厚殼貽貝新型抗菌肽myticalin和mytimacin的分子鑒定
2022-12-21 07:24:46楊金月陽宗欣何建瑜張曉林何夢嵐嚴小軍
水生生物學報 2022年12期
關鍵詞:結構
劉 璐 楊金月 陽宗欣 何建瑜 張曉林 何夢嵐 嚴小軍 廖 智
(浙江海洋大學海洋科學與技術學院, 海洋生物資源與分子工程研究室, 舟山 316022)
貽貝是我國東部海域重要的養殖貝類之一, 具有重要的經濟價值。目前, 在牡蠣, 蛤類等代表性貝類的養殖過程中已顯現出大規模病害的威脅, 并因此對貝類養殖產業帶來嚴重影響[1—4]。但值得注意的是, 與其他貝類相比, 貽貝在養殖過程中表現出對疾病的較強耐受性[4—6], 目前尚無貽貝屬(Mytilus)大規模病害的報道[4,5]。這表明, 與其他貝類相比, 貽貝的免疫系統可能具有其獨特性, 因此,貽貝已成為海洋生物免疫相關研究的重要對象[6—10]。以往的研究表明, 貽貝的免疫系統組成較為復雜,包括黏液免疫系統、細胞免疫系統及體液免疫系統[11], 涉及免疫識別、免疫信號傳遞和免疫效應發揮等不同階段。 涉及的具體免疫分子機制包括免疫細胞聚集、免疫細胞吞噬等細胞免疫過程, 以及酚氧化酶、氮自由基、裂解酶, 抗菌肽等分子免疫機制[7,11—13]。其中, 貽貝抗菌肽在貽貝的免疫系統中負責對入侵微生物的直接殺傷和抑制, 且其結構和功能具有較強的分子多樣性。目前已先后有10種不同類型的貽貝抗菌肽被報道[7]。其中, mytilin、myticin、mytimycin和Mytilus galloprovincalisdefensin (MGD)是最早從地中海貽貝(Mytilus galloprovincalis)血清中被分離得到[14—17]; 之后, myticusin和mytichitin從厚殼貽貝血清中分離得到[18,19]; 此外, 又陸續從地中海貽貝組織轉錄組中預測到big defensin、mytimacin、myticalin和arthropod like defensin (ALD)四種貽貝抗菌肽序列[7,20,21]。從已有研究結果來看, 貽貝抗菌肽不僅家族數眾多, 且在同一家族內也表現出極強的分子多樣性[22,23]。豐富的貽貝抗菌肽分子類型為新型生物抗生素的研發提供了重要的先導分子來源。
厚殼貽貝(Mytilus coruscus)是我國東部海域最重要的養殖貝類之一, 也是西太平洋沿岸的代表性貽貝物種。此前已鑒定厚殼貽貝抗菌肽mytilin[24]、myticin[25]、myticusin[18,26]及mytichitin[27]。但與其他貽貝屬物種, 如地中海貽貝(M.galloprovincalis)和紫貽貝(M.edulis)相比, 厚殼貽貝中抗菌肽研究仍存在很大空間。為進一步挖掘厚殼貽貝新型抗菌肽分子, 通過對厚殼貽貝基因組[28]及血細胞轉錄組[29]進行數據篩選, 從中獲得兩種新型的厚殼貽貝抗菌肽分子。序列分析結果表明, 該新型抗菌肽與地中海貽貝已報道的myticalin[20]和mytimacin[21,30]具有較高的序列相似性。myticalin是一種首次從貽貝中發現的, 其成熟肽序列中不含二硫鍵的抗菌肽, 對革蘭氏陽性和革蘭氏陰性菌均具有抑制作用[20]。而mytimacin則是一種富含半胱氨酸并形成多對二硫鍵的抗菌肽家族, 從結構上看, 屬于macin抗菌肽家族。macin家族抗菌肽首次發現于環節動物Theromyzon tessulatum, 具有多對二硫鍵以及對革蘭氏陽性菌較強的抑制活性[31,32]。此外, 軟體動物, 如貝和螺類中也均發現macin抗菌肽的存在[33,34], 表明macin是一種低等生物中較為保守的抗菌肽。