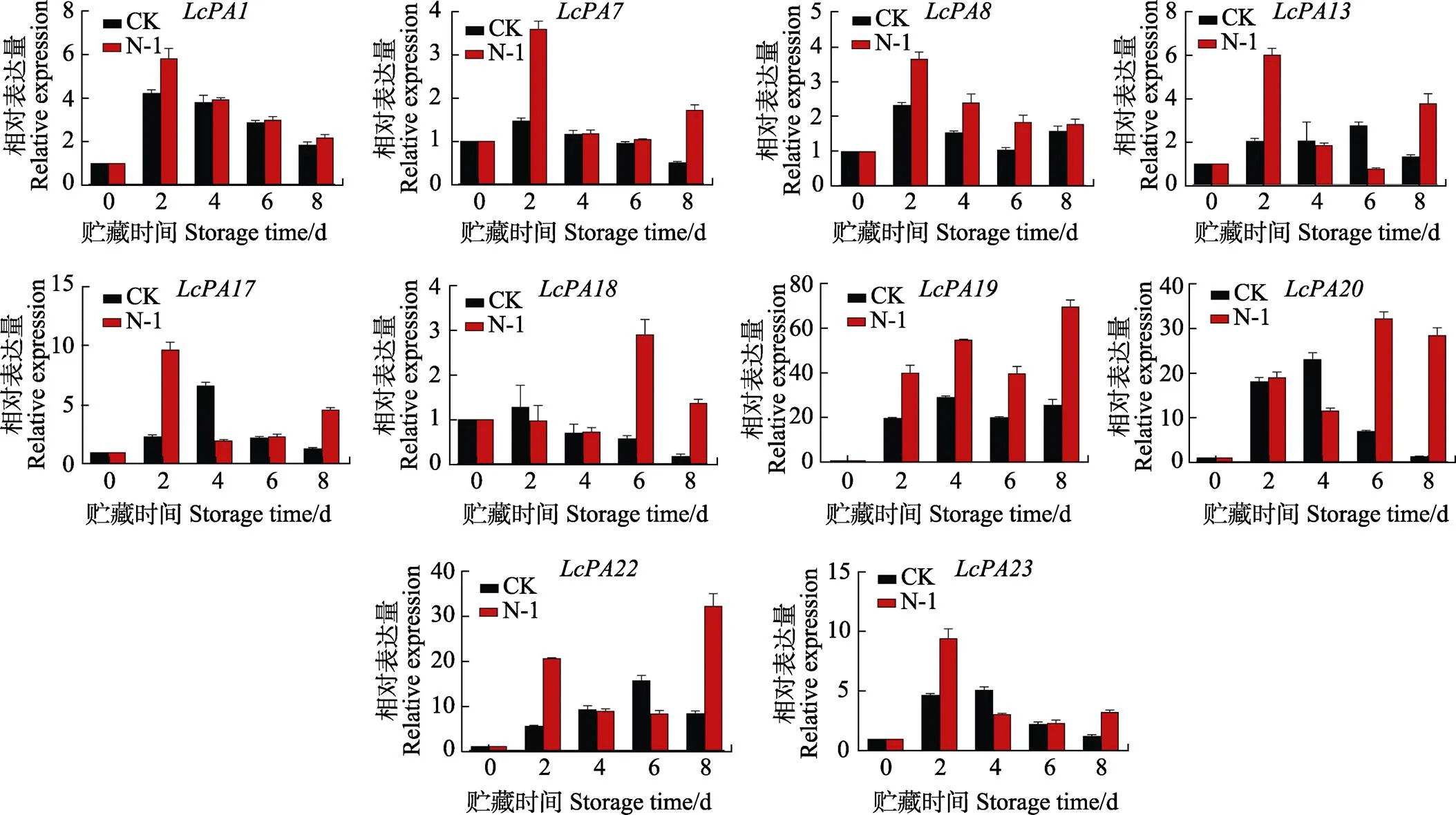
圖5 荔枝P型ATP酶基因家族成員在拮抗菌處理下的表達(dá)
3 討論
3.1 荔枝P型ATP酶家族基因的鑒定與結(jié)構(gòu)分析
采后果實(shí)是一個(gè)鮮活的有機(jī)體,時(shí)刻進(jìn)行著各種代謝活動,P型ATP酶基因家族在果實(shí)的能量代謝、酸代謝、脅迫信號轉(zhuǎn)導(dǎo)、逆境響應(yīng)等過程中發(fā)揮著重要的作用,目前已在蘋果[22]、柑橘[23-24]和梨[25]等多個(gè)物種中得到鑒定和研究。在本研究中,基于‘妃子笑’荔枝的轉(zhuǎn)錄組數(shù)據(jù),共鑒定出28個(gè)荔枝P型ATP酶家族家族基因,并劃分為5個(gè)亞族,說明該家族在進(jìn)化上可能存在5個(gè)分支,這在擬南芥P型ATP酶基因家族的鑒定上也得到了驗(yàn)證[1],這28個(gè)家族基因?qū)?yīng)蛋白的等電點(diǎn)、總平均疏水指數(shù)、α-螺旋和脂肪系數(shù)在荔枝中高度一致,表明該家族成員在進(jìn)化上比較保守,在motif的分布模式中,值高的motif基序基本分布于N-末端結(jié)構(gòu)域,說明N-末端結(jié)構(gòu)域比C-末端結(jié)構(gòu)域更加保守。荔枝P型ATP酶基因家族的亞細(xì)胞定位預(yù)測大多位于線粒體,而線粒體能夠參與植物細(xì)胞內(nèi)的諸多過程,如呼吸作用、產(chǎn)熱、抗逆機(jī)制、程序性細(xì)胞死亡和基因組進(jìn)化等[26],另外還有少量基因定位于葉綠體、細(xì)胞膜和細(xì)胞質(zhì)中,說明該基因家族不僅在能量代謝過程中發(fā)揮重要作用,還在植物的各種生理活動中發(fā)揮作用,也說明了該基因家族功能的多樣性。
3.2 P型ATP酶家族基因在采后荔枝貯藏保鮮過程中發(fā)揮重要作用
ATP酶作為能量代謝的關(guān)鍵酶,貫穿于細(xì)胞的整個(gè)生命過程。采后果實(shí)由于脫離了母體,切斷了能量供應(yīng),其貯藏的能量又被不斷消耗,已有諸多研究證實(shí),果實(shí)的能量虧損越嚴(yán)重,品質(zhì)劣變程度越嚴(yán)重,其衰老和死亡的速度和進(jìn)程越快[27]。高兆銀等[28]研究發(fā)現(xiàn)外源ATP處理可提高荔枝貯藏前期ATP合成酶基因表達(dá)量,保持荔枝果實(shí)的能量供應(yīng),顯著延緩荔枝果實(shí)的成熟衰老,這與本研究中拮抗菌處理誘導(dǎo)基因、和在荔枝貯藏第2天上調(diào)表達(dá)的結(jié)果相似,說明這些基因可能是最先響應(yīng)拮抗菌處理觸發(fā)的信號。基因和則在整個(gè)貯藏過程中均被拮抗菌顯著誘導(dǎo)高表達(dá),這些基因均定位于線粒體,研究表明,線粒體ATP合成酶是細(xì)胞功能的關(guān)鍵樞紐,控制著ATP的產(chǎn)生和細(xì)胞信號傳導(dǎo),它受ATP酶抑制因子1(IF1)的調(diào)節(jié),從而控制線粒體功能和線粒體活性氧的產(chǎn)生[29],這在生理水平上也得到了驗(yàn)證,解偶連劑DNP可加劇活性氧的積累,降低能荷水平從而加速草莓衰老[30]。