杉虎斑Galectin-8基因克隆及其在不同阿魏酸水平飼料下的表達響應
2023-01-05 07:48:44付偉杰鄺杰華羅君黃建盛陳有銘陳剛
生物技術通報 2022年12期
付偉杰 鄺杰華 羅君 黃建盛 陳有銘 陳剛
(1.廣東海洋大學水產學院,湛江 524088;2.廣東藍糧種業有限公司,湛江 524000)
半乳糖凝集素(galectins)屬于糖結合蛋白家族中的一員,通過保守的糖識別結構域(carbohydrate binding domain,CRD)結合β-半乳糖苷類,不依賴Ca2+的活性,并具有多種生物活性功能,包括細胞遷移、凋亡、免疫反應、炎癥細胞信號和神經保護等[1-2]。半乳糖凝集素在進化上是保守的,廣泛存在于從原始到高級的各種生物中,并在不同的組織與細胞中表達[3-5]。半乳糖凝集素主要位于細胞內,也存在于細胞外,其缺乏一個N 端信號肽序列來引導它們通過內質網進行分泌,但可以通過非經典途徑分泌到細胞外,從而具有細胞外功能[6]。此外,細胞內的半乳糖凝集素參與細胞功能過程,如前體mRNA 剪接、細胞生長調節和細胞周期過程[7-8]。半乳糖凝集素不但在微生物感染中發揮關鍵作用,也在先天性和適應性免疫反應中發揮潛在作用。迄今為止,已經在哺乳動物中鑒定出15 種半乳糖凝集素。根據結構特征、糖結合特異性和生物學功能,半乳糖凝集素可分為3 種類型:原型(Gale ctin-1、-2、-5、-7、-10、-11、-13、-14 及-15),含有一個CRD;串聯重復類型(Galectin-4、-6、-8、-9 和-12),包含兩個由連接肽連接不完全相同的CRD;嵌合體型(Galectin-3),由N 端的非凝集素結構域和C 端的CRD 所組成[9-10]。
Galectin-8 是一種串聯重復型半乳糖凝集素,具有氨基酸序列相似性約35%的N 端糖識別結構域(N-CRD)和C 端糖識別結構域(C-CRD),并且兩個結構域中均含有保守基序H-NPR 與WG-EE[11]。Galectin-8 在先天性和適應性免疫中具有重要作用。哺乳動物Galectin-8 可以通過監測核內體和溶酶體的完整性來檢測入侵的細菌,然后激活抗菌自噬以保護細胞免受細菌的感染[12]。據報道,半滑舌鰨(Cynoglossus semilaevis)的Galectin-8 對某些革蘭氏陰性菌病原體具有強大的殺菌活性,可直接破壞它們的膜和結構[13]。Galectin-8 可促進漿細胞的形成和調節其分化,并在促進抗體產生的過程中起到附加和冗余作用[14-15]。Galectin-8 通過串聯重復結構與白細胞介素2 和轉化生長因子-β 受體相互作用,促進T 細胞的增殖與分化[16]。此外,Galectin-8 被認為是一種早期的腦保護劑,不僅能抑制炎癥信號,促進抗氧化活性,還能減少β 淀粉樣蛋白寡聚化與神經原纖維纏結沉淀,增強對阿爾茨海默病的預防作用[2]。目前,關于Galectin-8在硬骨魚類中的研究僅見于尼羅羅非魚(Oreochromis niloticus)[17-18]、大彈涂魚(Boleophthalmus pectinirostris)[19]、黑巖魚(Sebastes schlegelii)[20]、牙鲆(Paralichthys olivaceus)[21]、鱖(Siniperca chuatsi)[22]和半滑舌鰨[13],缺乏關于石斑魚Galectin-8的研究報道。
