乙草胺處理草莓線粒體膜參數檢測數據集
2023-01-06 06:39:02魯曉峰孫海龍杜國棟徐樹廣
農業大數據學報 2022年3期
關鍵詞:除草劑
魯曉峰 孫海龍* 杜國棟 徐樹廣
(1.中國農業科學院果樹研究所/農業農村部園藝作物種質資源利用重點實驗室,興城 125100;2.沈陽農業大學園藝學院/沈陽市北方果樹栽培與生理生態重點實驗室,沈陽 110866)

數據庫(集)基本信息匯總表http://www.agridata.cn/data.html
1 引言
乙草胺是選擇性芽前處理除草劑,通過進入作物根圍土壤,經胚芽鞘或下胚軸吸收,吸收后向上傳導,阻礙蛋白質合成而抑制細胞生長,使雜草幼芽、幼根生長停止,通常通過觀察植物根系生理功能的影響,表征除草劑脅迫對根系發育的傷害程度[1]。Ghazi-Khansari M 等研究發現,利用草甘膦處理后,毛桃根系呈現出顏色變深、須根變短、變少等現象,說明除草劑脅迫對毛桃根系生長發育有較大傷害[2]。根系的活力直接影響植株整體的生長發育,是植株幼苗發育過程中的重要生理指標。李亞東等研究發現,高濃度乙氧氟草醚、二甲戊靈、仲丁靈明顯抑制棉花的根系活力,而五氟磺草胺即使低濃度時,對棉花的根系活力也表現出嚴重的抑制效果[3]。在正常生理狀態下,植物組織中的ROS 產生與清除處于平衡狀態[4]。當植物受到逆境脅迫時,植株體內會產生過量ROS,引起細胞膜系統受到損傷,引起膜脂過氧化物產物(MDA)激增[5-6]。土壤中除草劑西草凈的含量達到0.8~8.0 mg kg-1時,小麥根系中活性氧(ROS)大量產生,對根系細胞膜脂造成損傷[7]。且研究指出,外源施加除草劑丙酯草醚和丁草胺,均能不同程度破壞小麥、油菜和大麥體內的保護性酶活性,使植株體內的膜質過氧化產物MDA 含量提高[8-9]。溫銀元等人研究發現,受到除草劑撲草凈脅迫后,西府海棠幼苗根系中抗氧化酶SOD、POD 等含量均顯著升高,O2-、H2O2含量顯著高于對照[10];馮煜在研究除草劑谷友和苯·唑·2 甲鈉脅迫下糜子根系抗氧化酶活性變化時也發現,兩種除草劑均對糜子根系產生藥害,降低糜子根系SOD及POD活性,顯著提高MDA含量[11]。
