蘋果愈傷組織CRISPR/LbCpf1基因編輯體系的建立
2023-01-06 13:07:08劉楊張春玲肖旭由春香王小非
落葉果樹 2022年6期
關鍵詞:效率
劉楊,張春玲,肖旭,由春香,王小非
(山東農業大學園藝科學與工程學院/作物生物學國家重點實驗室/國家蘋果工程技術研究中心,山東泰安 271018)
CRISPR(Clustered Regularly Interspaced Short Palindromic Repeats)系統,因其高效、精確和便捷性,被廣泛應用于植物基因組基因編輯。CRISPR/Cas9(CRISPR-associated 9)系統已在多種園藝植物中開展研究,例如柑桔[1]、番茄[2]、黃瓜[3]、蘋果[4, 5]、葡萄[5, 6]、獼猴桃[7]和山藥[8]等。CRISPR/Cpf1(CRISPR fromPrevotellaandFrancisella1)是CRISPR/Cas9系統的一類衍生體,AsCpf1(Acidaminococcussp. BV3L6)、FnCpf1(Francisellatularensissubsp.novicidainU112)以及LbCpf1((Lachnospiraceae bacterium ND2006)是常用的三種Cpf1蛋白[9-11]。相對于Cas9對5’-NGG-3’ PAM(Protospacer adjacent motif)序列的偏好性,CRISPR/Cpf1傾向于富含T堿基的PAM序列并在PAM序列遠端造成雙鏈斷裂,形成粘性末端缺口,具有突變效率高、多基因編輯元件構建更簡易且脫靶率更低等優勢,極大拓展了CRISPR/Cas系統應用范圍[12, 13]。CRISPR/Cpf1已在水稻[14]、煙草[15]、柑桔[16]以及楊樹[17]等植物中實現基因編輯,但蘋果(Malus×domestic)中的成功應用報道較少。
相對于蘋果組培苗,轉基因愈傷組織更容易獲得,是驗證基因編輯工具效率以及基因功能的有效工具。實驗室前期發現,轉錄因子MdMYB1(MDP0000259614)基因正調控花青苷合成途徑,其突變體使王林愈傷組織花青苷積累受阻[18, 19]。因此,筆者以MdMYB1基因作為靶點,構建雙靶點的CRISPR/LbCpf1載體并轉入王林愈傷組織,分析基因編輯效率,以期在蘋果愈傷組織中建立高效的基因編輯體系,進而為蘋果分子育種研究提供參考。
1 材料與方法
1.1 試驗材料
王林蘋果(Orin)愈傷由實驗室保存,于1.5 mg/L 2,4-D和0.4 mg/L 6-BA的MS固體培養基中培養。25 ℃溫室中黑暗條件放置,每15 d繼代培養。轉基因愈傷在含有30 mg/L潮霉素的繼代培養基上每15 d繼代培養。
1.2 靶點選擇
根據實驗室前期MDP0000259614的序列在CRISPR-P 2.0網站(http://crispr.hzau.edu.cn/CRISPR2/)進行查詢,選擇評分低,即預測脫靶率低,GC含量在40%~60%,長度為22~24 bp,PAM序列為5’-NTTT-3’,位置位于編碼序列區的靶點1(Target1)(5’-CCTCTCCATGAATCTCAACGCACT-3’)與靶點2(Target2)(5’-GGGCTGAGGTCTTATCACATTGGT-3’),序列在基因組位置見圖1。