杉虎斑是由雌性褐點石斑魚(Epinephelus fuscoguttatus)與雄性清水石斑魚(E.polyphekadion)雜交的子一代,具有雜交優勢[23]。石斑魚具有營養豐富、肉質鮮美和經濟價值高等特點,已在十幾個沿海國家中廣泛養殖[24]。據聯合國糧食及農業組織(FAO)統計,2019年全球石斑魚養殖總產值為820 794 707 美元,養殖總產量更是達234 828.45 t,其中中國的養殖產量最大,占總產量的78%,多達183 127 t[25]。然而,高密度的集約化養殖與不恰當的投喂會損害魚類的免疫系統,使其易受病原體的感染[26]。研究報道,飼料中添加免疫刺激劑可以作為預防和治療水生動物感染性疾病的有效措施。草本植物及其衍生物可作為一種廉價的抗生素替代物,可以提高魚類的免疫和生長性能[27]。阿魏酸(ferulic acid,FA)屬于天然的植物化學物質,是一種低毒酚酸,可在植物界中廣泛獲得。阿魏酸具有多種生理活性功能,如抗菌、抗炎、抗氧化、抗癌、抗病毒和保護肝臟等[28]。研究石斑魚的免疫相關基因和增強免疫的方法對于獲得高產量至關重要。因此,本研究運用RACE 技術首次克隆出杉虎斑Galectin-8的全長cDNA 序列,然后對其進行生物信息學分析,并通過實時定量PCR(RT-qPCR)分析Galectin-8mRNA 的組織特異性分布;此外,還研究了飼料中添加不同水平阿魏酸對Galectin-8基因表達的影響,以期為進一步探究魚類免疫防御機制提供一定的依據,同時為控制水產養殖疾病暴發提供新的思路和策略。
1 材料與方法
1.1 材料
試驗用杉虎斑幼魚購自廣東湛江東簡鎮石斑魚苗場,在湛江恒興南方海洋科技有限公司室內1m3玻璃鋼桶中暫養馴化15 d,期間投喂石斑魚商品配合飼料。配制7 組添加不同水平FA 的等氮等脂試驗飼料,FA 水平分別為0(對照組)、40、80、160、320、640 和1 280 mg/kg,試驗飼料組成及營養水平見表1。暫養結束后,將試驗魚饑餓24 h,稱重后將初始體重為(9.55 ± 0.60)g 的試驗魚分組,挑選大小基本一致且健康的試驗魚,隨機分配到21 個玻璃鋼纖維桶中,養殖10周。試驗共設計7個處理組,每處理3 個重復,每重復1個0.5 m3的玻璃鋼纖維桶,每桶放30 尾魚。每天表觀飽食投喂2 次(8:00 和17:00),日投喂量約為石斑魚體重的3%-5%。試驗用水為經過沙濾和沉淀的天然海水,光照周期為自然光照,試驗期間不間斷充氧氣,溶解氧量不低于6 mg/L,鹽度為29%,溫度范圍為25-30℃,pH 7.6-8.2,且氨氮和亞鹽含量不大于0.05 mg/L。

表1 試驗飼料組成及營養水平Table 1 Ingredient composition and nutrient levels of the experimental diets
1.2 方法
1.2.1 樣品采集 養殖試驗結束后,將試驗魚禁食24 h,采樣時使用丁香酚進行麻醉。首先在對照組中進行取樣,每桶隨機取3 尾魚,解剖分離得到腦、胃、肝臟、鰓、腸道、中腎、頭腎、心臟、脾臟、肌肉和皮膚。在其余的處理組中,每桶取3 尾魚進行解剖獲得肝臟、腸道、脾臟及頭腎。各組織樣品收集于裝有RNA Later 的2 mL 無菌離心管中,液氮快速冷凍后置于-80℃超低溫冰箱中用于RNA 的提取。
1.2.2 RNA 提取及第一鏈cDNA 的制備 按照說明書,使用TranZol Up 試劑(TransGen)提取各組織總RNA。采用超微量分光光度計(SimpliNano,Biochrom,UK),在260 nm 條件下檢測RNA 的濃度,并用260 nm 與280 nm 的比值(范圍在1.8-2.1)測定其的純度。通過1.5%的瓊脂糖凝膠進行電泳,檢測提取的RNA 質量。通過反轉錄試劑盒EasyScript?One-Step gDNA Removal and cDNA Synthesis SuperMix(TransGen),在PCR 擴增儀中設置42℃ 15 min;85℃ 5 s,將1 μg RNA反轉錄成cDNA,并保存于-80℃用于之后的基因克隆和RT-qPCR 分析。