myticalin和mytimacin抗菌肽在厚殼貽貝中的發現, 豐富和拓展了對貽貝抗菌肽分子多樣性的認識, 但這兩種抗菌肽的分子特征及其對厚殼貽貝免疫防御的貢獻目前尚不清楚。為此, 對厚殼貽貝myticalin和mytimacin開展了序列分析, 固相化學合成及功能驗證, 以及在不同微生物誘導后, myticalin和mytimacin的免疫響應研究。結果表明, 厚殼貽貝myticalin和mytimacin分別與地中海貽貝的myticalin和mytimacin具有較強的序列相似性, 結構預測結果表明上述兩種抗菌肽具有不同的空間結構; 化學合成的myticalin和mytimacin具有廣譜的抑菌活性。此外, myticalin和mytimacin在應對不同微生物誘導時具有不同的免疫響應特征。上述研究一方面為深入了解厚殼貽貝的抗菌肽分子多樣性及myticalin和mytimacin在貽貝免疫過程中的分子角色奠定了基礎, 也為后續開發厚殼貽貝新型抗菌肽為來源的生物抗生素提供了科學依據。
1 材料與方法
1.1 厚殼貽貝血細胞轉錄組數據挖掘與序列分析
根據厚殼貽貝血細胞轉錄組所測63942條unigene[29], 根據其序列的注釋結果, 利用BlastX程序從中篩選myticalin和mytimacin基因序列, 閾值(Evalue)為1E-05。將篩選到的基因序列對比厚殼貽貝基因組數據庫以驗證序列的完整性, 選擇開放閱讀框完整的myticalin和mytimacin基因序列代表性序列開展后續分析。其中, 開放閱讀框采用Lasergene軟件的Editseq模塊(版本7.1.0)進行分析。氨基酸序列中的成熟肽、結構域和信號肽預測分別采用ProP軟件[35]、SMART軟件 (http://smart.emblheidelberg.de/)及SignalP 4.0軟件[36]進行分析。蛋白質二級結構和三級結構分別利用Prabi (http://pbil.ibcp.fr/)及SWISS MODEL服務器在線進行預測。多序列比對采用CLUSTALO軟件 (https://www.ebi.ac.uk/Tools/msa/clustalo/)在線進行。
1.2 固相多肽化學合成及鑒定
多肽固相化學合成參照文獻[37], 在十二通道半自動多肽合成儀(上海強耀)上合成, 合成方向為從羧基端向氨基端進行。合成后的線性多肽粗品以高效液相色譜儀(Agilent 1260, 美國安捷倫公司)進行分離純化, 采用C18分析型反相柱 (Kromasil 100-5, 4.6 mm×250 mm, 5 μm)進行分離, 洗脫液分別為A液 (含0.1%TFA的純水)和B液 (含0.1%TFA的乙腈); 其中, myticalin合成產物的洗脫梯度為20min內B液比例由20%上升到80%; mytimacin合成產物的洗脫梯度為25min內B液比例由15%上升到40%; 流速均為1.0 mL/min; 采用紫外檢測器進行檢測, 檢測波長為 280 nm。收集洗脫目標峰開展質譜分析, 采用質譜 (Agilent-6125B, 美國安捷倫公司)對合成后的多肽純品進行精確分子量鑒定, 質譜檢測條件參照文獻[37]進行。參照文獻[37]方法對合成后的多肽樣品開展基于谷胱甘肽的氧化復性。復性后的多肽進一步經高效液相色譜純化后開展功能分析。
1.3 抑菌活性測試
