棘莖楤木和樹參對林窗、林緣和林下生境的光合生理響應
2022-12-28 06:23:32李孝初王盼康華靖郭書亞周鈺鴻潘德月陳子林肖琛聞
浙江農業科學 2022年1期
李孝初,王盼,康華靖,郭書亞,周鈺鴻,潘德月,陳子林,肖琛聞
(1.寧波鄞州中銘農業科技有限公司,浙江 寧波 315040; 2.磐安縣中藥創新發展研究院,浙江 磐安 322300;3.溫州市農業科學研究院,浙江 溫州 325006; 4.浙江師范大學 植物學實驗室,浙江 金華 321004;5.浙江大盤山國家級自然保護區管理局,浙江 磐安 322300; 6.浙江省農業科學院,浙江 杭州 310021)
棘莖楤木(Araliaechinocaulis)屬五加科楤木屬,在我國甘肅、陜西、福建等地區均有分布[1],生于森林、灌叢或林緣路邊,垂直分布從海濱至海拔3 000 m左右。樹參(Dendropanaxdentiger)為五加科樹參屬植物,在我國南方地區分布廣泛,可生長在海拔10~2 000 m的常綠闊葉林或灌叢中[2]。前者具有鎮痛消炎、祛濕活血之效[3],同時也具有重要的營養價值,莖皮中含墩果酸、刺囊酸及馬栗樹皮素二甲酯等物質。后者的根[4]、莖[5]、葉[6]均有重要的藥用價值,可治偏頭痛、風濕痹痛等癥[7],嫩葉芽可以腌制、爆炒、涼拌等多種方式食用[6],味道鮮美,營養價值高。國內有關棘莖楤木和樹參的研究集中于幼苗繁育[8]、生長發育規律[9-10]、栽培技術[11]等方面。
發展林下經濟正成為充分利用林地資源、促進林業發展的一個新方向。利用林下特有生境種植藥用和食用植物,可提高林地資源利用效率,緩解耕地不足,增加森林經濟效益。光照是影響林下植物生長和產量的主要因子[12]。若光照過弱,則光合產物積累較少,影響植株生長;而光照過強,則會出現光抑制而降低光合作用,進而抑制植株的生長。探明棘莖楤木和樹參的適宜光照范圍及生理特性對其規模化推廣種植,批量化育苗等具有重要意義。為此,本試驗設置了林窗、林緣和林下3種生境,擬從光合氣體交換及葉綠素熒光參數兩個角度,探討棘莖楤木和樹參的適宜光強及光合生理特征,以期為其林下種植推廣提供參考。
1 材料與方法
1.1 試驗地點與處理設計
2020年2月在浙江省磐安縣祠下村杉木林林窗(A)、林緣(B)和林下(C)生境下分別種植棘莖楤木和樹參,日常實行統一管理。于2020年8月下旬進行試驗測量。每個生境選擇5~7株植物,每株植物選取1片頂部完全展開的葉子待測。
1.2 測定指標
利用便攜式光合儀(LI-6400-40, LI-COR INC., USA)于晴天9: 00—16: 30進行數據測量。數據測量前進行開機預熱及各項檢查。在自然光下誘導1.5~2.0 h后,采用開放式氣路,流速為400 μmol·s-1。根據預測量儀器中葉室溫度(Tblock)的動態變化,設定葉室溫度約為29.47 ℃(大氣溫度約為30.48 ℃左右),空氣相對濕度控制在45%~70%,光合有效輻射強度(PAR)梯度設置為:1 400、1 200、1 000、800、600、400、200、150、100、50和0 μmol·m-2·s-1,應用CO2注入系統提供400 μmol·mol-1穩定的CO2。在此條件下同時測定植物葉片的氣體交換參數和熒光參數。應用自動測量程序(FlrPn-Cacurve)進行數據測量。每次程序記錄最小等待時間為2 min,最大等待時間為3 min,數據記錄前儀器均自動進行參比室和樣品室之間的匹配。同樣,外界溫度、濕度、光強等環境因子數據也采用便攜式光合儀自帶的傳感器獲得。
1.3 數據分析
根據光合計算軟件4.1(井岡山大學生命科學學院/數理學院),應用光響應機理模型擬合電子傳遞速率對光強的響應曲線以及光合速率對光強的響應曲線。SPSS 19.0版本(SPSS, Chicago, IL)軟件進行顯著性分析。每個生境選取5個重復,采用Excel 2010作圖。
2 結果與分析
2.1 3種生境下的光強和溫度動態變化
由圖1可看出,以8:00—14:00的數據為例,A生境下的光強和溫度整體處于最高,其次是B生境,C生境下的光強和溫度則整體處于最低,說明林窗、林緣和林下生境具有較大差異。
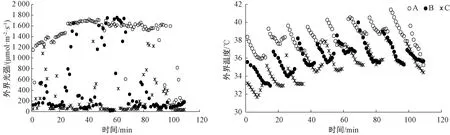
圖1 3種生境(A、B和C)下外界光強及溫度的動態變化
2.2 不同生境下棘莖楤木和樹參葉片氣體交換參數的響應
2.2.1 凈光合速率對光強的響應
由圖2可知,低光強(小于200 μmol·m-2·s-1)下,不同生境下的棘莖楤木和樹參葉片的凈光合速率均隨PAR的增加幾乎直線增加;在高光強(>800 μmol·m-2·s-1)下,其凈光合速率則均有明顯下降趨勢。不同生境下的光合速率值整體上有所差異。