1.2.3Galectin-8基因全長cDNA 的克隆 從課題組的杉虎斑轉錄組數據中篩選出注釋為Galectin-8的Unigene,并在NCBI 中進行BLAST 比對確定為魚類的Galectin-8基因片段。使用Primer Premier 5.0 設計特異性引物Gal-8-F 和Gal-8-R 克隆中間片段,PCR 程序設置為:95℃ 5 min;95℃ 30 s,60℃ 30 s,72℃ 45 s,共循環35 次;72℃ 10 min。用1.5%的瓊脂糖凝膠電泳檢測PCR 產物,并使用EasyPure Quick Gel Extraction Kit(TransGen) 進行目的片段切膠回收,并連接到pMD-18T Vector(TaKaRa),轉化到DH-5α 感受態細胞(TaKaRa)后,挑選出單克隆陽性菌落,隨后送至上海生工生物工程公司進行測序。按照SMARTer?RACE 5′/3′ Kit(TaKaRa)的操作說明,并根據獲得的中間片段序列,分別設計兩個基因特異性引物(Gal-8-5′GSP 和Gal-8-3′GSP)和兩個巢氏基因特異性引物(Gal-8-5′NGSP 和Gal-8-3′NGSP)。5′-RACE和3′-RACE cDNA 模板合成完后,進行cDNA 5′端和3′端的擴增,PCR 產物經純化后,進行切膠回收,連接,轉化與測序。根據重疊部分將測序得到的5′端和3′端序列與中間片段序列進行拼接,得到Galectin-8基因全長cDNA 序列。引物序列詳見表2。
1.2.4 生物信息學分析 使用ORF finder 在線工具(https://www.ncbi.nlm.nih.gov/orffinder/)來識別開放閱讀框序列(ORF)和預測氨基酸序列。應用Simple Modular Architecture Research Tool(SMART)(http://smart.embl-heidelberg.de)對Galectin-8 的結構域進行分析。信號肽和跨膜區的預測分別使用signalIP 5.0(https://services.healthtech.dtu.dk/service.php?SignalP-5.0) 和TMHMM 2.0(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)。利用BLAST 程序(http://blast.ncbi.nlm.nih.gov/Blast.cgi/)對cDNA 序列和氨基酸序列進行同源分析。通過DNAMAN 9.0 對氨基酸序列進行多序列比對分析,并用MEGA X 軟件,采用鄰接法(Neighbor-Joining)構建系統發育進化樹,bootstrap 值設置為1 000。多序列比對與構建系統發育進化樹所用的氨基酸序列見表3。采用SWISS-MODEL(http://www.swissmodel.expasy.org)程序進行蛋白質三級結構的構建,ProtParam tool(http://web.expasy.org/protparam/)預測蛋白質的理化性質(分子量、理論等電點和親水性)。蛋白質二級結構和亞細胞定位的預測分別采用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html/) 和PSORT II Prediction(http://psort.hgc.jp/form2.html)。使用ExPASy 服務器的NetNGlyc 1.0 server(https://services.healthtech.dtu.dk/service.php?NetNGlyc-1.0) 和NetPhos 3.1server(https://services.healthtech.dtu.dk/service.php?NetPhos-3.1)分別進行N-糖基化位點和磷酸化位點的預測。

