論雜種優勢機理的邏輯體系
2022-12-28 05:01:54潘春桂
廣西農學報 2022年4期
潘春桂
(安徽省東至縣農業農村局,安徽 池州 247210)
揭示雜種優勢的產生機理,無論是對于遺傳學本身學科的發展,還是對于在實踐中提高雜種優勢利用效率,增加經濟效益都具有極其重要的意義。到目前為止,關于雜種優勢機理學說和研究方法的多樣性并未澄清其發生機理[1],研究者均寄希望于遺傳學等學科的進一步發展。很少看到對雜種優勢必然性的哲學思考。目前主要靜態地研究雜種一代與雙親的差異,對外部條件的作用機制幾乎很少研究。如果假設雜種優勢初始值為零,它能夠和外部條件相互作用發生轉化嗎?影響雜種優勢的外部條件很多,環境養分供給可能是最重要及最易受控的條件。潘春桂于2020年提出了一種植物養分獲取總量雜種優勢模型[2],本文在這個模型的基礎上,對不同的養分環境供給量之下雜種優勢的變化進行分析,以期獲得有益的啟示,希望引發對雜種優勢機理邏輯體系的思考。
1 模型與分析
假設環境中各種養分供給量是一定的,植株個體存在這樣一種養分吸收過程,吸收過程可分為兩個階段:固定速度吸收期(飽和期)及停止吸收期(停止期),各種養分停止吸收時刻或轉折點由最先進入自身臨界點的養分所決定,將這種相對虧缺的養分稱為主導養分。這里認為不同養分臨界環境消耗率在同一遺傳類型中相同,雜種一代為加性遺傳,飽和期各種養分吸收速度也為加性遺傳。在某一時刻任一遺傳類型養分獲取總量由各種養分吸收速度大小及它們進入自身臨界點同步性所決定。
先簡單介紹養分的獲取過程,首先設環境三種養分A、B、C總供給(vt,質量單位)分別為5、4、3,當任一種養分臨界消耗率為K時,三種養分臨界消耗量(vt)分別為5 K、4 K、3 K。對于P1,三種養分A、B、C的吸收速度(v,質量時間比)分別為3、2、1,臨界消耗率為100%時,三種養分臨界消耗量(vt)分別為5、4、3,經過時間1.66(t,時間單位)時,A、B、C三種養分消耗量(vt)分別為5.00、3.32、1.66,可看出A養分已達到臨界消耗量,1.66(t)就是轉折點,可稱A養分為主導養分。這時個體停止對各種養分的吸收,即由飽和期進入停止期,養分吸收總量停止在5.00+3.32+1.66=10(vt)上。對于P2,養分臨界消耗率為50%,則三種養分臨界消耗量(vt)分別為2.5、2、1.5,它們吸收速度(v)分別為2、3、1,經過0.66(t)時,三種養分消耗量(vt)分別為1.32、2、0.66,養分B達到臨界消耗量,總吸收量停止在(2+3+1)×0.66=4.00(vt),0.66(t)就是轉折點(時刻),可稱B養分為主導養分,當然轉折點之前養分吸收總量為三種養分吸收速度之和與時間乘積。對于雜種一代P1×P2,養分臨界消耗率為加性遺傳,為75%,三種養分臨界消耗量(vt)分別為3.75、3.00、2.25,各種養分飽和期吸收速度也為加性遺傳,則三種養分A、B、C吸收速度(v)分別為2.5、2.5、1,在1.2(t)時,養分B達到臨界點,1.2(t)就是轉折點,可稱B養分為主導養分,總吸收量停止在(2.5+2.5+1)×1.2=7.2(vt)。如果在2(t)時進行(取樣)計算,以上組合的總養分獲取量離親值為0.20(vt),中親優勢為2.85%。先將各種養分吸收速度的衰減定義為各種養分速度相對于其飽和速度的減小比例,再從養分吸收速度衰減角度看看2(t)時雜種優勢是如何形成的。