參照文獻[38]方法, 采用生長曲線抑制法對復性后的myticalin和mytimacin開展抑菌活性測試。myticalin和mytimacin以生理鹽水分別配置成200、100、50、25、12.5、6.25、3.13、和1.56 μmol/L濃度梯度。測試菌種購自北京中國普通微生物菌種保藏管理中心, 包括5種革蘭氏陰性菌, 分別為大腸桿菌Escherichia coli、銅綠假單胞菌Pseudomonas aeruginosa(ATCC9027)、哈維氏弧菌Vibrio harveyi(ATCC33842)、溶藻弧菌Vibrio alginolytica(ATCC19659)和副溶血弧菌Vibrio Parahaemolyticus(ATCC17802); 4種革蘭氏陽性菌, 分別為巨大芽孢桿菌Bacillus megaterium(ATCC19161)、金黃色葡萄球菌Staphylococcus aureus(ATCC25923)、枯草芽孢桿菌Bacillus subtilis(ATCC19659)和藤黃疊球菌Sarcina lutea(ATCC4698); 真菌為白色念珠菌Candida albicans(ATCC10231)。
1.4 紅細胞毒性分析
取綿羊血 (購自南京森貝伽生物科技有限公司), 離心 (1000×g, 10min, 4℃)收集紅細胞, 以PBS緩沖液 (pH 7.4)洗滌3次, 之后以PBS緩沖液(pH 7.4)重懸細胞制備成1% 紅細胞懸浮液。合成的myticalin和mytimacin以PBS緩沖液 (pH 7.4)配置成1 mmol/L濃度, 按照1﹕9比例分別加入抗菌肽溶液和紅細胞懸浮液, 抗菌肽終濃度分別為100、50、25、12.50、6.25、3.13、1.56和0.78 μmol/L濃度梯度, 以 PBS緩沖液(含0 μmol/L抗菌肽)為陰性對照, 以 triton X-100為陽性對照。37℃培養4h后取出, 經離心 (1000×g, 10min, 4℃)后, 上清液轉移至96孔板, 以酶標儀于波長405 nm處測定吸光度。抗菌肽溶血率按照 (檢測孔A405–陰性孔A405)/(陽性孔A405–陰性孔A405)×100%進行計算。
1.5 掃描電鏡觀察
以溶藻弧菌、金黃葡萄球菌和白色念珠菌作為指示菌, 分別加入myticalin和mytimacin (100 μmol/L); 孵育8h后, 離心收集菌體, 經4%多聚甲醛固定后, 參照文獻[37]方法對其開展掃描電鏡(Nova nano SEM 450, 美國FEI)觀察。
1.6 熒光定量PCR分析
根據厚殼貽貝myticalin和mytimacin序列中開放閱讀框序列設計特異性引物 (表1), 參照文獻[37]方法進行PCR擴增及測序分析。進一步設計熒光定量PCR引物 (表1)并開展myticalin和mytimacin的基因表達分析。細菌誘導實驗參照文獻[39]進行。誘導微生物包括金黃色葡萄球菌、溶藻弧菌和白色念珠菌, 誘導后分別于0.5h、1h、2h、 4h、8h、12h、24h、48h 和72h, 采集厚殼貽貝血淋巴, 血淋巴經離心 (1500×g, 10min, 4℃)后獲得血細胞。對血細胞進行總RNA提取和逆轉錄, 所得血細胞cDNA作為模板開展后續熒光定量PCR分析。
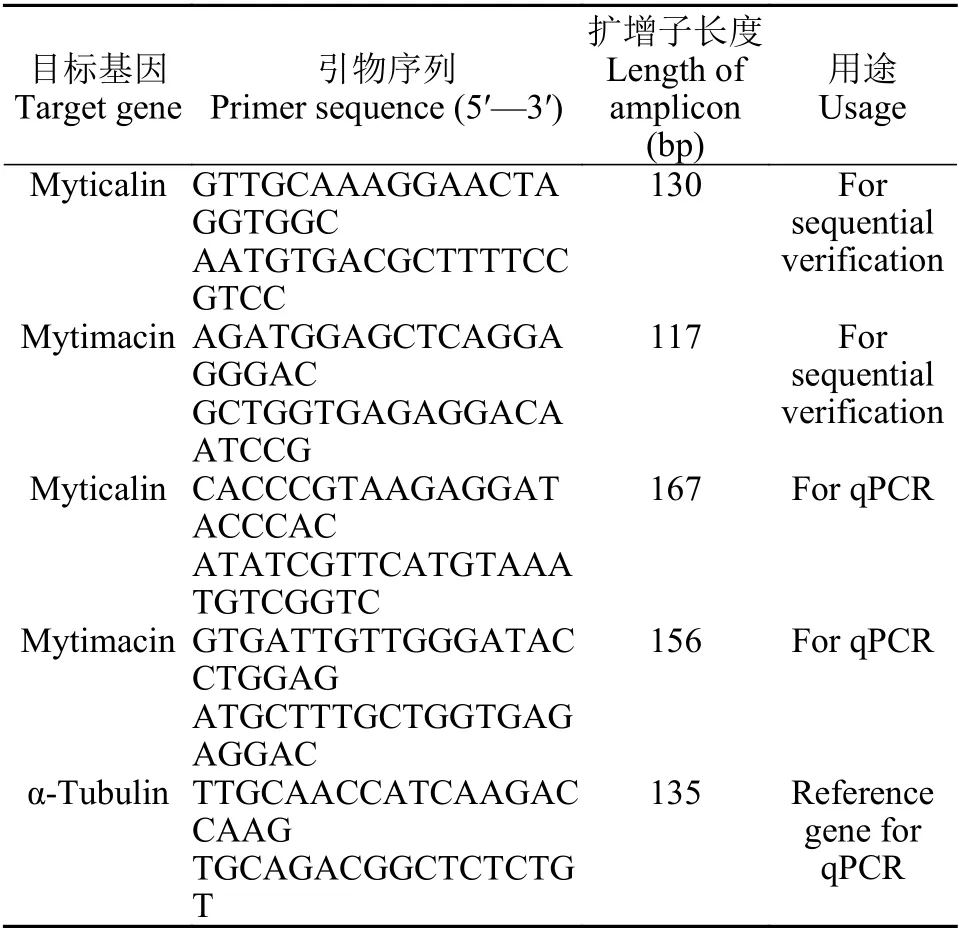
表1 熒光定量PCR研究的特異性引物Tab.1 Primers for qPCR analysis
熒光定量PCR參照文獻[37]方法在實時定量PCR系統 (MX3000P, 美國Stratagene公司)進行。表達量依據Ct值采用2–??Ct方法[40]進行分析。采用3次重復試驗, 所得數據以SPSS 軟件(v25.0)的Oneway ANOVA方法進行顯著性差異分析,P<0.05代表具有顯著性差異。
2 結果