表3 杉虎斑Galectin-8 氨基酸序列多重比對與構建系統進化樹所用到的氨基酸序列Table 3 Amino acid sequences used for multiple sequence alignment and phylogenetic tree construction for Galectin-8 from hybrid grouper
1.2.5 實時定量PCR 根據克隆獲得的cDNA 序列,設計實時熒光定量引物Gal-8-RT-F 和Gal-8-RT-R(表2)。在LightCycler 96(Roche,Switzerland)儀器中進行RT-qPCR,使用的是PerfectStartTMGreen qPCR Super Mix(TransGen)試劑盒。10 μL 反應體系包括5 μL 的2×PerfectStartTMGreen qPCR Super Mix、0.5 μL 的正向引物、0.5 μL 的反向引物、0.5 μL 的cDNA和3.5 μL 的ddH2O(Double distilled water)。設置反應程序為94℃ 30 s(1 個循環);94℃ 5 s,60℃ 15 s,72℃ 10 s(40 個循環);95℃ 10 s,65℃ 60 s,95℃1 s(1 個循環);37℃ 30 s(1 個循環)。通過Light-Cycler? 96 Software 分析熒光定量數據,獲得Cq 值。以β-actin為內參基因,用2-ΔΔCt方法計算基因的相對表達量。在統計分析之前,對數據進行了正態性和同質性檢驗,并使用SPSS 22.0 軟件進行單因素方差分析(One-Way ANOVA),組間若有顯著性差異,采用Duncan 氏法進行多重比較,P<0.05 表示差異顯著。每個處理組均有3 個生物學重復。
2 結果
2.1 Galectin-8基因cDNA全長克隆及序列分析
杉虎斑Galectin-8基因的cDNA 全長為1 368 bp(GenBank 登錄號:OL826837),其中5′非翻譯區(5′untranslated region,5′UTR)為16 bp,3′UTR 為394 bp,ORF 長960 bp,共編碼319 個氨基酸,還包括一個典型的加尾信號序列(AATAAA)和一個長為27 bp 的poly(A)尾巴(圖1)。Galectin-8 蛋白分子質量為35.26 kD,理論等電點為7.65,總平均親水性為-0.149(親水),不穩定系數為40.82(不穩定)。TMHMM 2.0 和signalIP 5.0 預測結果顯示,杉虎斑Galectin-8 蛋白無跨膜結構域與信號肽。亞細胞定位結果表明,Galectin-8 蛋白定位于細胞質、細胞核、線粒體、內質網和分泌小泡的概率分別為60.9%、21.7%、8.7%、4.3%和4.3%。NetNGlyc 1.0 server 和NetPhos 3.1server 預測結果表明,Galectin-8 蛋白含有1 個N-糖基化位點和29 個磷酸化位點(圖1)。