圖1 MdMYB1基因的靶點位置示意圖
1.3 載體構建
CRISPR/LbCpf1多基因編輯載體由朱健康團隊贈送[20]。根據載體圖譜,利用SnapGene軟件進行載體設計,將包含BsaI酶切位點、靶點序列以及crRNA的基因片段通過設計長片段引物LbTarget1-F/LbTarget2-R進行PCR擴增,將目的大小的單一明亮條帶進行膠回收,膠回收具體方法參見DNA膠回收試劑盒(諾唯贊,南京)。將載體與片段進行BsaI(New England Biolabs)雙酶切,反應體系為50 μL,包含5 μL的10× NEB Buffer、1 μg DNA以及1 μL的BsaI酶,反應時間為1 h,進行凝膠電泳后回收。最后通過T4連接酶(Takara,大連)將載體與插入片段連接,流程見圖2。連接產物轉化Trans 5α(全式金,北京),利用相應引物U6-F/Cpf1-R的PCR反應鑒定出陽性菌落,獲得MdMYB1雙靶點CRISPR/Cpf1基因編輯載體——MYB1-LbCpf1。
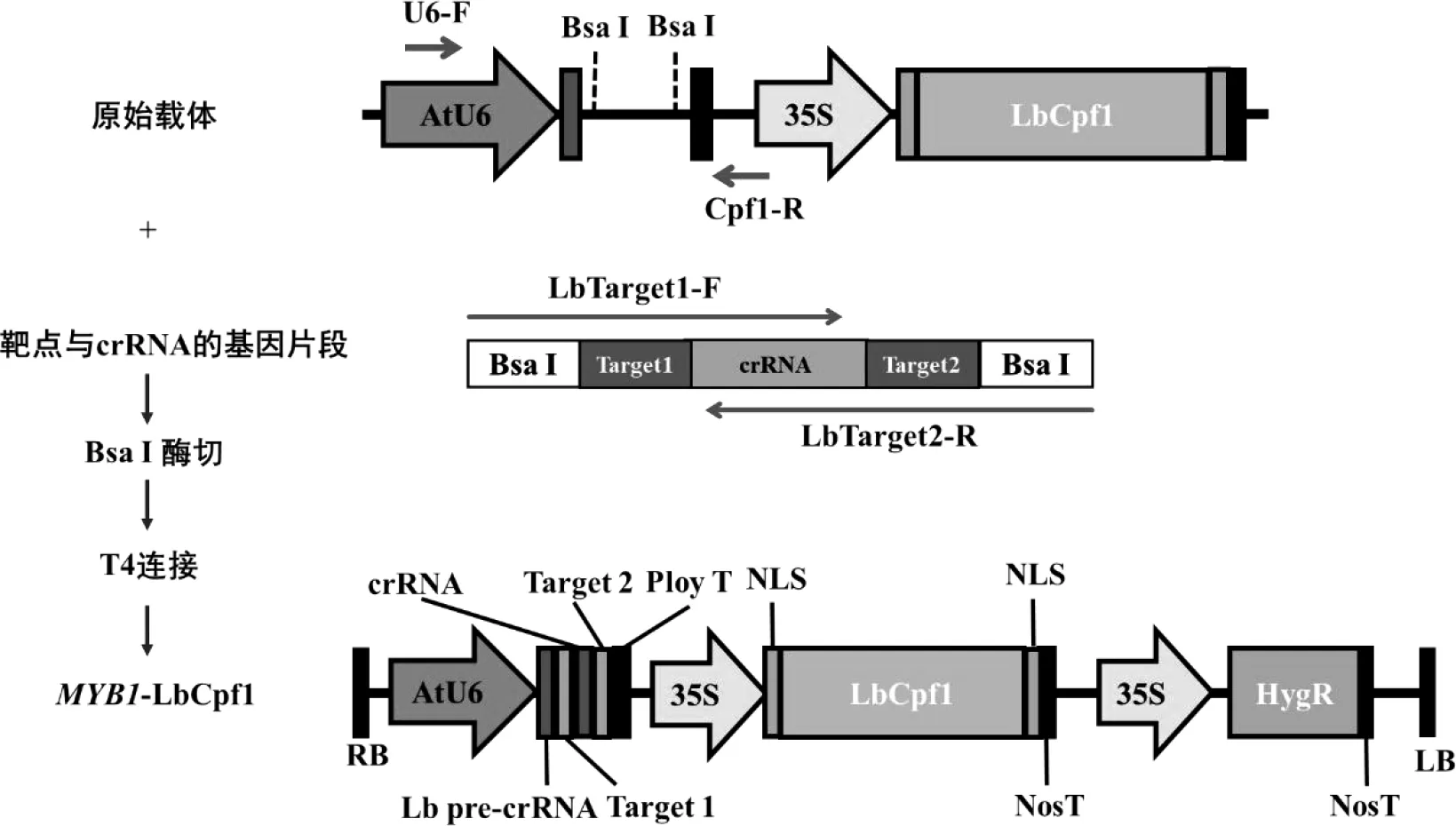
圖2 載體構建流程圖

表1 所用引物
1.4 王林愈傷侵染
載體轉化EHA105農桿菌(全式金,北京),陽性菌落于50 mg/L卡那霉素與10 mg/L利福平LB培養基中培養至OD600=1.0。5 000 rpm收集菌體并用滅菌ddH2O重懸至OD600=0.8。將15 d長勢良好的王林愈傷組織壓碎浸泡于上述重懸液中,室溫下避光振蕩15 min,撈出并于濾紙上吸干懸浮液,鋪至無抗生素的愈傷培養基。將24 ℃恒溫培養箱中暗培養2 d的愈傷組織取出,均勻平鋪于含有30 mg/L潮霉素和250 mg/L頭孢霉素的愈傷組織培養基中,恒溫箱中暗培養30~45 d,將新生的陽性愈傷組織繼代在含抗生素培養基中,待3~4輪后取樣提取DNA,利用引物進行鑒定。轉基因率(%)=轉基因愈傷株系/陽性愈傷株系×100。
1.5 基因編輯事件檢測
對Target1與Target2 DNA區域分別設計引物MdMYB1-F/R與MdMYB1-F1/R1。試劑盒(康為世紀,北京)法提取陽性愈傷組織DNA,利用Phanta EVO高保真DNA聚合酶(諾唯贊,南京)對包含相應靶點的DNA序列進行擴增,反應體系為50 μL,包含5× EVO Buffer 10 μL、dNTP Mix 1 μL、10 μM引物各1 μL、愈傷組織DNA 100 ng以及1 μL酶,通過95 ℃ 3 min;95 ℃ 15 s,58 ℃ 15 s,72 ℃ 30 s,32個循環;72 ℃ 7 min;4 ℃保溫。對PCR產物進行凝膠電泳觀察,回收純化,連接至克隆載體pEASY?-Blunt Simple Cloning Kit(全式金,北京)并轉化大腸桿菌Trans 5α。每個株系挑選8個陽性菌落,送至青島擎科梓熙生物技術有限公司進行測序,利用SnapGene軟件進行序列比對并統計。基因編輯效率(%)= 基因編輯事件發生愈傷株系/總轉基因株系×100。