2.1 厚殼貽貝myticalin和mytimacin的序列特征
兩種新型厚殼貽貝抗菌肽序列已上傳, 其在GenBank數據庫中的編號分別為MW922042.1/QXT 26521.1 (myticalin)和MW922041.1/QXT26520.1(mytimacin)。myticalin基因開放閱讀框長度為336 bp,編碼一條113個氨基酸殘基組成的前體多肽, 包括23個氨基酸殘基組成的信號肽、37個氨基酸殘基組成的成熟肽和53個氨基酸殘基組成的pro區。myticalin成熟肽理論分子量為4459.25 Da, 理論等電點為12.30, 屬于堿性多肽; 其成熟肽序列中含量最豐富的氨基酸為脯氨酸, 其含量達到28.6%, 此外, 精氨酸含量也較高, 達到19.1%。mytimacin基因開放閱讀框長度為261 bp, 編碼的前體肽長度為86個氨基酸殘基, 包括22個氨基酸殘基組成的信號肽, 17個氨基酸殘基組成的pro區和47個氨基酸殘基組成的成熟肽。其成熟肽序列中含有6個半胱氨酸, 推測形成3對二硫鍵。此外, mytimacin成熟肽序列中甘氨酸和絲氨酸含量較豐富, 占比均為12.8%; 其理論分子量為5232.08 Da, 理論等電點為9.20, 屬堿性多肽。
二級結構預測結果表明, myticalin前體肽序列的二級結構以無規卷曲為主, 占比達到88.57%。其前體肽序列中, 除信號肽之外無其他可預測的結構域, 但是存在低復雜度區域(Low complexity region),其位置分別在25—54號殘基和64—76號殘基, 其特征在于富含脯氨酸和天冬氨酸。在mytimacin序列中, Alpha螺旋區含量占比10.64%, 完全伸展區占比19.15%, 無規卷曲占比53.19%。其前體肽序列中含有一個典型的macin結構域 (結構域編號: PF14865),其位置位于前體肽的29—85號殘基。Macin結構域是一種由二硫鍵維持其穩定的致密結構域, 含macin結構域的蛋白通常均具有抑菌活性, 如theromain、neuromacin和hydramacin-1等[33,34,41]。在結構上, macin結構域的典型特征在于形成一個二硫鍵穩定的α/β超二級結構模體[41]。
三級結構預測結果表明, myticalin在其成熟肽序列中無半胱氨酸及由此形成的二硫鍵, 且序列中存在豐富的脯氨酸, 因而整體結構呈現無規卷曲狀態, 但不排除局部肽段形成α-螺旋。由于myticalin在三級結構預測中無合適的模板結構, myticalin的三級結構采用從頭預測 (De novo)手段進行預測, 因此, 其結果與二級結構預測結果并不完全一致; mytimacin的三級結構呈現典型的二硫鍵穩定α/β結構[42]。 其三級結構特征包括均包含一段位于序列N端的2段反平行β-折疊和C端的α-螺旋, 且三級結構預測結果與二級結構預測結果具有類似特征。
對myticalin和mytimacin的前體肽序列的同源搜索比對結果表明, myticalin與來自貽貝屬的myticalin具有較高序列相似性, 其序列一致性達到70%以上, 其保守區域主要集中在信號肽及pro區, 而成熟肽序列差異較大。而mytimacin則與來自貽貝屬、牡蠣屬及蛤類的mytimacin具有較高序列相似性, 特別是在序列中段, 即macin結構域所在區域具有較高的保守性。
2.2 myticalin和mytimacin的固相化學合成
