
青稞硝酸鹽轉運蛋白基因HvnNPF4.5 的克隆和亞細胞定位
2022-12-20 13:12:26劉凡語安立昆姚曉華姚有華崔永梅白羿雄吳昆侖
湖南農業科學 2022年11期
劉凡語,安立昆,姚曉華,姚有華,崔永梅,白羿雄,李 新,吳昆侖
(青海大學農林科學院,青藏高原種質資源研究與利用實驗室,青海省青稞遺傳育種重點實驗室,國家麥類改良中心青海青稞分中心,青海 西寧 810016)
青稞(Hordeum vulgareL. var.nudumHook. f.)是青藏高原地區最具特色的作物,是當地居民賴以生存的糧食和飼料、食品加工原料,在青藏高原農業歷史文化和農業經濟發展中有著重要的地位[1-4]。青藏高原地區氣候惡劣,青稞在青藏高原有著悠久的栽培歷史,雖然青稞已經適應了青藏高原的惡劣條件,但是各種逆境脅迫,尤其是貧瘠的土壤依然嚴重制約著青稞的種植和生產[5-8]。氮元素是影響作物生長發育和產量品質的重要大量元素之一。在作物的種植過程中為了提高產量,會施用大量的氮肥。然而,氮肥施用過量和氮肥流失一直是困擾我國農業生產的重大問題[9]。長期過量施用氮肥會造成土壤結構破壞,流失的氮肥還會污染農田周邊環境,不利于生態農業可持續發展[10]。因此,減少農業生產中氮肥的施用量、提高作物對氮肥的利用效率對于農業可持續發展具有重要意義。青稞主要種植地集中在生態極為脆弱敏感的青藏高原地區,長期大量施用氮肥會對青藏高原地區生態環境平衡造成嚴重破壞[11-13]。因此,在保證青稞產量的同時,盡可能減少青稞生產過程中氮肥的施用,對青藏高原農業生態的可持續發展具有重要意義。
植物硝酸鹽轉運蛋白家族NPFs 是廣泛存在于各種植物中的一類低親和硝酸鹽轉運蛋白家族,其主要功能是在植物體受到低氮脅迫時對硝酸鹽進行吸收和轉運,此外還參與植物次生代謝產物以及激素等物質的轉運[14-15]。很多研究表明,NPFs 家族中一些成員參與菌根調控的硝酸鹽吸收和同化過程。楊曉鋒[16]研究水稻硝酸鹽轉運蛋白基因OsNPF4.5時發現,OsNPF4.5位于水稻第1 條染色體上,具有7 個外顯子和6 個內含子,可編碼609 個氨基酸,亞細胞定位結果顯示其蛋白位于細胞膜上,該基因的表達受叢枝菌根強烈誘導,而同一家族的其他基因表達并不受叢枝菌根的影響,此外該基因的表達受外源硝酸鹽的誘導,外源的脫落酸和水楊酸會抑制其表達。