線粒體是根系細胞主要的細胞器之一,是感受外界脅迫相關信號主要部位[12-13]。研究表明,逆境脅迫會導致線粒體的結構和功能發生明顯變化[14]。如鹽堿脅迫下,番茄根系線粒體內H2O2和MDA 含量增加,線粒體膜通透性增大[15];干旱脅迫條件下,銀沙槐幼苗葉片中線粒體膜完整性低,出現內部嵴消失、空泡化等現象[16];鎘脅迫導致平邑甜茶根系細胞線粒體電子傳遞受阻,氧自由基含量增加,造成線粒體膜質過氧化,細胞發生程序性死亡[17]。同時,線粒體也是產生ROS 的場所,其內含有的抗氧化酶對機體有一定保護作用[18]。過量的ROS 會引起線粒體膜氧化損傷,導致線粒體的電子傳遞受阻,內膜的完整性和呼吸電子傳遞鏈受到破壞,進一步破壞線粒體的抗氧化防御體系,使線粒體O2-產生速率加快、H2O2和MDA含量增高、膜脂過氧化加劇[19]。在植物根系中,線粒體呼吸鏈電子漏是O2-產生的主要來源[20],而線粒體膜電位的下降又能導致線粒體呼吸鏈電子漏的增加[21]。細胞色素Cyt c 介導的呼吸鏈電子漏旁路,可以清除由于呼吸鏈漏電子而產生的O2-和H2O2。如在外源丁香酸和鄰苯二甲酸脅迫下,根系線粒體膜透性增大,降低膜電位及根系線粒體內膜上細胞色素Cyt c/a 含量,線粒體內膜細胞完整性遭到破壞,電子傳遞受阻[22]。徐建興研究也發現,當植物遭受逆境脅迫時,線粒體內膜的完整性和呼吸電子傳遞鏈受到破壞,最終導致線粒體O2-產生速率加快,H2O2和MDA 含量增高[23]。目前,雖然有關除草劑影響作物生長發育的研究較多,但對作物根系線粒體膜結構和功能的損傷機制尚未見詳細報道。本文以草莓植株為試驗材料,研究乙草胺誘發ROS 對草莓根系線粒體膜電位、膜通透性等生物膜結構和功能的影響,為生產上有效應對乙草胺類除草劑脅迫提供借鑒。
2 數據采集與處理方法
本試驗于2017年5—9月在沈陽農業大學苗木良種繁育基地進行,供試材料為日本草莓品種‘寶交早生’(Fragaria×ananassaDuch.),取自遼寧省沈陽市沈北新區國邦園藝農場草莓苗木生產基地。濃度99%乙草胺乳油購自吉林美聯化學品有限公司。
將‘三葉一心’的草莓假植幼苗定植于內徑寬和高分別為16.0 cm和16.0 cm的塑料盆缽中,栽培基質為園土∶泥炭土∶有機肥=3∶1∶1(按體積比均勻混合),每盆裝入基質1.25 kg,栽植草莓苗1 株,重復30次。試驗設乙草胺處理(處理D,乙草胺濃度為使用推薦劑量的10倍,每缽添加濃度0.2 mol·L-1的乙草胺量100 ml)及對照(CK,未施加乙草胺)處理。草莓植株生長30 d 后,選擇長勢基本一致的草莓植株,按試驗設定劑量將除草劑溶液澆灌于盆缽中,對照施入等量的蒸餾水,各處理重復5次,分別于處理后0 d、1 d、3 d、5 d、7 d和9 d,取草莓根系前端1~2 cm分生區部位根段樣品,用于各生理指標的測定。