圖2 不同生境下棘莖楤木和樹參葉片的凈光合速率對光的響應曲線
利用光響應機理模型擬合不同條件下光合作用對光的響應曲線,得到最大凈光合速率(Pnmax)、凈光合速率對應的飽和光強(Isat)、光補償點(Ic)和暗呼吸速率(Rn)等光合參數見表1。由表1可知,擬合曲線所得到的擬合值與其所對應的實測值高度符合,其確定系數均大于0.90。從最大光合速率來看,C生境下的棘莖楤木和樹參光合速率顯著較高,分別為19.325和12.451 μmol·m-2·s-1。而同一樹種在不同生境的飽和光強均較為接近,棘莖楤木和樹參的光飽和點分別約為900和800 μmol·m-2·s-1。另外,不同生境下棘莖楤木和樹參葉片的光補償點均較低,其范圍分別為19.590~28.392 μmol·m-2·s-1和13.200~23.829 μmol·m-2·s-1。

表1 不同生境下棘莖楤木和樹參葉片光合作用對光的響應曲線擬合結果
2.2.2 蒸騰速率及氣孔導度對光強的響應
由圖3可知,棘莖楤木葉片蒸騰速率隨著光強的增加而上升,高光強下趨于穩定;而樹參葉片蒸騰速率隨著光強的增加而呈上升趨勢。不同生境下棘莖楤木葉片的蒸騰速率整體上均存在顯著差異,以C生境下的值為最高;同樣,樹參葉片的蒸騰速率也以C生境下較高。由圖4可知,在較低光強(<400 μmol·m-2·s-1)下,不同生境下棘莖楤木葉片氣孔導度均隨著光強的增加先直線上升,而后均呈緩慢下降趨勢;而不同生境下樹參葉片氣孔導度的變化趨勢有所差異。另外,同一植物不同生境下葉片的氣孔導度整體上均存在顯著差異,且均以C生境下的值為最高。
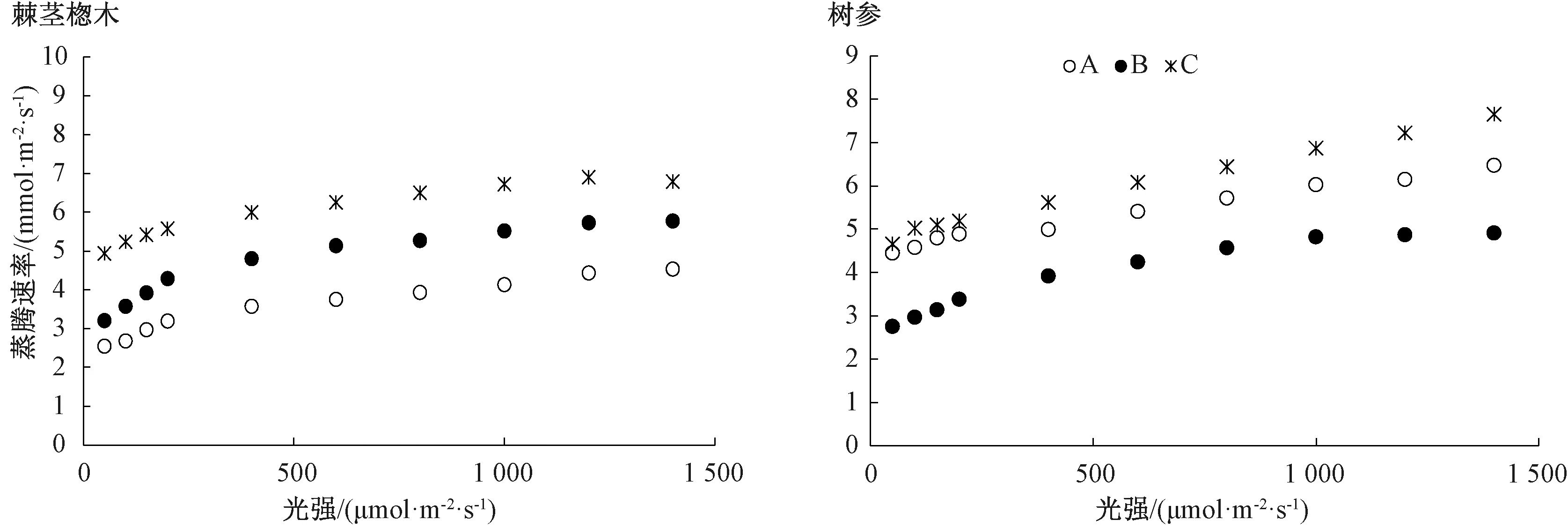
圖3 不同生境下棘莖楤木和樹參葉片的蒸騰速率對光的響應曲線