Wang 等[17]發現水稻OsNPF4.5 蛋白定位于細胞膜上,與玉米ZmNPF4.5 和高粱SbNPF4.5 親緣關系最近,是一種參與水稻菌根吸收和同化硝酸鹽的低親和硝酸鹽轉運蛋白;將OsNPF4.5轉入水稻中,轉基因植株的氮素吸收效率明顯高于野生型植株,且生長速度加快,其菌根途徑貢獻的氮大約有45%通過OsNPF4.5 轉運,表明OsNPF4.5 在禾本科植物通過菌根途徑吸收利用硝酸鹽的過程中起主導作用。黃圣宇[18]對枳硝酸鹽轉運蛋白基因PtrNPF5.2進行了研究,發現PtrNPF5.2具有12 個跨膜結構,在第6~7 跨膜域間存在一個大親水環,其蛋白定位于細胞膜上,符合硝酸鹽轉運蛋白家族的典型特征,PtrNPF5.2主要在與叢枝菌共生的細胞中表達,表明該基因可能參與菌根調控的硝酸鹽吸收和同化過程。馮慧敏等[19]的研究表明,水稻OsNPF7.9基因表達的蛋白定位于細胞膜上,有12 個跨膜結構,在葉片、花和根中都有表達,但其表達不受硝態氮或銨態氮及其濃度差異的影響,將其轉入水稻中發現轉基因植株由根系向地上部的硝酸鹽轉運能力增強,表明OsNPF7.9 在植物將硝酸鹽從根系向地上部轉運的過程中發揮著重要的作用。夏秀東[20]的研究表明,水稻硝酸鹽轉運蛋白基因OsNPF2.4由2個內含子和3 個外顯子組成,其蛋白由583 個氨基酸組成,有12 個跨膜結構,定位于質膜上,在葉鞘、葉片、花粉和剛萌發的幼芽中均有表達,主要在根的表皮細胞、木質部薄壁細胞等中表達,另外,該基因敲除突變體對NO3-的吸收量明顯低于野生型,說明OsNPF2.4的敲除會影響水稻體內的運輸,而其過表達會顯著增加水稻對的吸收。
目前,關于植物硝酸鹽轉運蛋白基因NPFs家族的研究在青稞中尚未見報道。該研究從 “昆侖14”青稞中克隆得到青稞硝酸鹽轉運蛋白基因HvnNPF4.5,對其基因和蛋白結構進行了生物信息學分析,并對其亞細胞定位進行了研究,以期為青稞硝酸鹽轉運蛋白進一步研究利用提供參考。
1 材料與方法
1.1 試驗材料