為檢測myticalin和mytimacin的抑菌活性, 采用固相多肽合成策略, 完成對這兩種抗菌肽的成熟肽區序列的化學合成。由圖1可知, mytimacin合成后的粗品經高效液相色譜純化, 其目標峰出峰時間為9.28min, 根據曲線下面積計算, 其純度達到95%以上; 質譜鑒定結果表明, 合成的mytimacin分子量為5232.07 Da, 與理論分子量(5232.08 Da)一致 (圖1A);合成的myticalin經高效液相色譜純化, 其純度達到96%以上, 其質譜鑒定分子量為4459.40 Da, 與理論分子量(4459.25 Da)一致 (圖1B)。

圖1 固相化學合成myticalin (A)和mytimacin (B)粗品的HPLC純化與分子量質譜鑒定Fig.1 Purification of solid chemical synthesized myticalin (A)and mytimacin (B)by HPLC and the mass spectrum of the fraction collected from HPLC elution
2.3 化學合成的myticalin和mytimacin的抑菌活性
厚殼貽貝myticalin和mytimacin分子經化學合成后進行氧化復性, 進一步通過高效液相色譜法對復性后的多肽進行純化。由圖2可知, 復性后的多肽經C8反相柱分離后, 純度較高, 可用于后續抑菌活性實驗。myticalin和mytimacin的抑菌活性結果見圖3, 合成的myticalin對不同微生物的抑菌活性存在差異, 其最低抑菌濃度(Minimal inhibitory concentration, MIC)在1.56—200 μmol/L。其中, myticalin對巨大芽孢桿菌和藤黃疊球菌抑制活性較強, 其MIC值低于8 μmol/L; 而對革蘭氏陽性菌的抑制活性普遍較弱, 其最低抑菌濃度均在50 μmol/L及以上;而對白色念珠菌則只有微弱抑制活性, 其MIC值達到200 μmol/L (圖3A)。合成的mytimacin對所測試菌種均表現出明顯的抑制活性, 但整體而言, 其對革蘭氏陽性菌的抑制活性要弱于革蘭氏陰性菌。其對革蘭氏陽性菌的MIC值在100 μmol/L, 而對革蘭氏陰性菌的MIC值多為50 μmol/L, 但對銅綠假單胞菌抑制活性較弱, 其MIC值達到200 μmol/L, 此外, mytimacin對白色念珠菌的MIC值為50 μmol/L,相比myticalin表現出對真菌較強的抑制活性 (圖3B)。

圖2 復性后的myticalin和mytimacin的反相液相色譜純化圖Fig.2 The purification of myticalin and mytimacin by reverse-phase HPLC after oxidation after oxidation

圖3 化學合成的myticalin (A)和mytimacin (B)的抑菌活性Fig.3 The antimicrobial activities of chemical synthesized and refolded myticalin (A)and mytimacin (B)
2.4 紅細胞毒性分析及掃描電鏡觀察結果
抗菌肽溶液與綿羊紅細胞懸浮液經孵育,OD405檢測結果表明, 與對照組相比, 不同濃度的myticalin和mytimacin均未能導致紅細胞發生明顯溶血現象, 不同濃度的myticalin和mytimacin其溶血率均小于5%, 與對照組無明顯差異(P>0.05; 圖4)。

圖4 myticalin和mytimacin對綿羊紅細胞的溶血活性測試 (平均值±標準差, n=3)Fig.4 Erythrotoxicity activity of myticalin and mytimacin against sheep red blood (mean±SD, n=3)