圖4 不同生境下棘莖楤木和樹參葉片的氣孔導度對光的響應曲線
2.3 不同生境下棘莖楤木和樹參葉片葉綠素熒光參數的響應
2.3.1 電子傳遞速率對光強的響應
由圖5可知,PAR<400 μmol·m-2·s-1時,棘莖楤木和樹參葉片的電子傳遞速率均隨PAR的增加幾乎呈直線增加;之后電子傳遞速率均在達到最大值后發生較為明顯的PSⅡ動力學下調現象。PAR大于600 μmol·m-2·s-1時,不同生境下棘莖楤木葉片的電子傳遞速率存在顯著差異;當光強高于500 μmol·m-2·s-1時,B和C生境下棘莖楤木葉片的電子傳遞率則有顯著差異;C生境下棘莖楤木和樹參的最大電子傳遞速率均為最高(P<0.05),分別為145.263和99.053 μmol·m-2·s-1。同樣,兩個樹種葉片的最大電子傳遞速率所對應的飽和光強也均以C生境下的值為最高,分別為1 044.110和847.615 μmol·m-2·s-1(表2)。

圖5 不同生境下棘莖楤木和樹參葉片的電子傳遞速率對光的響應曲線

表2 不同生境下棘莖楤木和樹參葉片光合電子傳遞速率對光的響應曲線結果
2.3.2 實際光化學量子產量(ΦPSⅡ)和天線轉化效率(F′v/F′m)對光強的響應
由圖6可知,不同生境下棘莖楤木和樹參葉片ΦPSⅡ均隨著光強的增加而下降。與B和C生境相比,A生境下棘莖楤木葉片ΦPSⅡ值整體較低,在相對較高光強(>400 μmol·m-2·s-1)下存在顯著差異;而不同生境下樹參葉片的ΦPSⅡ值均較為接近。不同生境下棘莖楤木和樹參葉片F′v/F′m對光強的響應見圖7。不同生境下棘莖楤木和樹參葉片F′v/F′m均隨著光強的增加而下降。A和C生境下,棘莖楤木葉片之間的F′v/F′m值在光強大于400 μmol·m-2·s-1時存在顯著差異;不同生境下樹參葉片之間的F′v/F′m值整體上無顯著差異,只有在光強大于1 000 μmol·m-2·s-1時A生境下的F′v/F′m值顯著較低。

圖6 不同生境下棘莖楤木和樹參葉片的實際光化學效率對光的響應曲線

圖7 不同生境下棘莖楤木和樹參葉片的F′v/F′m 對光的響應曲線
2.3.3 光化學猝滅系數(qP)和非光化學猝滅系數(qN)對光強的響應
由圖8可知,不同生境下棘莖楤木和樹參葉片qP均隨著光強的增加而整體呈直線下降。不同生境下,棘莖楤木和樹參(除高光強下樹參A和B生境之間)葉片的qP整體上均無顯著差異。由圖9可知,不同生境下棘莖楤木和樹參葉片qN均隨著光強的增加而下降,A和C生境下,棘莖楤木葉片的qN值整體上無顯著差異,而當光強較高時(≥400 μmol·m-2·s-1)樹參葉片qN值整體上差異顯著。