供試青稞品種“昆侖14”以及對照品種本氏煙草(Nicotiana benthamiana)均由青藏高原種質資源研究與利用實驗室保存。TransZol Up Plus RNA 提取試劑盒和PCR 產物連接及cDNA 合成試劑盒EasyScript First-Strand cDNA Synthesis SuperMix(AE301-03)購自北京全式金生物技術有限公司。高保真PCR酶PrimeSTAR 和Mighty TA-cloning Reagent Set for PrimeSTAR 克隆試劑盒購自Takara 寶生物工程(大連)有限公司。Gateway 克隆試劑盒購自Invitrogen 公司。大腸桿菌感受態菌株為自制DH5α 感受態。引物合成和測序由上海生工生物工程有限公司完成。
1.2 試驗方法
1.2.1 青稞DNA 和RNA 提取采用CTAB 法從青稞葉片中提取DNA,采用TransZol Up Plus RNA 提取試劑盒從青稞葉片中提取RNA,并參照cDNA 合成試劑盒合成cDNA,所有樣品于-20℃冰箱保存。
1.2.2 青稞HvnNPF4.5 基因克隆和啟動子區域序列克隆將已報道的水稻OsNPF4.5基因序列輸入NCBI (https://www.ncbi.nlm.nih.gov/)中 搜 索 得 到大麥HvNPF4.5,根據其基因和起始密碼子ATG 前2 000 bp 序列設計引物。基因克隆正向引物F:5'-ATG TACTTCATGTTGGTGATG-3',反向引物R:5'-CTACAT GACAGACTTGCTG-3'。啟動子區域序列克隆正向引物F:5'-CGTGCGTGCTAACGTTCT-3',反向引物F:5'-GA TAAGGTTCACGGCTAGA-3'。以青稞基因組cDNA 和DNA 為模板分別克隆青稞HvnNPF4.5序列和啟動子區域序列。PCR 程序為:94℃ 2 min;98℃ 10 s,58℃30 s,68℃ 1 min(擴增啟動子區域序列為2 min),35個循環;4℃保存。將PCR 產物與TA 克隆載體連接并轉化大腸桿菌DH5α,篩選陽性克隆進行測序。