在拮抗菌的誘導(dǎo)下,荔枝線粒體ATP酶基因上調(diào)表達(dá),荔枝ATP酶活性升高,其保護(hù)機(jī)制可能也是通過抑制線粒體活性氧的產(chǎn)生,進(jìn)而保護(hù)細(xì)胞膜結(jié)構(gòu),最終抑制了荔枝果實(shí)品質(zhì)劣變,而基因基因和可能是維持荔枝高能荷狀態(tài)、抑制荔枝果實(shí)品質(zhì)劣變的關(guān)鍵基因。ATP除了作為能量貨幣的功能外,還起著胞外信號分子的作用,當(dāng)果實(shí)ATP含量過低時(shí),會產(chǎn)生反饋調(diào)節(jié)[31],基因、和在荔枝貯藏后期的上調(diào)表達(dá)說明其可能是ATP反饋調(diào)節(jié)的響應(yīng)基因。本研究表明以上幾個(gè)基因在荔枝采后貯藏保鮮過程中發(fā)揮著重要作用,可作為后續(xù)的研究重點(diǎn),進(jìn)行基因功能的驗(yàn)證。
[1] KUHLBRANDT W. Biology, structure and mechanism of P-type ATPases[J]. Nature Reviews Molecular Cell Biology, 2004, 5(4): 282-295.
[2] XKRISTIAN B. A, MICHAEL G. P. Inventory of the superfamily of P-type ion pumps in[J]. Plant Physiology, 2001, 126(2): 696-706.
[3] XU Z C, MAROWA P, LIU H, DU H N, ZHANG C S, LI Y Q. Genome-wide identification and analysis of P-type plasma membrane H+-ATPase sub-gene family in sunflower and the role ofandin the development of salt stress resistance[J]. Genes, 2020, 11(4): 361.
[4] CHEN W, SI G Y, ZHAO G, ABDULLA H M, GUO N, LI D H, SUN X, CAI Y P, LIN Y, GAO J S. Genomic comparison of the P-ATPase gene family in four cotton species and their expression patterns in[J]. Molecules, 2018, 23(5): 1092.