SMART 預測結果顯示,Galectin-8 蛋白含有兩個CRD,分別位于N 末端(15-150,136 aa)和C末端(187-319,133 aa),屬于串聯重復型,由序列長為36 個氨基酸的連接肽(Linker peptide)連接。此外,在兩個CRD 上均鑒定出兩個與β-半乳糖苷類結合的保守基序,分別為 HXNPR 和WGXEE,其中X 表示任意氨基酸(圖1)。利用SWISS-MODEL對杉虎斑Galectin-8 蛋白進行同源建模(圖2),發現與人的Galectin-8 三級結構相似,序列相似性為56.94%,GMQE 為0.78。其中,α- 螺旋占2.51%,β-轉角為8.46%,延伸鏈占33.86%,無規則卷曲為55.17%。

圖1 杉虎斑Galectin-8 的全長cDNA 核苷酸序列和推導的氨基酸序列Fig.1 Full length cDNA nucleotide and deduced amino acid sequences of Galectin-8 in hybrid grouper

圖2 同源建模預測的Galectin-8 蛋白三維結構Fig.2 Tertiary structure of Galectin-8 predicted by homology modeling
2.2 同源性及系統發育進化樹分析
杉虎斑與其他物種Galectin-8 的氨基酸序列同源性分析結果如圖3所示。通過NCBI 上blastp 比對,分別獲得杉虎斑Galectin-8 推斷的氨基酸序列與鞍帶石斑魚、許氏平鲉、藍鰭金槍魚、條石鯛、?魚、大西洋鮭、智人、原雞和熱帶爪蟾的一致性(表3),其中與鞍帶石斑魚的一致性最高,達到98.43%,許氏平鲉次之為82.61%,而與熱帶爪蟾的一致性最低為49.69%。同源性分析表明,Galectin-8 在不同動物中均具有保守性。參與氨基酸多序列比對的各物種Galectin-8 均具有串聯重復半乳糖凝集素的典型結構特征,包括一個N-CRD 和一個C-CRD,并且兩個CRD 均有兩個保守基序H-NPR 和WG-EE。

圖3 杉虎斑與其他物種 Galectin-8 氨基酸序列的多重比對Fig.3 Multiple sequence alignment of Galectin-8 amino acid sequences between hybrid grouper and other species
為了研究杉虎斑Galectin-8 與其他脊椎動物的進化關系,采用鄰接法構建系統發育進化樹(圖4),共包括20 個代表物種的氨基酸序列。結果表明,杉虎斑與其他硬骨魚類的Galectin-8 單獨聚為一支,爬行綱、兩棲綱、哺乳綱和鳥綱動物的Galectin-8 各自聚為一支。杉虎斑Galectin-8 與鞍帶石斑魚具有最近的親緣關系,許氏平鲉次之,而與黃姑魚的親緣關系相對較遠,與爬行綱、兩棲綱、哺乳綱和鳥綱其他動物的親緣關系相對更遠。
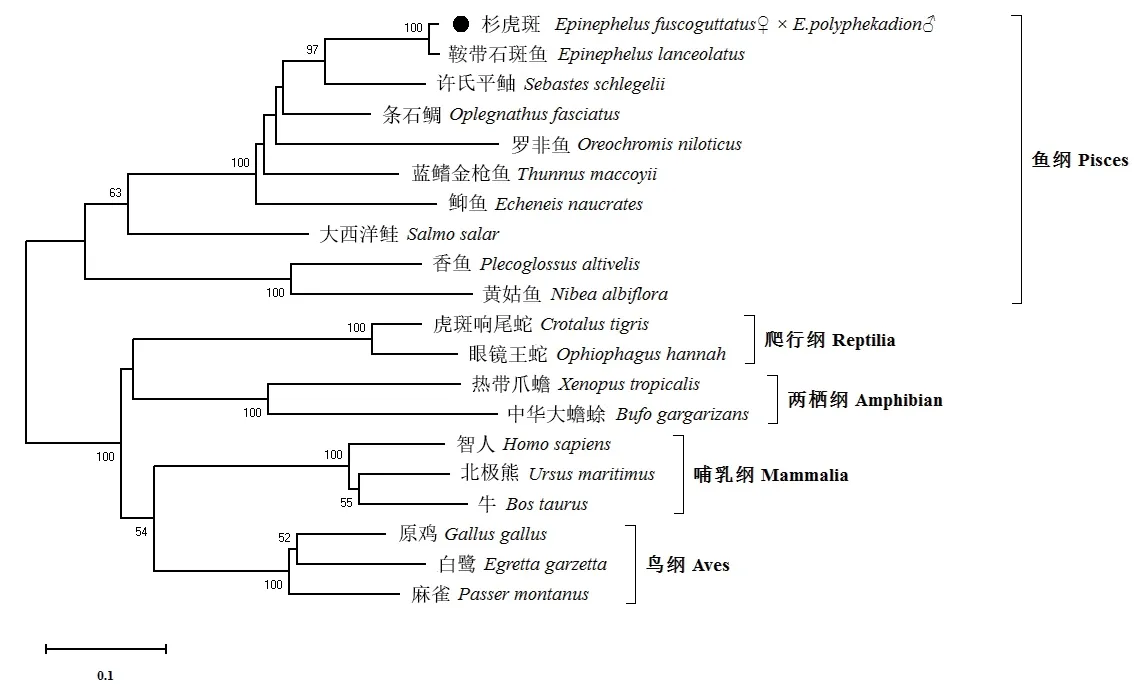
圖4 鄰接法構建Galectin-8 基因氨基酸序列的系統發育進化樹Fig.4 Phylogenetic tree of the amino sequences of Galectin-8 genes constructed by neighbour-joining method
2.3 杉虎斑Galectin-8基因組織表達分析
利用RT-qPCR 分析了Galectin-8在杉虎斑不同組織類型中的表達情況。結果顯示(圖5),Galectin-8在所有被檢測的組織中均有表達,且其在不同組織中的表達水平不同。Galectin-8在脾臟中表達量最高,其次是中腎、心臟、腸道、肝臟、頭腎和鰓,而在腦、胃、皮膚和肌肉組織中的表達量相對較少。