1.2.3 青稞HvnNPF4.5 生物信息學分析使用GSDS 2.0(http://gsds.gao-lab.org/)對基因結構進行預測。使用Plant CARE(http://bioinformatics.psb.ugent.be/web tools/plantcare/html/)對啟動子區域元件進行預測分析。使 用Protparam(https://web.expasy.org/protparam/) 對蛋白質的物理和化學性質進行預測。使用kinasephos2(http://kinasephos.mbc.nctu.edu.tw/index.php)對蛋白磷酸化位點進行預測。使用SignalP 5.0 服務器(http://www.cbs.dtu.dk/services/SignalP/)對蛋白信號肽進行預測分析。使用TMHMM-2.0 (http://www.cbs.dtu.dk/services/TMHMM-2.0/)對跨膜結構進行預測。使用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)對蛋白預測二級結構進行分析。使用SWISS-MODEL(https://swissmodel.expasy.org/)進行同源建模,生成三級結構再建模。
在NCBI 中搜索得到擬南芥(Arabidopsis thaliana)、甘藍型油菜(Brassica napusL.)、節節麥(Aegilops tauschii subsp.tauschii)、水稻(Oryza sativaL.)、二穗短柄草(Brachypodium distachyon)、玉米(Zea maysL.)、羊黍(Panicum hallii)、黍稷(Panicum miliaceum)、柳枝稷(Panicum virgatum)、二型花(Dichanthelium oligosanthes)、小米(Setaria italica)、狗尾草(Setaria viridis)、南荻(Miscanthus lutarioriparius)、高粱(Sorghum bicolor)、大麥(Hordeum vulgareL.)