線粒體的提取參照楊玖英[24]的方法;線粒體膜電位測定參照Braidot[25]等方法;線粒體膜通透性的測定參照金超芳[26]等方法;線粒體細胞色素c/a(Cyt c/a)測定參照Tonshin 及馬懷宇[27]等方法。丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法測定[28];過氧化氫酶(CAT)活性采用KMnO4滴定法測定[29];超氧化物歧化酶(SOD)活性采用氮藍四唑(NBT)顯色法測定[30];過氧化物酶(POD)活性采用愈創木酚法測定[31];超氧陰離子(O2-)活性采用羥胺法測定[32];過氧化氫(H2O2)含量采用四氯化鈦法測定[33]。根系活力采用TTC 法測定[34],單位為mg·g-1· h-1;質膜H+-ATPase 測定參照Blumwald 等[35]方法,單位為μmolPi mg-1protein · h-1。采 用Excel 2010 軟件進行原始 數據處理,Sigma Plot 10.0軟件繪圖。
3 數據樣本描述
3.1 乙草胺處理對草莓根系線粒體膜功能的影響
施加乙草胺后,根系線粒體膜Δψm 呈持續下降的趨勢,于處理5 d 下降幅度最明顯,較對照下降64.3%,處理后期無明顯變化(圖1-a)。由圖1-b 可知,與對照相比,前期草莓根系線粒體內膜上細胞色素Cyt c/a 含量明顯下降,于處理7 d 后達最低值,后期的線粒體Cyt c/a 變化不明顯。乙草胺處理一定程度上影響了線粒體功能的發揮,與對照相比,在乙草胺處理5 d 后,根系線粒體膜透性(MPT)明顯上升,比對照上升138.0%,處理9 d 后,線粒體膜的MPT 達最高值。
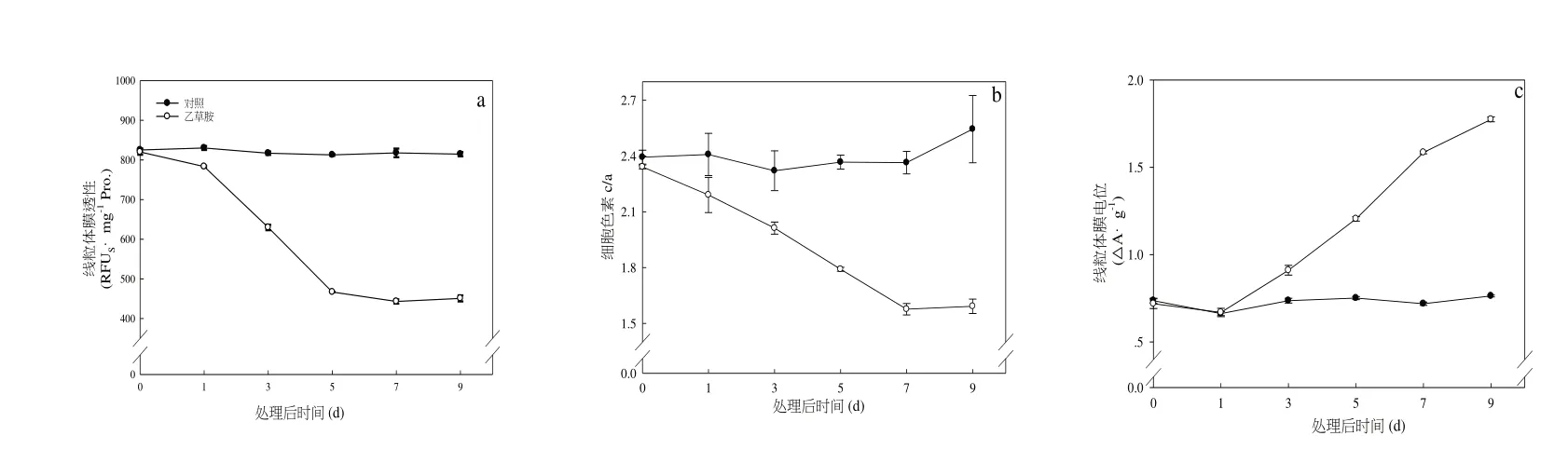
圖1 乙草胺對草莓根系線粒體功能的影響Fig.1 Effects of acetochlor on the mitochondrial function of strawberry roots
3.2 乙草胺處理對草莓根系活力、H+ -ATPase及丙二醛含量的影響
由圖2-a 可知,與對照相比,乙草胺處理的草莓根系活力呈現直線下降的趨勢。其中,乙草胺處理9 d 后,根系活力僅分別為對照的54.8%。草莓根系細胞質膜H+-ATPase 活性均呈現明顯下降趨勢(2-b)。丙二醛(MDA)是植物受到逆境脅迫所生成膜質過氧化物質,其含量常常用來表征植物機體內膜質過氧化程度。由圖2-c 可以看出,與對照相比,在除草劑乙草胺脅迫下,根系MDA 含量均呈現持續下降趨勢。