[5] ZHAO B Q, WU H C, XU W J, ZHANG W, CHEN X, ZHU Y Y, CHEN H T, ZENG H. Q. Genome-wide identification, characterization, and expression analyses of P-Type ATPase superfamily genes in soybean[J]. Agronomy, 2020, 11(1): 543.
[6] IVAN B, JASON T, MICHAEL R, MARC B, MICHAEL G. P, MICHAEL G, JEFFREY F. H, N KRISTIAN B. Genomic comparison of P-type ATPase ion pumps in arabidopsis and rice[J]. Plant Physiology, 2003, 132(2): 618-628.
[7] LI Y B, PROVENZANO S, BLIEK M, SPELT C, APPE- LHAGEN I, MACHADOL, VERWEIJ W, SCHUBERT A, SAGASSER M, SEIDEL T, WEISSHAAR B, KOES R, QUATTROCCHIO F. Evolution of tonoplast P‐ATP are transporters involved in vacuolar acidification[J]. New Phytologist, 2016, 211(3): 761-764.
[8] STRAZZER P, CORNELIS E. S, LI S J, BLIEK M, CLAIRE T. F, MIKEAL L. R, KOES R, FRANCESCA M. Q. Hyperacidification of citrus fruits by a vacuolar proton-pumping P-ATPase complex[J]. Nature Communications, 2019, 10(1): 744.
[9] APPELHAGEN I, NORDHOLT N, SEIDEL T, SPELT K, KOES R, QUATTROCHIO F, SAGASSER M, WEISSHAAR B. TRANSPARENT TESTA 13 is a tonoplast P3A-ATP ase required for vacuolar deposition of proantho cyanidins inseeds[J]. The Plant Journal, 2015, 82(5): 840-849.
[10] HUDA K M K , YADAV S , BANU M S A , TUTEJA T N. Genome wide analysis of P-Type-II Ca2+ATPases gene family from rice and arabid-opsis: potential role in abiotic stresses[J]. Plant Physiology and Biochemistry, 2013, 65: 32-47.