、康藏嵩草(Carex littledalei)、蘆筍(Asparagus officinalis)、山藥(Dioscorea cayenensissubsp.rotundata)、油棕(Elaeis guineensis)、海棗(Phoenix dactylifera)、野芭蕉(Musa acuminatasubsp.malaccensis)、無 油 樟(Amborella trichopoda)、沉水樟(Cinnamomum micranthum)、毛果楊(Populus trichocarpa)、蓖麻(Ricinus communis)、甘藍型油菜(Brassica napus)、蘿卜(Raphanus sativus)、白菜型油菜(Brassica rapa)、鹽芥(Eutrema salsugineum)、亞 麻 芥(Camelina sativa)、薺 菜(Capsella rubella)的NPF4.5 蛋白。將擬南芥AtNPF4.5、甘藍型油菜BnNPF4.5、節節麥AtsNPF4.5、水稻OsNPF4.5、二穗短柄草BdNPF4.5、玉米ZmNPF4.5 的蛋白序列,采用DNAMAN7.0 與大麥HvNPF4.5 和青稞HvnNPF4.5蛋白序列進行比對,并根據NCBI 提供的蛋白信息分析各蛋白的保守結構域。將所有物種的NPF4.5 蛋白序列輸入MEGA7 中以最大似然法構建系統進化樹。
1.2.4 青稞HvnNPF4.5 亞細胞定位研究采用載體pSATN1-mkate 利用gatway 方法構建亞細胞定位載體。設計帶有attB 位點的引物,正向引物F:5'-GGGGAC AAGTTTGTACAAAAAAGCAGGCTATGTACTTCATG TTGGTGATG-3',反向引物R:5'-GGGGACCACTTTG TACAAGAAAGCTGGGTCATGACAGACTTGCTG-3'。對已克隆的HvnNPF4.5CDS 序列進行克隆,按照gatway 反應試劑盒說明書完成載體構建,并對載體進行測序驗證。將構建好的載體通過凍融法轉化入農桿菌EHA105 中。制備好農桿菌重懸液后將菌液注射入煙草葉片下表皮,置于全黑暗環境22℃培養3 d 后將葉片切成1.5 cm2大小,置于Nikon C2-ER 激光共聚焦顯微鏡下觀察并拍照。
2 結果與分析
2.1 HvnNPF4.5 生物信息學分析