給出兩種定義,對于每種遺傳類型,(1)固有衰減:每種養分的吸收速度按照本身所處狀態(由飽和吸收速度、自身臨界消耗量等決定)的衰減;(2)實際衰減:每種養分被迫與固有衰減最大的養分(主導養分)同步衰減。例如,在2(t)時,P1對A養分吸收速度的固有衰減為(1-1.666/2)×100%= 16.7%,對于B、C兩種養分,仍處于飽和期,固有衰減均為0.0。因此P1對三種養分實際衰減均為16.7%。再看對A養分吸收速度中親值固有衰減為{1-(3×1.666/2+2×1.25/2)×0.5/[(3+2)× 0.5]}× 100%=25.0%,對A養分吸收速度中親值實際衰減為{1-(3×1.666/2+2×0.666/2)×0.5/[(3+2)× 0.5]}×100%=36.7%。2(t)時各種養分吸收速度固有及實際衰減,詳見表1。
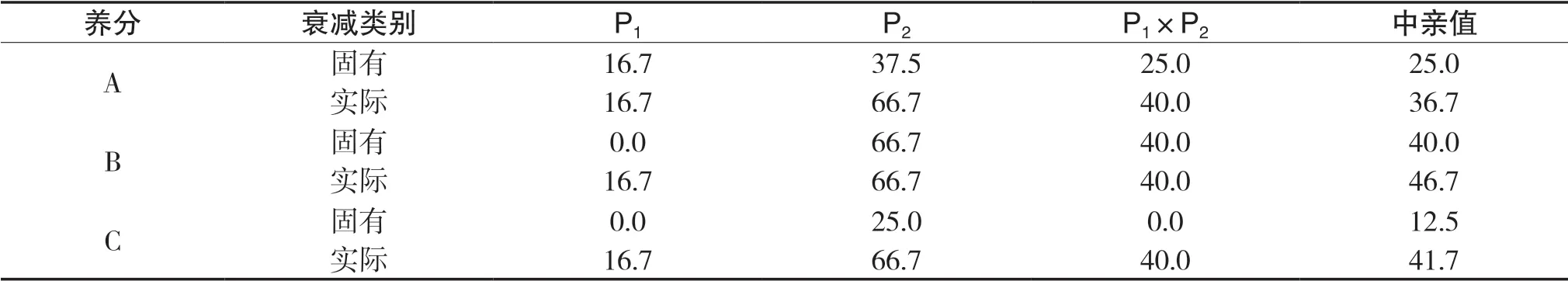
表1 2(t)時各種養分吸收速度固有及實際衰減 單位:%
由以上分析可以推論,當雙親主導養分不同時,則可能導致每種養分吸收速度中親值的實際衰減程度大于自身固有衰減,而中親值固有衰減可認為大致等于雜種一代的固有衰減;如果雜種一代各種養分進入自身臨界點同步性好或完全同步,各種養分吸收速度的固有衰減程度相近或相同,則雜種一代每種養分固有衰減等同于實際衰減。即可得出結論:每種養分吸收速度中親值的實際衰減程度大于雜種一代實際衰減。這有利于獲得較大的雜種優勢。當雙親的主導養分為同一養分時,這使雜種一代各種養分進入自身臨界點難以或不能達到同步,各種養分中親值與雜種一代的實際衰減相近,自然不利于雜種優勢的形成。給予簡單證明,設三種養分供給量為M、N、G,P1三種養分飽和吸收速度為a1、b1、c1,P2三種養分飽和吸收速度為a2、b2、c2,根據 條 件,M/a1<N/b1、M/a1<G/c1及M/a2<N/b2、 M/a2<G/c2,則可得:2 M/(a1+a2)<2 N/(b1+b2)、2 M/(a1+a2)<2 G/(c1+c2),即雜種一代不能達到同步性:2 M/(a1+a2)=2 N/(b1+b2)=2 G/(c1+c2)。可看出同雙親一樣,雜種一代第一種養分也為主導養分,其吸收速度固有衰減介于雙親之間,則雜種一代每種養分實際衰減也介于雙親之間,在很多情況下,這不利于形成較大的總養分獲得量離親值。當雙親臨界消耗率不相等時,也存在同以上相似的結論,只需初等數學就能證明(此處省略)。容易看出,對中親優勢的負效應的不確定性有所增加。2(t)時P1×P2在不同養分供給條件下的總養分獲得量的雜種優勢詳見表2。從下文可以看到,當某一種養分嚴重短缺時可形成這種現象,這直接證明本文結論的正確性。將主導養分在雙親中是否相同,雜種優勢大小存在差異的現象稱為主導養分效應。

表2 2(t)時P1×P2在不同養分供給條件下的總養分獲得量的雜種優勢
如果保持雙親各種養分飽和吸收速度及臨界消耗率如上文一樣,只改變養分供給量,從表2可看出,在第2種養分條件下,各種養分均減少了60%,離親值亦減小60%,中親優勢沒有改變。在第3、4、5養分條件下,分別只有一種不同養分減小了60%,可見這種減少的養分均為雙親的主導養分,這時離親值均顯著減小,中親優勢也多有減小。在第6種養分條件下,養分供給總量未變,只是B、C兩種養分量互有增減,雜種一代各種養分進入自身臨界點完全同步,這時獲得了較大的離親值與中親優勢。