[11] TianZ h,Li Y Y, Qian L P,D Y j,ChenC w,Chao a L. Antifungal activities and the mechanisms of biocontrol agent WE-3 against postharvest sour rot in citrus[J]. European Journal of Plant Pathology,2021,161(3): 723-733.
[12] SIVAKUMAR D, ZEEMAN K, KORSTEN L. Effect of a biocontrol agent () and modified atmosphere packaging on postharvest decay control and quality retention of litchi during storage[J]. Phytoparasitica, 2007, 35(5): 507-518.
[13] 張正敏, 楊藝琳, 李美琳, 王 靜, 季娜娜, 鄭永華. 2,4-表油菜素內(nèi)酯處理對桃果實(shí)軟腐病及能量代謝的影響[J]. 食品科學(xué), 2019, 40(5): 207-213.
ZHANG Z M, YANG Y L, LI M L, WANG J, JI N N, ZHENG Y H. Effect of 2,4-epibrass-inolide treatment on rhizopus rot and energy metabolism in postharvest peach fruit[J]. Food Science, 2019, 40(5): 207-213. (in Chinese)
[14] CAO G, Y H, WEI M L, LI C Y, CHEN Y R, LV J Y, LI J R. Effect of acibenzolar-S-methyl on energy metabolism and blue mould of Nanguo pear fruit[J]. Scientia Horticulturae, 2017, 225: 221-225.
[15] CAO S F, CAI Y T, YANG Z F, JOYCE D C, ZHENG Y H. Effect of MeJA treatment on polyamine, energy status and anthracnose rot of loquat fruit[J]. Food Chemistry, 2014, 145: 86-89.
[16] 陳夢茵. DNP和ATP調(diào)控Chi侵染所致龍眼果實(shí)采后病害發(fā)生的機(jī)理研究[D]. 福州: 福建農(nóng)林大學(xué), 2015: 16-20.
CHEN M Y. Study on the mechanism of DNP and ATP regulation the disease development of harvested longan fruit infected byChi[D]. Fuzhou: Fujian Agriculture and Forestry University, 2015: 16-20. (in Chinese)
[17] YOU W, GE C, JIANG Z, CHEN M, SHAO Y. Screening of a broad-spectrum antagonist-, and its possible mechanisms to control postharvest disease in tropical fruits[J]. Biological Control, 2021, 157(11): 104584.
[18] 劉志鑫, 孫 宇, 葉 子, 羅睿雄, 李 忠, 蒲金基, 張 賀. 芒果VOZ轉(zhuǎn)錄因子基因家族成員鑒定及生物信息學(xué)分析[J]. 南方農(nóng)業(yè)學(xué)報(bào), 2021, 52(7): 1762-1770.
LIU Z X, SUN Y, YE Z, LUO R X, LI Z, PU J J, ZHANG H. Identification and bioinformatics analysis of VOZ transcription factor gene family members in[J]. Journal of Southern Agriculture, 2021, 52(7): 1762-1770. (in Chinese)
[19] 林 歡, 段偉科, 周 怡, 祝夢全, 王云鵬, 孫 敏, 黃志楠. 辣椒CDPK基因家族的鑒定、進(jìn)化與表達(dá)分析[J]. 核農(nóng)學(xué)報(bào), 2021, 35(1): 7-17.
LIN H, DUAN W K, ZHOU Y, ZHU M Q, WANG Y P, SUN M, HUANG Z N. Identification, evolution and expression analysis of the CDPK gene family in pepper[J]. Journal of Nuclear Agricultural Sciences, 2021, 35(1): 7-17. (in Chinese)
[20] 邵正偉, 曾志鵬, 陳彥竹, 何敏紅, 張 毅, 陳善蘭, 朱宏波.甘薯全基因組SBP-box基因家族鑒定及表達(dá)分析[J]. 分子植物育種, 2021, 5: 1-20.