2.1.1 HvnNPF4.5 的基因結構和啟動子功能元件預測分析測序結果表明,青稞和大麥的HvnNPF4.5基因和啟動子序列一致,HvnNPF4.5所對應的轉錄本包含2 個內含子和3 個外顯子(圖1)。HvnNPF4.5基因啟動子區域發現48 個TATA-box 和23 個CAAT-box,以及一些與干旱和脫落酸、赤霉素、生長素等植物激素相關的響應元件,其中,關于干旱、脫落酸、赤霉素和生長素的順式作用元件各1 個(表1)。
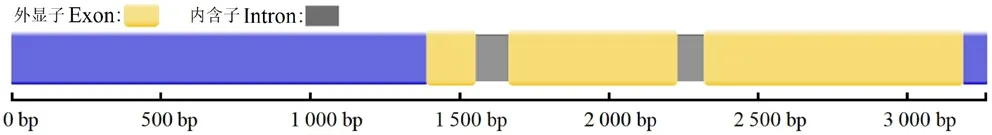
圖1 青稞HvnNPF4.5 基因結構圖
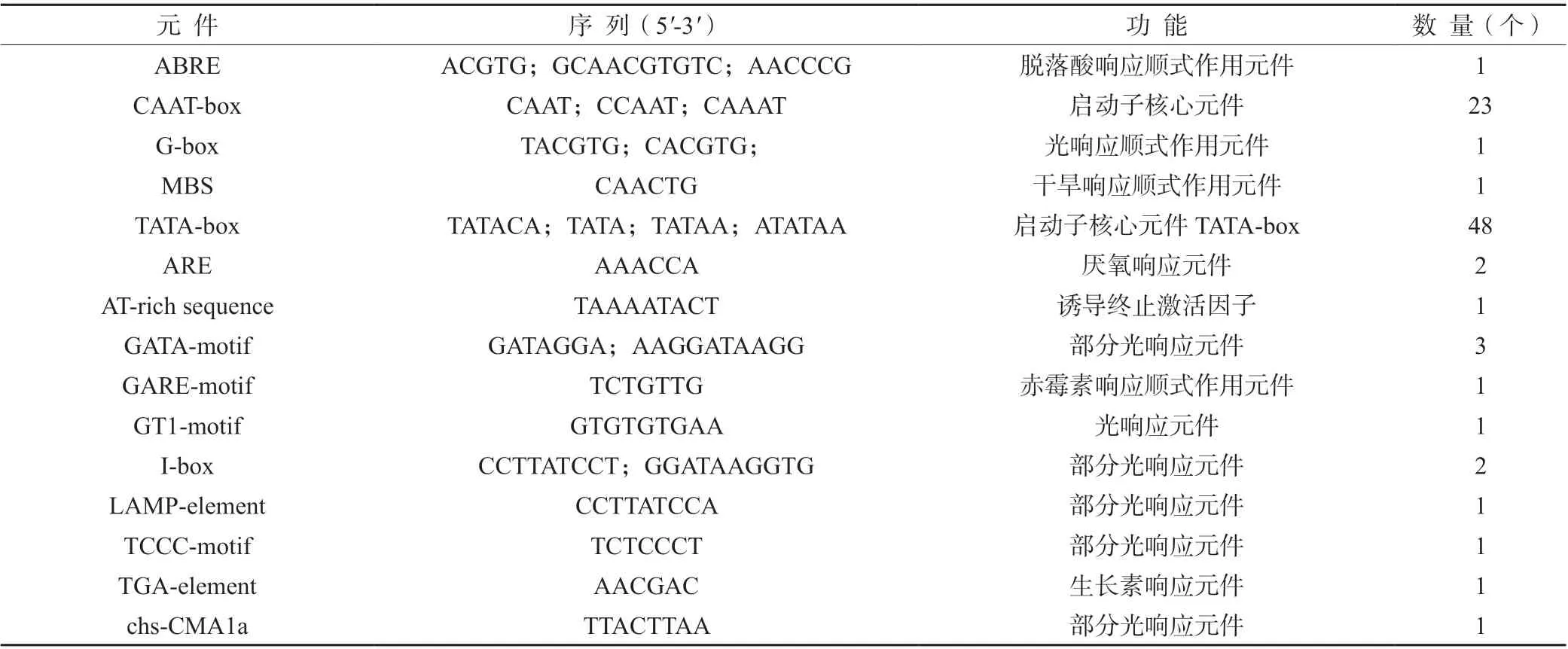
表1 青稞HvnNPF4.5 基因啟動子元件分析
2.1.2 HvnNPF4.5 蛋白的理化性質和結構分析由表2 可知,HvnNPF4.5 由600 個氨基酸組成,是疏水穩定蛋白,有12 個跨膜結構(圖2);HvnNPF4.5 蛋白序列中有5 個絲氨酸(Ser)、3 個蘇氨酸(Thr)、2 個酪氨酸(Tyr),共8 個磷酸化位點(圖3);無信號肽(圖4);HvnNPF4.5 蛋白二級結構中α 螺旋、延長鏈、β 轉角、無規則卷曲分別占48.83%、11.83%、3.5%、35.83%(圖5);對青稞HvnNPF4.5 蛋白進行同源建模生成三級結構模型,其結構和二級結構預測結果基本一致(圖6)。

表2 青稞HvnNPF4.5 蛋白質理化性質分析
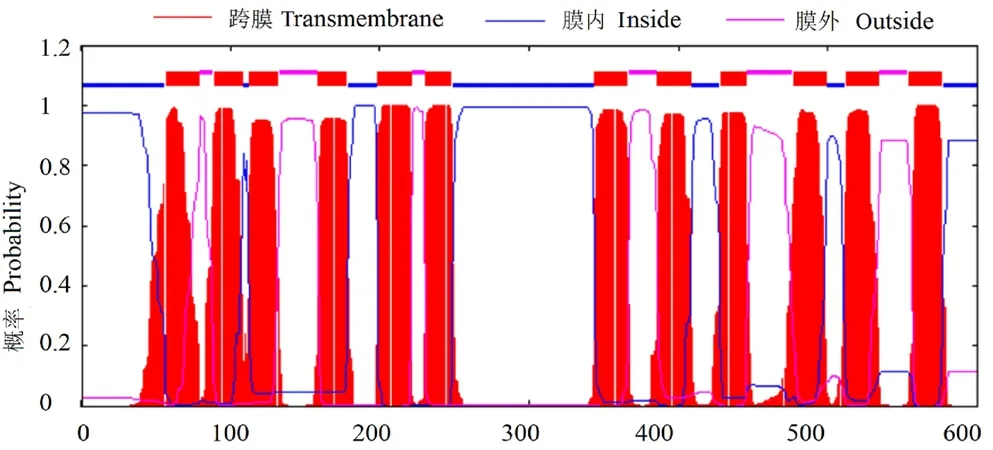
圖2 青稞HvnNPF4.5 蛋白跨膜結構預測分析

圖3 青稞HvnNPF4.5 蛋白磷酸化位點預測
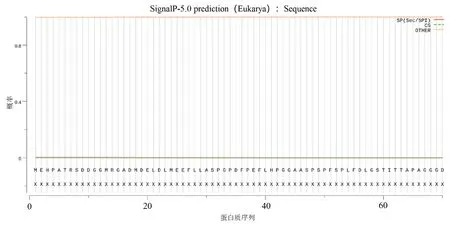
圖4 青稞HvnNPF4.5 蛋白信號肽預測
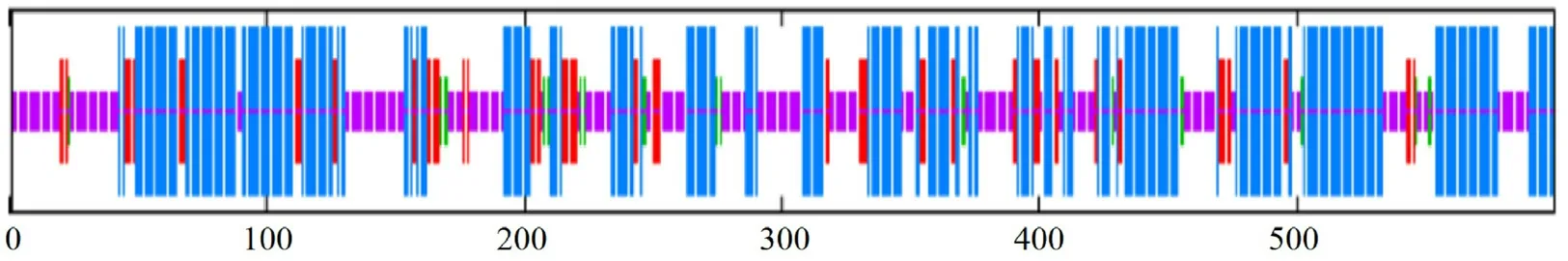
圖5 青稞HvnNPF4.5 蛋白二級結構
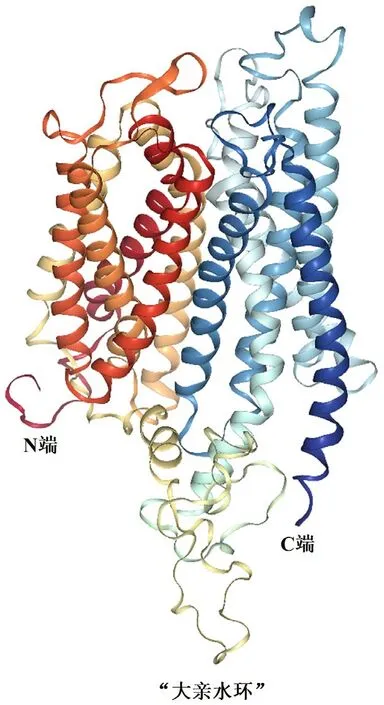
圖6 青稞HvnNPF4.5 蛋白三級結構預測分析圖