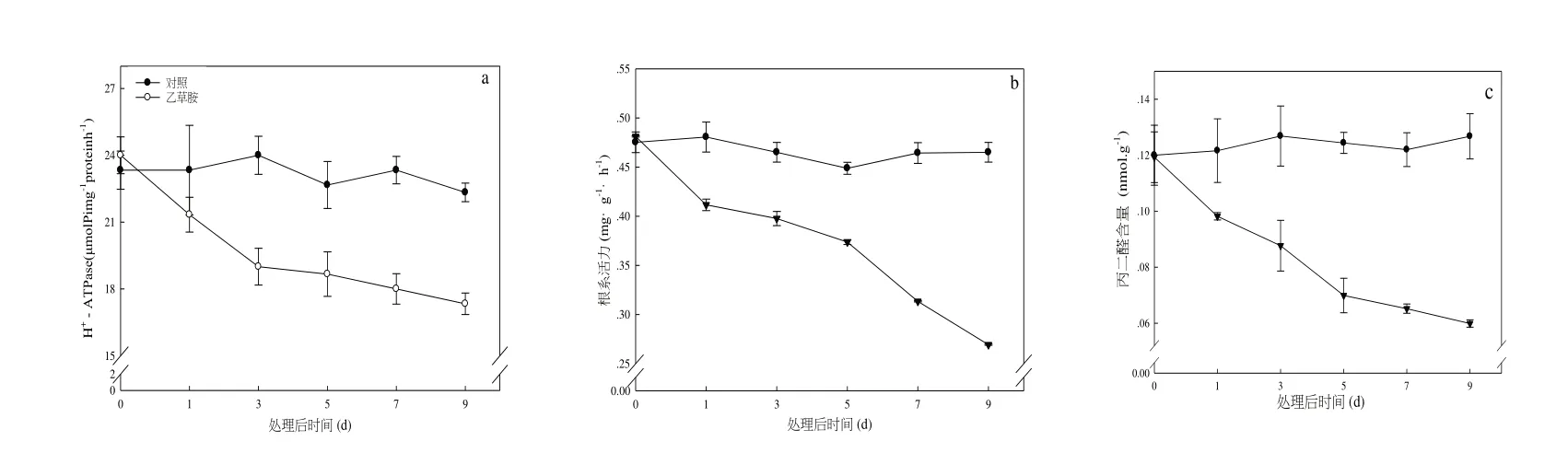
圖2 乙草胺對草莓根系活力、H+-ATPase及丙二醛含量的影響Fig 2 Effects of acetochlor phytotoxicity on root activities,H+-ATPase and MDA in strawberry roots
3.3 乙草胺處理對草莓根系活性氧含量的影響
如圖3-a 所示,隨著處理時間延長,乙草胺處理后草莓根系O2-含量呈逐漸上升趨勢,處理第5 d 后,含量較對照增長14.9%,處理9 d后O2-含量達到最高值。由圖3-b 可知,與對照相比,隨著脅迫時間的持續,乙草胺處理后根系H2O2含量呈現上升趨勢,處理7 d后,H2O2含量達到最高值,較對照增長32.5%。
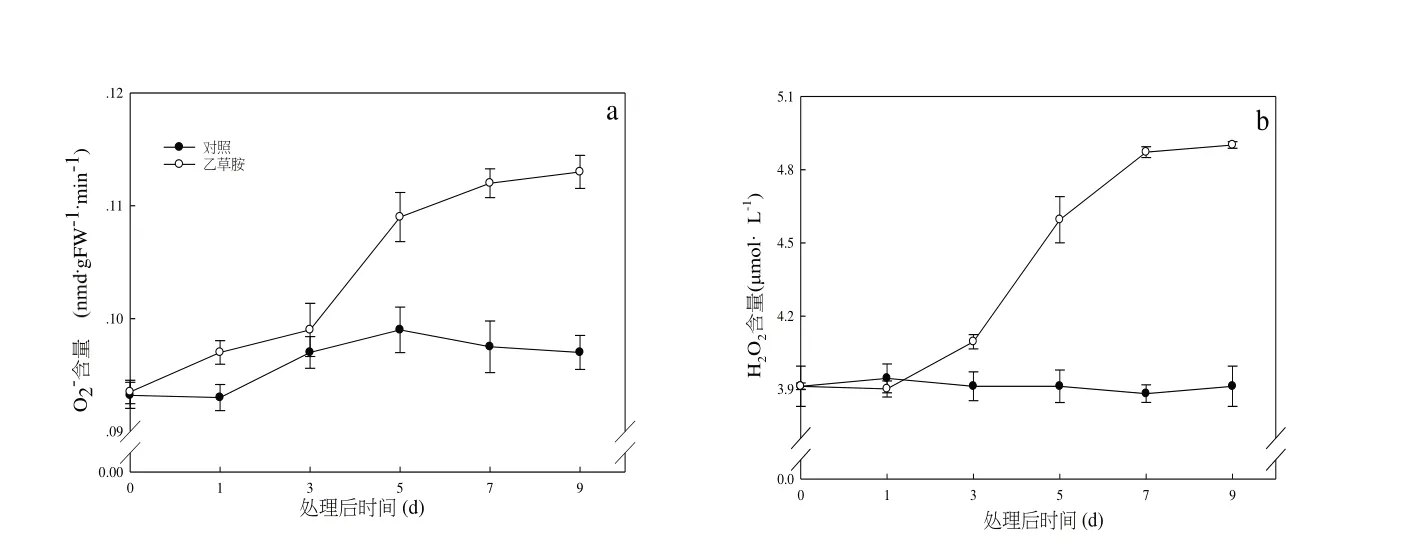
圖3 乙草胺對草莓根系ROS含量及H2O2含量的影響Fig.3 Effects of acetochlor on reactive oxygen species and hydrogen peroxide contents of strawberry roots
3.4 乙草胺處理對草莓根系保護性酶活性的影響
由圖4-a 可以看出,隨著除草劑脅迫處理時間增加,草莓根系SOD 活性均呈逐漸上升趨勢。其中,在脅迫處理9 d后,根系SOD活性比對照增長32.8%。如圖4-b 可知,隨乙草胺脅迫處理時間的增加,根系POD 活性呈現上升趨勢,于處理9 d 后達最高水平。隨著除草劑脅迫程度加重,根系CAT 活性呈先升高后降低的趨勢,于脅迫處理3 d 后達最高水平,隨后CAT 活性迅速降低,處理9 d 后的CAT 活性達最低值,較對照降低了17.1%(圖4-c)。

圖4 乙草胺對草莓根系保護性酶活性的影響Fig.4 Effects of acetochlor on protective enzyme activity of strawberry roots
4 數據質量控制與驗證/評估