圖5 杉虎斑Galectin-8 在各組織的相對表達量Fig.5 Relative expressions of Galectin-8 in the different tissues from hybrid grouper
2.4 飼料中添加不同水平阿魏酸對杉虎斑Galectin-8基因表達的影響
根據圖6可知,杉虎斑Galectin-8基因的表達受到阿魏酸(FA)的影響。隨著飼料中FA 水平的增加,Galectin-8表達量在肝臟、頭腎和脾臟中均呈現先升高后降低的趨勢。FA 添加水平為40、80、160 和320 mg/kg 時,肝臟、頭腎和脾臟的Galectin-8表達量均顯著高于對照組和高劑量添加組(640 與1 280 mg/kg)(P< 0.05)。在腸道中,與對照組相比,添加FA 會顯著上調Galectin-8的表達量,其中FA 水平為1 280 mg/kg 時表達量最高,并顯著高于其他組(P< 0.05)。

圖6 飼料阿魏酸不同添加水平對杉虎斑肝臟、腸道、頭腎和脾臟的Galectin-8 基因表達量的影響Fig.6 Effects of dietary supplementation of ferulic acid(FA)on the relative mRNA expressions of Galectin-8 in the liver,intestine,head kidney and spleen of hybrid grouper
3 討論
凝集素是先天免疫系統中的重要的體液組成成分,具有一個或多個CRD,可作為模式識別受體(pattern recognition receptors,PRRs),識別潛在致病細菌細胞表面上的病原體相關分子模式(pathogenassociated molecular patterns,PAMPs),在先天免疫和抗病方面起著重要作用[17,29-31]。Galectin-8 具有多種功能,包括促進漿細胞形成、T 細胞和B 細胞增殖、腫瘤調控、凋亡誘導以及免疫調節[17]。本研究描述了杉虎斑Galectin-8的序列特征,檢測了其組織分布,并探究不同FA 水平對其表達量的影響。
人類Galectin-8基因通過可變剪接產生14 種不同的轉錄本,共編碼6 種蛋白質,并且大多數轉錄本具有短的5′UTR 和長的3′UTR[11]。通過RACE 技術獲知杉虎斑Galectin-8基因的cDNA 全長為1 368 bp,5′UTR 為16 bp,3′UTR 為394 bp,共編碼319個氨基酸。半乳糖凝集素家族成員不含信號肽序列,主要定位于細胞質和細胞核,然而它們可通過非經典途徑分泌到細胞外,如直接易位和細胞外囊泡中釋放,具有細胞內和細胞外的功能[9,32]。此外,半乳糖凝集素的特定功能取決于其表達的細胞類型與在細胞或細胞外的定位,因此它們的分泌必須受到嚴格的調控才能正常發揮作用[33]。序列分析表明,杉虎斑Galectin-8 是一種分泌蛋白,但未發現跨膜結構域與信號肽,主要位于細胞質中。目前,半乳糖凝集素分泌的機制尚不清楚,為了對Galectin-8 的分泌有一個全面的了解,還需要進一步的研究。基于結構域分析,發現杉虎斑Galectin-8 屬于串聯重復類型半乳糖凝集素,包含兩個由36 個氨基酸的連接肽所連接的CRD,位于蛋白質序列的兩端。同時,CRD 結構域中有兩個保守的基序,H-NPR 和WGEE,這與其他半乳糖凝集素家族成員一致[11]。翻譯后修飾分析顯示杉虎斑Galectin-8 中有1 個潛在的N-糖基化位點和29 個預測的磷酸化位點。在小鼠的Galectin-8 中已觀察到廣泛的翻譯后修飾,包括糖基化與磷酸化[34]。多重比對結果表明,在Galectin-8中兩個β-半乳糖苷類結合基序在進化中是高度保守的;此外,雖然各物種的連接肽之間無序列同源性,但該區域中的氨基酸數量會導致蛋白質大小的變化[20]。系統進化樹分析顯示,杉虎斑Galectin-8與其他硬骨魚類單獨聚為一支,其中與鞍帶石斑魚的親緣關系最近,而爬行類、兩棲類、哺乳類和鳥類動物分別聚為一支。