2.1.3 NPF4.5 蛋白氨基酸序列對比和同源進化分析對青稞HvnNPF4.5 蛋白進行序列比對發現,該蛋白均具有MFS(Major Facilitator Superfamily) 結構域,青稞HvnNPF4.5 與大麥HvNPF4.5 的蛋白序列完全一致(圖7)。將青稞HvnNPF4.5 和其他28 種植物的同源蛋白序列構建系統進化樹,發現青稞HvnNPF4.5 與大麥HvNPF4.5、節節麥AtsNPF4.5 的親緣關系最近(圖8)。

圖7 青稞HvnNPF4.5 蛋白氨基酸序列對比分析
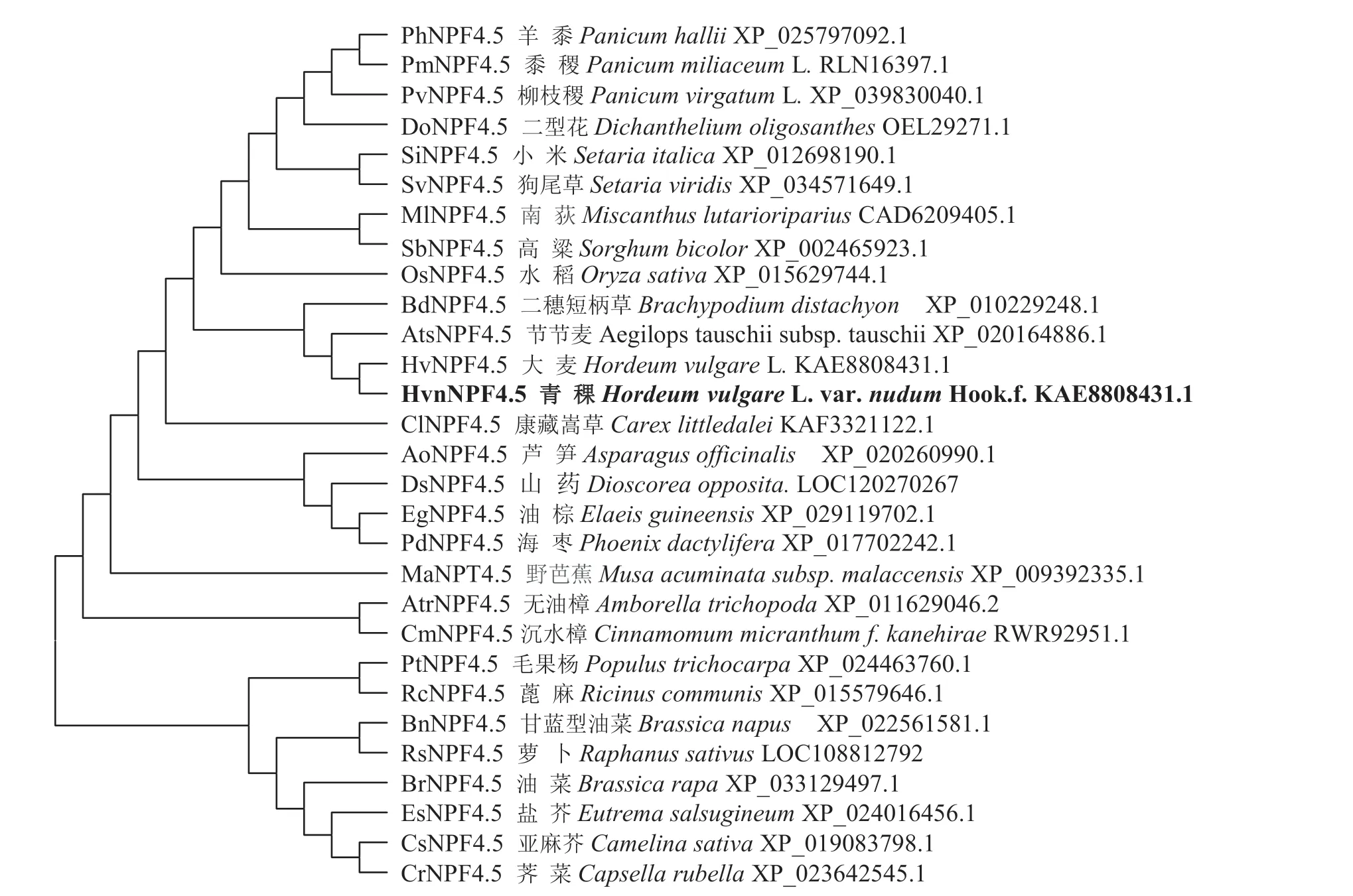
圖8 青稞HvnNPF4.5 蛋白與其他植物同源蛋白系統進化樹
2.2 青稞HvnNPF4.5 蛋白亞細胞定位結果
將注射了農桿菌的煙草葉片在顯微鏡下觀察,發現在細胞膜上檢測到紅色熒光信號,說明HvnNPF4.5蛋白定位在細胞膜上(圖9)。
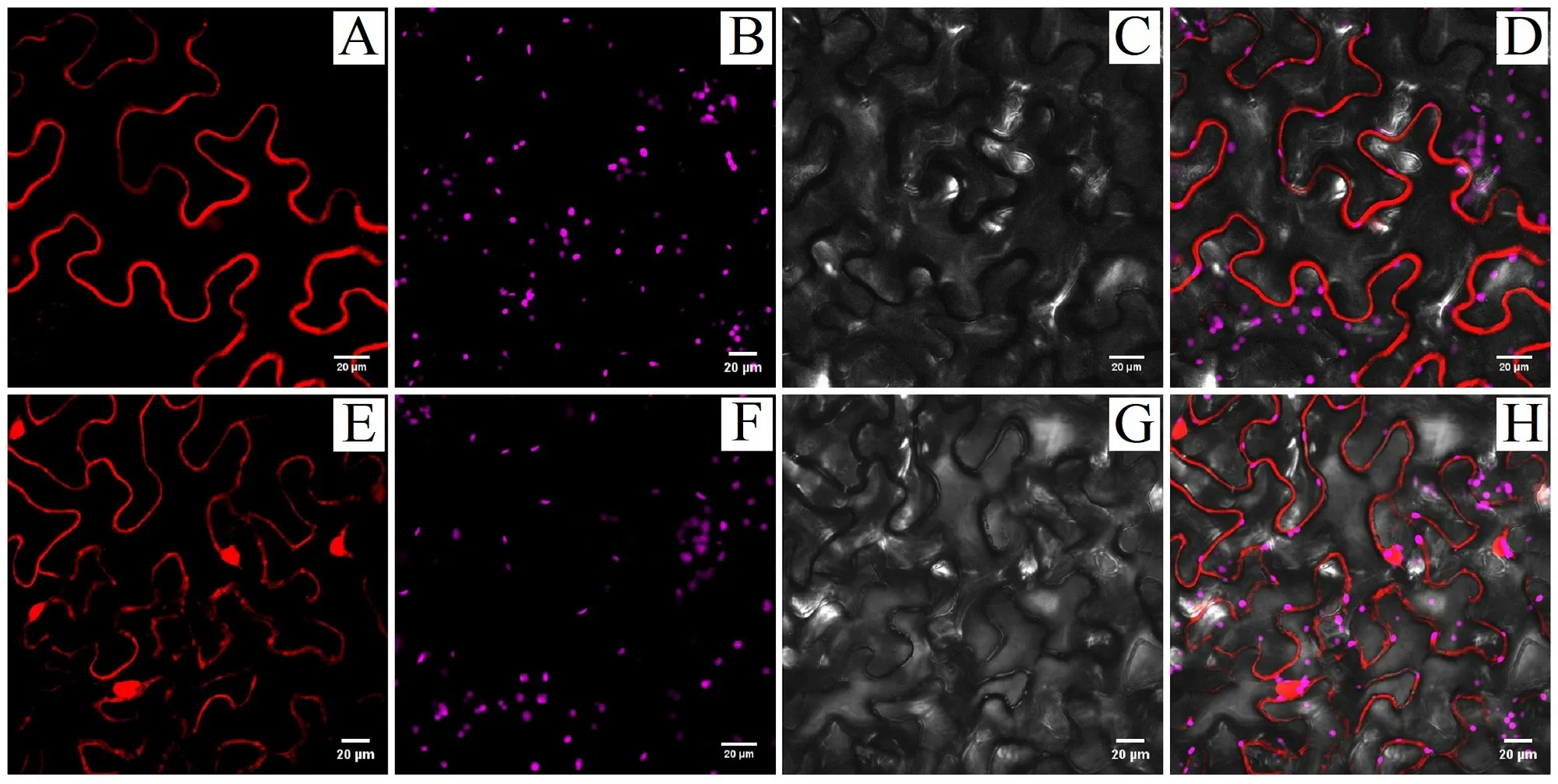
圖9 青稞 HvnNPF4.5 蛋白亞細胞定位
3 結 論
植物硝酸鹽轉運蛋白分為高親和力和低親和力2 種類型,高親和力硝酸鹽轉運蛋白主要是在正常條件下維持植物體對硝酸鹽的吸收和轉運,低親和力硝酸鹽轉運蛋白主要是在植物體受到低氮脅迫時發揮作用,為植物提供更充足的氮元素[21-23]。硝酸鹽轉運蛋白基因家族NPFs屬于低親和力硝酸鹽轉運蛋白,是植物吸收、轉運和同化硝酸鹽的主要基因家族,在植物氮元素吸收利用中發揮著重要作用[16,24-25]。
除了水稻等模式植物外,其他植物的硝酸鹽轉運蛋白基因家族NPFs研究報道也較多。袁婷婷等[26]的研究發現,毛竹中有27 個NPFs家族基因成員,分為7 個亞家族,PeNPFs啟動子序列中包含多種與低溫和干旱等非生物脅迫以及GA3和NAA 等激素響應相關的元件,PeNPFs主要定位于細胞膜和液泡中,在葉片、花序、根、鞭和筍中均有表達,并且受多種非生物脅迫和植物激素的誘導表達,PeNPFs的不同成員在不同組織以及應對不同環境過程中發揮著多樣化的功能。王倩[27]的研究發現,73 個蘋果MdNPFs成員可分為8 個亞族,且分布在17 條染色體上,有12 個跨膜結構域,在根、莖、葉、花和果實中均有表達,其表達受硝酸鹽濃度的影響,將其中的MdNPF6.5基因轉入蘋果愈傷組織中發現轉基因愈傷組織在低氮下鮮重明顯高于對照,表明MdNPF6.5在提高蘋果耐低氮能力方面具有重要作用。