整個生長周期在基地溫室內進行,定期澆灌Hoagland 營養液,補充充足的水分及養分,確保植株正常生長。草莓植株生長30 d后,選擇長勢基本一致的草莓植株,按試驗設定劑量將除草劑溶液澆灌于盆缽中,對照施入等量的蒸餾水,各處理重復5 次,為防止溶液流失,在培養缽下放置托盤,將外滲的溶液重新澆回培養缽中。分別于處理后0 d、1 d、3 d、5 d、7 d和9 d,取草莓根系前端1~2 cm 分生區部位根段樣品,用于各生理指標的測定。取樣前制定標準統一的方案,由1 人獨立完成,保證選取樣品的固定性。測定各指標前對所有參與實驗操作的人員進行集中培訓,盡可能保證參與人員的固定性,減少人為誤差。
5 數據價值與使用建議
本研究公開發表了2017 年乙草胺處理草莓線粒體膜參數檢測數據集,內容包括了草莓根系線粒體膜電位、細胞色素Cyt c/a、膜透性、根系活力及H+-ATPase、丙二醛及活性氧含量等數據,該數據可結合線粒體細胞器表觀變化,進一步確定除草劑脅迫對線粒體結構影響的具體作用位點,可進行分子標記和相關基因挖掘,也可結合其他除草劑對根系線粒體作用程度,篩選出更優質的除草劑,同時也可為生產上有效應對乙草胺類除草劑脅迫提供參考。本數據集原始數據可通過國家農業科學數據中心獲取,登錄首頁后點“資源服務”下的“數據服務”,進入相應頁面。
數據作者分工職責
魯曉峰(1992—),女,遼寧人,碩士,助理研究員,研究方向為李杏資源與育種研究。主要承擔工作:田間調查、數據處理、質量控制及論文撰寫。
孫海龍(1988—),男,黑龍江人,碩士,助理研究員,研究方向為李杏資源與育種研究。主要承擔工作:方案設計、組織實施、論文構架及質量控制。
杜國棟(1971—),男,遼寧人,博士,教授,研究方向為果樹栽培與生理生態。主要承擔工作:方案設計、組織實施、論文構架及質量控制。
徐樹廣(1973—),男,遼寧人,農藝師,研究方向為李杏資源與育種研究。主要承擔工作:田間調查及質量控制。
猜你喜歡
世界農藥(2019年3期)2019-09-10 07:04:10
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
長江蔬菜(2016年10期)2016-12-01 03:05:27
獸醫導刊(2016年12期)2016-05-17 03:51:29
現代農業(2016年5期)2016-02-28 18:42:36
雜草學報(2015年2期)2016-01-04 14:58:05
種業導刊(2016年9期)2016-01-03 01:27:14
營銷界(2015年23期)2015-02-28 22:06:18
營銷界(2015年22期)2015-02-28 22:05:11