進一步利用掃描電鏡觀察myticalin與mytimacin對三種不同微生物形態學影響, 結果表明, 與對照組相比, 兩種抗菌肽均對金黃色葡萄球菌產生明顯影響, 導致金黃葡萄球菌產生凝集現象(圖5A—C),且凝集后的菌體出現裂解和內容物釋放現象。此外, myticalin與mytimacin對溶藻弧菌的表面形態結構也具有明顯影響, 如圖5D—F所示, 與對照組相比, 兩種抗菌肽均導致溶藻弧菌表面結構變得粗糙, 出現皺縮或片狀凸起, 但未導致細菌內容物的釋放。此外, 與對照組相比, mytimacin導致白色念珠菌表面出現破壁現象, 導致出現火山口狀孔洞(圖5G和H)。而myticalin則對白色念珠菌的形態結構無明顯影響 (圖5I)。

圖5 掃描電鏡觀察myticalin與mytimacin對金黃葡萄球菌、溶藻弧菌和白色念珠菌的形態結構影響Fig.5 Scanning electron microscopic observation of the effects of myticalin and mytimacin on the morphology and structure of S.aureus,V.alginolyticus, and C.albicans, respectively
2.5 myticalin和mytimacin在不同微生物刺激下的免疫反應模式
為進一步分析myticalin和mytimacin在不同微生物刺激下其在厚殼貽貝血細胞中的相對表達量變化。分別采用金黃葡萄球菌、溶藻弧菌及白色念珠菌對厚殼貽貝進行注射誘導, 熒光定量PCR結果見圖6。由圖6A可見, 在血細胞中, myticalin對金黃葡萄球菌誘導具有較強敏感性, 與對照組相比其相對表達量在1—12h均出現顯著上調 (P<0.05),其峰值出現在誘導后4h, 且其相對表達量相對于對照組上升了約7倍; 而在溶藻弧菌誘導下, 與對照組相比myticalin相對表達量僅在2h和4h出現顯著上調 (P<0.05), 其最大上升幅度約3.5倍; 此外, 在白色念珠菌誘導下, myticalin的相對表達量與對照組相比僅在4h和12h出現顯著上調 (P<0.05), 最大上升幅度約3.5倍 (圖6A)。
Mytimacin則對白色念珠菌和金黃葡萄球菌誘導表現出較強敏感性。在白色念珠菌及金黃葡萄球菌誘導下, mytimacin的相對表達量與對照組相比在2—8h出現顯著上調(P<0.05), 其最大上升幅度分別為3倍和2.8倍; 在溶藻弧菌誘導下, mytimacin相對表達量與對照組相比僅在4h和8h出現表達量的顯著上調(P<0.05), 且其最大上升幅度約為2倍(圖6B)。
3 討論
3.1 myticalin和mytimacin的序列特征
貽貝體內豐富的抗菌肽家族已成為新型生物抗生素研發的重要先導分子來源[43,44]。隨著厚殼貽貝基因組序列及其多種組織轉錄組數據的公布,為預測和篩選厚殼貽貝新型抗菌肽分子奠定了基礎。但厚殼貽貝中多數預測的抗菌肽分子因其在體內含量極低, 因此, 常規的蛋白質分離純化技術不足以獲得足夠樣品而進行后續分析。常規的蛋白質重組表達策略因抗菌肽序列中豐富的半胱氨酸及其形成的二硫鍵, 因而成功的例子也不多。目前厚殼貽貝中僅myticusin和mytichitin兩種抗菌肽的真核重組表達獲得了成功[26,45]。