SHAO Z W, ZENG Z P, CHEN Y Z, HE M H, ZHANG Y, CHEN S L, ZHU H B. Genome-wide identification and expression analysis of S-BP-box gene family in sweet potato[J]. Molecular Plant Breeding, 2021, 5: 1-20. (in Chinese)
[21] 黃金鳳, 呂天星, 王 尋, 王穎達(dá), 王冬梅, 閆忠業(yè), 劉 志. 蘋果LRR-RLK基因家族鑒定和表達(dá)分析[J]. 中國農(nóng)業(yè)科學(xué), 2021, 54(14): 3097-3112.
HUANG J F, LV T X, WANG X, WANG Y D, WANG D M, YAN Z Y, LIU Z. Genome-wide identification and expression analysis of LRR-RLK gene family in apple[J]. Agricultural Sciences in China, 2021, 54(14): 3097-3112. (in Chinese)
[22] 陳厚彬, 蘇鉆賢. 2021年全國荔枝生產(chǎn)形勢分析[J]. 中國熱帶農(nóng)業(yè), 2021(2): 5-18.
CHEN H B, SU Z X. Analysis of national litchi production situation in 2021[J]. Chinese Tropical Agriculture, 2021(2): 5-18. (in Chinese)
[23] SHI C Y, SONG R Q, HU X M, LIU X, JIN L F. LIU Y Z. Citrus PH5-like H+-ATPase genes: identification and transcript analysis to investigate their possible relationship with citrate accumulation in fruits[J]. Frontiers in Plant Science, 2015, 6: 135.
[24] SHI C Y, HU S B, HAN H, SM ALAM, LIU D, LIU Y Z. Reduced expression of, a P-type ATPase gene, is the major factor leading to the low citrate accumulation in citrus leaves-science direct[J]. Plant Physiology and Biochemistry, 2021, 160: 211-217.
[25] ZHANG Y X, LI Q H, XU L L, QIAO X, LIU C X, ZHANG S L. Comparative analysis of the P-type ATPase gene family in seven Rosaceae species and an expression analysis in pear (Rehd.)[J]. Genomics, 2020, 112(3): 2550-2563.
[26] IANM M, ALLAN G R, OLIVIER V A. Plant mitochondria -past, present and future[J]. The Plant journal, 2021, 108(4): 912-959.
[27] 王 鑫, 李 雯. 果實(shí)能量代謝與采后品質(zhì)關(guān)系的研究進(jìn)展[J]. 分子植物育種, 2021, 11: 1-13
WANG X, LI W. Research progress on the relationship between fruit energy metabolism and quality deterioration on fruit[J]. Molecular Plant Breeding, 2021, 11: 1-13. (in Chinese)
[28] 高兆銀, 胡美姣, 趙 超, 張正科, 李 敏, 李煥苓, 王 果, 孫進(jìn)華, 文 婕, 王家保. 荔枝能荷水平及能量相關(guān)基因表達(dá)與采后褐變關(guān)系的研究[J]. 園藝學(xué)報(bào), 2018, 45(1): 51-60.
GAO Z Y, HU M J, ZHAO C, ZHANG Z K, LI M, LI H L, WANG G, SUN J H, WEN J, WANG J B. Effects of energy charge levels and ex-pression of energy charge genes on browning of post-harvest litchi fruit[J]. Acta Horticulturae Sinica, 2018, 45(1): 51-60. (in Chinese)
[29] 梁 爽, 劉 歡, 李巧玲, 白 楊, 閆宇彤, 郭金麗. 能量調(diào)控對草莓果實(shí)采后衰老過程中活性氧及保護(hù)酶的影響[J].安徽農(nóng)業(yè)大學(xué)學(xué)報(bào), 2016, 43(5): 810-814.