圖8 不同生境下棘莖楤木和樹參葉片的光化學猝滅系數對光的響應曲線

圖9 不同生境下棘莖楤木和樹參葉片的非光化學效率對光的響應曲線
3 小結與討論
光是影響植物光合作用的重要因子。不同的光照條件會影響到植物的光合生理特性,以及葉綠素、可溶性糖、蛋白質含量和各種酶的活性,最終會影響到植物的生長,體現在苗高、地徑、根重及生物量的變化[13-16]。本試驗的環境因子監測表明,林窗生境下的光照強度相對較高,林緣其次,林下生境的光照強度最低,這一結果與前人的研究結果相似[17]。研究不同光環境下植物的光合特性可了解其對光照強度變化的生理適應性。柯世省等[18]研究表明,云錦杜鵑(Rhododendronfortunei)樹苗對林窗和林緣不同強度光照存在不同的生理響應,林緣是林下云錦杜鵑最適宜的轉移生境。
明確植物的光飽和點對植物的高效栽培至關重要。本文應用光響應機理模型可很好地擬合不同生境條件下棘莖楤木和樹參葉片電子傳遞速率和凈光合速率對光的響應曲線,而且得到的主要光合參數與實測值高度符合(R2>0.99),這與其他學者的結論相同[19,21]。光合速率對光的響應曲線結果表明,棘莖楤木和樹參的光飽和點分別約為900和800 μmol·m-2·s-1;電子傳遞速率對光的響應曲線結果表明,棘莖楤木和樹參的光飽和點范圍分別為867.957~1 044.110 μmol·m-2·s-1和755.893~847.615 μmol·m-2·s-1,兩個角度擬合的結果一致性較好。由此可見,棘莖楤木和樹參均為半陽生植物,其最適宜光強約分別為900和800 μmol·m-2·s-1。這與吳曉云等[22]的相關研究結果基本一致。林下種植物種選擇關鍵是其對林下弱光環境的適應能力[12]。光合速率對光的響應曲線擬合結果表明,棘莖楤木和樹參葉片的光補償點較低(約20 μmol·m-2·s-1),說明棘莖楤木和樹參利用弱光的能力較強。另外,B和C生境下棘莖楤木和樹參葉片光補償點顯著降低,說明棘莖楤木和樹參可通過生理調節主動適應弱光環境。這與楊亞男等[23]對四季桂(Osmanthusfragrans)的研究結果相一致。由此可見,棘莖楤木和樹參適合林下種植。
蒸騰作用是植物對水分吸收和運輸的主要動力,可促進植物體對礦質元素的吸收,在高溫條件下可通過水分蒸發帶走大量熱量,降低葉片的溫度,從而減少高溫對葉片的灼傷。熊慧等[24]發現,生境對植物氣孔密度和長度有顯著影響,由于林下光照強度低,相對濕度高與土壤含水量高,所以林下植物以較低的蒸騰成本可獲得較大的光合效益,這與本試驗結果一致。3種生境下的棘莖楤木和B生境下樹參的蒸騰速率都隨光照強度的增加而上升,隨后趨于穩定,而A和C生境下樹參的蒸騰速率在光照強度為1 500 μmol·m-2·s-1時未達到最大值。不同生境下棘莖楤木和樹參葉片的蒸騰速率都存在顯著差異,兩種植物C生境下的蒸騰速率均處于最高。葉片氣孔導度表示氣孔張開的程度,這影響著植物的光合作用,蒸騰作用。本研究發現不同生境下棘莖楤木和樹參葉片的氣孔導度整體上均存在顯著差異,以C生境下的值為最高。
光化學淬滅系數qP反映PSⅡ天線色素吸收的光能用于光化學電子傳遞的份額,并進一步反映PSⅡ反應中心的開放程度和電子傳遞活性的大小。蘇金等[25]對不同遮陰下紫珠(Callicarpabodinieri)的光合特性研究表明,與全日照相比,遮陰處理會顯著降低葉片的qP值。本試驗結果顯示,棘莖楤木和樹參葉片qP在不同生境下整體上均無顯著差異。另外,除用于光合作用的光能外,還有一部分光能可通過非光化學猝滅(qN)過程保護PSⅡ反應中心免受因吸收過多光能而引起的光氧化和光抑制傷害。本試驗結果表明,不同生境下棘莖楤木葉片的qN值整體上無顯著差異,而光強相對較弱的生境(B和C)下樹參葉片qN值整體相對較高,說明棘莖楤木以及B和C生境下樹參葉片具較好的耗散過量能量能力。ΦPSⅡ的明顯降低是光抑制發生的可靠判據[26]。不同生境下的結果顯示,A生境下棘莖楤木葉片ΦPSⅡ值整體較低,B和C生境下的ΦPSⅡ值極為接近;而3種生境下樹參葉片的ΦPSⅡ值均較為接近。由此可見,A生境下的棘莖楤木發生了一定程度的光抑制。
綜合本試驗結果表明,林下生境的棘莖楤木和樹參葉片的最大光合速率、蒸騰速率、氣孔導度,以及最大電子傳遞速率、ΦPSⅡ和F′v/F′m值整體上均顯著高于林窗和林緣生境;同時,在林窗和林緣生境下,棘莖楤木和樹參也具有較強的耗散過量能量能力。結合葉片光飽和點和光補償點范圍,認為棘莖楤木和樹參均屬于半陽生樹種,且兩者對弱光的利用能力較強,適合林下種植。建議在浙中低海拔山區種植時,生境光強為800~900 μmol·m-2·s-1最為適宜。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