Myticalin[20]和mytimacin[32]最早從地中海貽貝基因組中預測得到。通過對厚殼貽貝基因組及轉錄組學數據的篩選, 我們發現厚殼貽貝中也存在上述兩種抗菌肽的序列。但是對這兩種抗菌肽分子特性及其在貽貝免疫過程中的作用和機制尚不清楚, 為此, 我們對厚殼貽貝myticalin和mytimacin開展了固相化學合成, 抑菌活性測試以及不同微生物誘導后的表達譜開展研究。由于兩種抗菌肽在貽貝體內含量極低, 因此無法通過常規的蛋白質分離純化手段獲得其樣品, 因此, 對其序列分析主要依賴于生物信息學手段。分析結果表明, 兩種抗菌肽前體序列中均含有pro區, 該區域被認為是貽貝抗菌肽普遍具有的結構特征之一[46]。但是該區域在兩種抗菌肽序列中的位置不同, Myticalin的pro區位于其預測成熟肽區的C端, 而在mytimacin中則位于成熟肽與信號肽之間, pro區在部分生物毒素多肽中被認為是蛋白翻譯后修飾的特征之一, 該區肽段可在翻譯后被羧肽酶E切除[47,48]。目前對pro區的生物學意義尚不清楚, 有研究發現富含酸性氨基酸的per-pro區可能有助于富含堿性氨基酸的成熟肽在細胞內的穩定[49], 三級結構預測結果表明 Myticalin和mytimacin的pro區也符合該特征, 其序列中酸性氨基酸數量較多, 特別是myticalin的pro區中, 其酸性氨基酸(天冬氨酸和谷氨酸)占比超過35%, 而其成熟肽中其堿性氨基酸(精氨酸)占比達到19%,因此, 其pro區高含量的酸性氨基酸可能有助于維持多肽在細胞內的穩定。
3.2 myticalin和mytimacin的結構特征與其抗菌活性之間的關聯
從厚殼貽貝myticalin和mytimacin的結構來看,兩種抗菌肽表現出不一樣的結構特征。其中, 最大的區別在于myticalin成熟肽序列中不含半胱氨酸,這也是目前發現的貽貝抗菌肽中唯一不含有半胱氨酸的抗菌肽家族。此外, myticalin的成熟肽序列中富含脯氨酸和精氨酸, 該特征類似于哺乳動物中的經典抗菌肽cathelicidin[50]。其中, 脯氨酸已被證明對抗菌肽活性發揮具有重要作用, 特別是可通過非裂解方式破壞細菌細胞膜從而發揮抑菌活性[51]。三級結構預測結果表明, 缺乏二硫鍵的myticalin主要形成無規卷曲結構, 但在部分肽段出現螺旋結構。雖然脯氨酸對規則二級結構具有破壞作用, 但連續的脯氨酸也可能會形成螺旋結構, 且該結構與蛋白質聚集體形成及位點識別具有關聯[52]。事實上, 富含脯氨酸的抗菌肽已在多種物種中被發現,包括哺乳動物、節肢動物及植物[53]。此外也有研究發現, 精氨酸同樣對這類抗菌肽的抑菌活性具有重要貢獻[54,55]。因此, 我們推測, myticalin成熟肽序列中高含量的脯氨酸和精氨酸可能是其具有抑菌活性的內在原因。此外, 以往的研究表明, 富含脯氨酸的抗菌肽通常對真核細胞毒性較低[56,57], 而我們的研究結果同樣表明, 合成后的myticalin對真菌抑制效果不明顯(圖3), 同時對綿羊紅細胞也未表現出明顯毒性(圖4)。與myticalin相比, mytimacin則表現出貽貝抗菌肽的普遍特征, 首先具有多對二硫鍵, 其三級結構表現為典型的二硫鍵穩定的α/β結構。此外, mytimacin前體序列中pro區位于信號肽與成熟肽之間, 也符合多數軟體動物與節肢動物抗菌肽的特征[16,58]。