2 討論和結論
大量試驗表明,單一缺乏某養分如缺氮、缺磷、缺鉀、缺水時,中親優勢或離親值近似為零[3-7]或顯著減小[8-13],這與上文分析的結論是一致的。當環境中某種養分嚴重短缺時,則可能這種養分對于雙親同樣屬于相對性缺乏或同為主導養分,可導致單株養分獲取總量離親值顯著減小,從而引發生物量或產量的離親值顯著減小,這是由于兩者之間存在內在緊密的聯系,在不同作物的雜交試驗中,均存在類似的結論。這表明,主導養分效應在雜種優勢機理中占有重要的地位,這必將引發對目前雜種優勢機理研究方法的重新審視或反思。可以設想,當環境中所有養分不足時比僅一種養分不足時可能會產生較大的離親值或中親優勢[14,15],因為后者雙親主導養分可能相同。非常有必要通過試驗對這種現象進行進一步驗證。
人類認識到雜種優勢現象已有近兩個世紀,一直以來,科學家提出了各種假設試圖解釋其形成機制。近年來由于分子遺傳學的快速發展,已經在多個層面上獲得了進展[16-18]。目前的研究方法是基于這樣的一種邏輯體系,由于雜種一代是雙親兩套遺傳物質的結合,不存在新的遺傳物質,因此雜種優勢必定來源于它們的相互作用,這導致了雜種一代獲得了有別于雙親的分子生物學或遺傳學的特異性表現或過程[19,20]。但這種方法已面臨了一個窘境,即當外部試驗條件改變時,結論可能不一致。另外一點,科學家可能有意無意忽略了這樣一個問題,雜種一代這種內在的、微觀的特異性是否屬于一種本能的或自然的生命過程,也可能如雜種一代外在的、可視的表現一樣,如具有較大的營養體或旺盛的生長勢,也是雜種優勢的一種表現,而不是其形成原因。因為已經有研究表明,環境效應可誘導出基因表達的差異性[21]。從上文模型與分析可看出,表面上雜種一代與雙親是處在相同的外部條件之下,如果這些條件具備某些特征,便能夠使雜種優勢完成了從無到有的轉化。在模型中并沒有預設雜種一代具有區別于雙親的特異性,這種特異性可有利于雜種優勢的形成。而是僅假設了各種養分飽和吸收速度及臨界環境消耗率均為加性遺傳。既然最終形成了總養分獲得量的雜種優勢,必然反過來可能使雜種一代發生內在分子生物學或遺傳學的改 變[22],形成某種內在特異性,可見這種內在特異性并不是本能或自然形成的,是與外界條件相互作用的結果,并不能認為它是雜種優勢產生的原因,它與雜種一代外在表現特異性處于同等地位。簡單地說,當這種外部環境條件對雜種一代相對有利時,便產生的雜種優勢。可見目前研究的邏輯體系存在很大的局限性。上文模型與分析重建了一種全新的邏輯體系,包括兩個方面:第一,不存在必然的雜種優勢,即使存在,也屬于偶然性。第二,雜種優勢主要是在外界條件作用下轉化形成的。因此研究的重點應著眼于轉化的條件及過程,而不是像以往那樣,主要針對于雜種一代與雙親的差異性。由于試驗要素中已包含了外部條件,也避免了像以往那樣,不同條件下,結論可能存在差異。雖然植物生物量大多來自于光合產物,但與植株獲得的養分存在顯著的正相關關系[23]。本文簡單地以環境養分供給量作為試驗條件,并假定各種養分飽和吸收速度為加性遺傳,由于環境中養分消耗或不足,當雜種一代各種養分吸收速度相對于其飽和速度的衰減如果小于中親值或雙親的相應衰減,便產生了雜種優勢。目前形成的共識是,雜種優勢機理問題在現階段科學發展水平之下是不能得到解決[24],只有對未來有所期待。