LIANG S, LIU H, LI Q L, BAI Y, YAN Y T, GUO J L. Effects of energy regulating on reactive oxygen and protective enzyme activity in ageing process of strawberry fruit[J]. Journal of Anhui Agricultural Sciences, 2016, 43(5): 810-814. (in Chinese)
[30] ESPARZA M P B, ROMERO C I, Nú?EZ D A C, PEREIRA M P, BLANCO N, PARDO B, BATES G, SáNCHEZ C C, ARTUCH R, MURPHY M, ESTEBAN J, CUEZVA J. Generation of mitochondrial reactive oxygen species is controlled by ATPase inhibitory factor 1 and regulates cognition[J]. PLoS biology, 2021, 19(5): e3001252.
[31] MORTEZA S A, ABBASALI J, ZISHENG L, GOPINADHAN P. Ensuring sufficient intracellular ATP supplying and friendly extracellular ATP signaling attenuates stresses, delays senescence and maintains quality in horticultural crops during postharvest life[J]. Trends in Food Science & Technology, 2018, 76: 67-81.
Identification of P-type ATPase Gene Family and Analysis of Expression Pattern in Response to Antagonistic N-1 During Postharvest Storage in Litchi
WANG Xin1, SHAO Yuanzhi2, TANG Yue2, LI Wen1*
1. Key Laboratory for Quality Regulation of Tropical Horticultural Crops of Hainan Province, School of Horticulture, Hainan University, Haikou, Hainan 570228, China; 2. School of Life Sciences, Hainan University, Haikou, Hainan 570228, China
P-type ATPase is an important membrane transporter in plants. It plays a significant role in plant energy metabolism and acid metabolism. In addition, the enzyme is widely involved in plant ion transport, stress resistance, cell signal transduction and other life activities. However, p-type ATPase gene family has not been comprehensively analyzed in litchi or other sapindaceae plants. In this study, based on the transcriptome data of ‘Feizixiao’ litchi, 28 p-type ATPase genes were identified through physiological and biochemical analysis, bioinformatics analysis and expression analysis, to explore the potential function of P-type ATPase during postharvest storage of litchi. The structure and phylogenetic analysis showed that the gene could be divided into 5 subfamilies, the genes in the same subfamily had similar gene structure and motifs. The secondary structure of protein was mainly consisted of α -helix, and the N-terminal domain of LcPAs was more conserved than the C-terminal domain. Subcellular localization prediction analysis showed that LcPAs were mostly located in mitochondria (21/28). According to transcriptome analysis, the up-regulated genes of P type ATPase family genes in litchi were significantly higher than the down-regulated genes during postharst storage, which suggesting that they may play a positive role in the resistance of quality deterioration of litchi after harvest. As a safe and healthy preservation technology, the mechanism of antagonistic biological control is very complex. Results showed that the antagonistic N-1 treatment could effectively inhibit the browning and disease occurrence of litchi fruits after harvest. Combined with RT-qPCR results, it was found that antagonistic N-1 treatment could induce up-regulated expression of,,andduring the whole storage process, which further indicating that P-type ATPase family genes would play an important role in the antagonistic N-1 inhibition of litchi lesions. This study would provide a reference for understanding the evolution and biological function analysis of p-type ATPase gene family in litchi, it also provide new evidence for further understanding the molecular regulation mechanism of antagonistic N-1 preservation mechanism.
litchi; p-type ATPase gene family; transcriptome; antagonist N-1
S667.1
A
10.3969/j.issn.1000-2561.2022.11.021
2022-02-25;
2022-04-04
海南省自然科學(xué)基金創(chuàng)新研究團(tuán)隊(duì)項(xiàng)目(No. 320CXTD430);海南省研究生創(chuàng)新科研課題(No. Qhys2021-244)。
王 鑫(1994—),男,碩士研究生,研究方向:荔枝采后貯藏保鮮。*通信作者(Corresponding author):李 雯(LI Wen),E-mail:liwen9-210@163.com。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財(cái)經(jīng)界(學(xué)術(shù)版)(2015年20期)2015-12-23 09:20:13
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