為驗證myticalin和mytimacin的體外抑菌活性,采取固相化學合成策略結合多肽復性手段, 獲得myticalin和mytimacin的化學合成樣品。體外抑菌試驗結果表明, 兩種抗菌肽均表現出抑菌活性。但是合成的mytimacin的抑菌活性要強于myticalin, 這可能與mytimacin具有較多規則二級結構有關。此外, myticalin表現出對革蘭氏陰性菌較強的抑制作用, 這一點此前其他富含脯氨酸的抗菌肽類似[20,53]。而mytimacin則對革蘭氏陽性菌抑制作用較強, 這與此前對地中海貽貝mytimacin的研究結果類似[31]。從掃描電鏡結果來看, 兩種抗菌肽對金黃葡萄球菌和溶藻弧菌的形態學影響類似, 意味著兩種抗菌肽可能具有相似的作用機制。此外, mytimacin對真菌也具有明顯抑制作用, 而myticalin則對真菌的抑制作用較弱.掃描電鏡結果也證實了這一結果, mytimacin能顯著改變白色念珠菌的形態結構, 而myticalin則無此影響。上述研究結果表明myticalin和mytimacin對各種微生物的抑制作用差異, 可能與其在貽貝體內可能具有不同的分工有關。兩種抗菌肽在體外實驗中均無明顯的溶血活性, 表明兩種抗菌肽均具有開發成為抗生素的潛力。
3.3 myticalin和mytimacin的免疫響應特征
考慮到血細胞在貽貝免疫過程中的重要作用[6],我們選擇了血細胞研究myticalin和mytimacin在微生物刺激條件下的表達量變化。同時, 考慮到貽貝對不同微生物具有不同的免疫特征[59], 我們分別選取了金黃葡萄球菌、溶藻弧菌及白色念珠菌作為刺激條件。從研究結果來看, myticalin和mytimacin均表現為微生物誘導后產生顯著的免疫響應(圖6),表明兩種抗菌肽分子均參與了厚殼貽貝的免疫過程, 但兩種抗菌肽對不同微生物的反應敏感性具有差異。Myticalin表現出對金黃葡萄球菌的較強敏感性, 其響應持續時間及表達量上調倍數均強于溶藻弧菌和白色念珠菌誘導。而mytimacin則表現出對白色念珠菌的反應敏感性, 其次是對金黃葡萄球菌, 而對溶藻弧菌反應較弱。上述結果表明, myticalin和mytimacin在貽貝不同微生物刺激下具有差異化的免疫響應模式。該模式推測可能與兩種抗菌肽具有不同的免疫識別和免疫信號傳導機制有關。不同類型抗菌肽具有不同的免疫響應特征在節肢動物抗菌肽研究中已有較為深入的了解。例如, 已發現具抗真菌活性的節肢動物防御素的表達受到Toll 信號通路的調控, 而其他防御素的表達則受到免疫缺陷(Immune Deficiency, IMD)信號通路的調控[60]。貽貝中目前發現多種與免疫相關的信號通路[61,62], 但其與貽貝抗菌肽表達之間的關聯目前尚未明確。我們推測, myticalin和mytimacin在貽貝體內參與免疫防御過程可能受到不同的免疫信號通路調節, 但該推測還需進一步的實驗證實。

圖6 三種不同微生物刺激下厚殼貽貝myticalin和mytimacin在血細胞中的表達量變化時間曲線 (平均值±標準差, n=3)Fig.6 The relative expression of myticalin and mytimacin in hemocytes of Mytilus coruscus, respectively, after infection with Staphylococcus aureus, Vibrio alginolyticus and Candida albicans (mean±SD, n=3)
綜上所述, 對厚殼貽貝中新發現的兩種抗菌肽myticalin和mytimacin 開展了序列分析, 固相化學合成, 功能驗證,以及表達譜分析。該研究證實了兩種抗菌肽分子在厚殼貽貝免疫防御過程中的作用,拓展了對貽貝抗菌肽分子多樣性的理解。同時, 該研究結果也為后續在基于貽貝抗菌肽結構與功能研究的基礎上, 深入了解貽貝抗菌肽的構效關系和抗菌機制, 并最終開發貽貝抗菌肽來源的新型生物抗生素奠定基礎。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