在魚類中,已發現半乳糖凝集素在多種組織和細胞中表達[20]。本文中杉虎斑Galectin-8在檢測的組織中均有不同程度的表達,其中在脾臟的表達量最高。這與先前的研究一致,羅非魚和牙鲆的Galectin-8也在脾臟中具有最高的表達量[18,21]。同時在頭腎和肝臟中,杉虎斑Galectin-8也有較高的表達。脾臟、頭腎和肝臟是魚類重要的免疫器官[35]。魚的脾臟也是一個重要的周圍淋巴器官,在抗原捕獲中起著關鍵作用,且與頭腎相比,脾臟是體液免疫反應的主要器官[36]。頭腎類似于哺乳動物的骨髓,具有造血功能,可負責吞噬、抗原處理以及IgM 的形成[37]。盡管脊椎動物的肝臟傳統上被視為代謝、營養儲存和解毒中心,但它還是一個重要的免疫器官,產生細胞因子、趨化因子、補體成分和急性時相蛋白以應對病原體感染[38]。此外,杉虎斑Galectin-8在腦、鰓和腸道中有著相對較高的表達量。腦中的小膠質細胞可積極參與對損傷和致病性感染的快速應答,而鰓和腸道屬于先天免疫系統中的物理屏障[36,39]。綜上所述,這些結果提示Galectin-8可能在杉虎斑的免疫系統中發揮作用[13,18]。
免疫刺激劑是一種能增強防御機制或免疫系統(先天性和適應性)的物質,從而使動物對疾病和外部脅迫有更強的抵抗力[40]。一些研究認為,功能性飼料添加劑可以通過兩種方式激活水生動物的先天免疫系統:(1)直接刺激先天免疫系統;(2)促進共生微生物菌群的生長。飼料添加劑對魚類的免疫刺激作用評估主要集中在非特異性免疫參數上[41]。FA 是一種有效的免疫刺激劑,并且被認為是提高動物免疫力和改善動物生理條件的最重要的添加劑之一,這部分歸因于其抗氧化能力對細胞的保護作用[42-43]。本研究中,與對照組相比,飼料中添加FA 為40-320 mg/kg 時,腸道、肝臟、頭腎和脾臟的Galectin-8表達量顯著上調。類似地,金頭鯛(Sparus aurataL.)攝入含有NEXT ENHANCE?150(由香芹酚和百里香酚組成)的試驗飼料后,Galectin-1在后腸的表達量顯著增加[44];用添加椰棗提取物的飼料喂養歐洲海鱸(Dicentrarchus labrax)4 周后,巖藻糖結合凝集素的表達量顯著上調[45]。半乳糖凝集素轉錄水平的增加,能夠促進微生物的識別和啟動下游免疫反應,如活化T 細胞的增殖、細胞因子的產生、補體激活和白細胞募集[20]。因此,FA 能夠促進Galectin-8基因的表達,這表明杉虎斑的先天免疫得到增強。然而,較高水平的FA(640 與1 280 mg/kg)對Galectin-8的表達量沒有顯著影響。有報道稱,長期補充高濃度的免疫刺激劑可能會導致免疫抑制[46]。此外,Galectin-8 在杉虎斑中的免疫功能尚不清楚。因此需要進一步的研究來闡明杉虎斑Galectin-8 的免疫調節功能,以確定其在防御病原體入侵中的作用。
4 結論
本研究克隆的杉虎斑Galectin-8 屬于串聯重復型半乳糖凝集素,N 端與C 端的CRD 由連接肽所連接,與鞍帶石斑魚具有同源性和高度的一致性,并廣泛表達于各種組織中,其中脾臟的表達量最高。飼料中添加FA 為40-320 mg/kg 時顯著上調腸道、肝臟、頭腎和脾臟的Galectin-8表達量,表明適量添加免疫刺激劑FA 能夠增強杉虎斑的先天性免疫。