計麗[24]的研究發現,大多數木薯MeNPFs基因含有2~5 個內含子,少數只含有1 個內含子,72 個MeNPFs基因不均勻分布于18 條染色體上;進一步研究發現MeNPF6.2和MeNPF6.3有12 個跨膜結構,均定位于細胞膜上,MeNPF6.2和MeNPF6.3分別在木薯葉片和幼苗根中受硝酸鹽誘導表達;將MeNPF6.2基因轉入木薯和水稻中發現,轉基因木薯植株葉面積更大、莖桿更加粗壯、側根數量也明顯增多,轉基因水稻則表現為百粒重和粒寬顯著增加,表明MeNPF6.2基因在木薯響應低氮脅迫時起重要作用,在作物耐低氮改良過程中應用前景廣闊。馬嘉俊等[14]對白菜BrNPFs進行研究時發現,白菜BrNPFs基因家族成員分為8 個亞族,不均勻地分布在10 條染色體上,共有72 個成員,亞細胞定位預測結果顯示BrNPFs 均定位在質膜上。筆者的研究結果與以上研究報道基本相符。HvnNPF4.5包含2 個內含子,啟動子區有與干旱以及脫落酸、赤霉素、生長素相關的順式活性作用元件;HvnNPF4.5 蛋白的分子量為66 875.31 Da,理論等電點為8.85,具有8 個磷酸化位點,無信號肽結構,也具有12 個跨膜結構,在第6~7 跨膜域間同樣存在一個大親水環;HvnNPF4.5和其他物種的同源蛋白都有MFS 結構域,青稞HvnNPF4.5 與大麥HvNPF4.5、節節麥AtsNPF4.5 的親緣關系最近;亞細胞定位結果顯示青稞HvnNPF4.5定位于細胞膜上。
青稞是青藏高原地區最具特色的作物,減少青稞生產中氮肥的施用量,提高青稞對氮肥的利用效率對于青稞種植業可持續發展具有重要意義。該研究克隆得到了青稞硝酸鹽轉運蛋白基因HvnNPF4.5,對其基因和蛋白進行了生物信息學分析,并對其進行亞細胞定位研究,其結果為青稞氮元素吸收、轉運和同化相關基因研究提供了一定的參考。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
文苑(2020年6期)2020-06-22 08:41:52
科技傳播(2019年22期)2020-01-14 03:06:54
文苑(2019年22期)2019-12-07 05:29:00