1.6 花青苷積累試驗
將15 d生長狀態一致的野生型與轉基因愈傷組織鋪在不含氮的培養基中預培養10 d,移至強光低溫培養箱(光子通量密度約為100 μmol/s·m2,溫度為19 ℃)中培養,觀察著色情況。
2 結果與分析
2.1 轉基因愈傷的獲得與鑒定
將圖2中載體轉入王林蘋果愈傷組織中,35 d后篩選培養基中有明顯新生的愈傷組織,抗性培養基中繼代3次獲得陽性株系,其中MYB1-LbCpf1載體共獲得10個株系,命名為LbCpf1-T1-T2#1至#10。將繼代3次的愈傷組織提取DNA,利用潮霉素標簽引物進行PCR檢測T-DNA插入情況,結果發現野生型愈傷(WL)中出現假陽性條帶。于是利用U6-F/Cpf1-R引物檢測,結果如圖3所示,雖然WL中有雜帶產生,但與陽性對照可顯著區分,10個MYB1-LbCpf1陽性愈傷組織中有9個含有T-DNA插入區段,表明MYB1-LbCpf1載體的T-DNA片段成功插入轉基因愈傷基因組中。
2.2 基因編輯事件檢測
提取轉基因愈傷組織DNA,利用Target1和Target2區域對應引物進行擴增,將目的片段連接克隆載體并轉化大腸桿菌,每個株系的每個靶點挑取陽性單克隆8個送至測序。測序結果表明,9個轉基因植株中共有2個株系發生Target1位置的突變,突變效率為22.2%,共有3個株系發生Target2位置的突變,突變效率為33.3%,由于#3在兩靶點均發生突變,因此CRISPR/Cpf1雙靶點載體造成的總突變率為44.4%(表2)。在發生基因編輯事件的株系送測的所有克隆中,Target1有4個克隆序列發生突變,占16個克隆的25%;Target2有16個克隆發生突變,占25個克隆的64%。Target1的4個突變類型中,3/4為小片段缺失型突變,缺失堿基數為1 bp、5 bp和7 bp,發生突變株系為#1與#3,1/4為7 bp的替換型突變,發生株系為#3,其中#3發生突變頻率最高。而Target2的16個突變類型中,13/16為2~10 bp的小片段缺失型突變,發生株系為#2、#3與#5;1/8為20 bp與94 bp的大片段缺失型突變,發生株系為#3與#5;1/16為2 bp的替換型突變,發生株系為#2。上述Target2所有突變類型中,#2發生頻次最高,而#3次之(圖4)。以上結果表明,本研究中CRISPR/Cpf1引導的雙靶點基因編輯在Target2位置有更高的效率。

圖3 陽性愈傷組織獲得以及T-DNA插入片段的PCR擴增產物電泳

表2 CRISPR/LbCpf1在王林愈傷中的靶點突變率
2.3 CRISPR/Cpf1引導的基因編輯特征分析
統計Target2發生的所有基因編輯事件,分析所有的突變類型發現,CRISPR/Cpf1在蘋果基因組中引導的堿基刪除事件中,7 bp的小片段刪除為發生頻率最高的事件(25%),8 bp的刪除次之(18.8%),而9 bp與2 bp的刪除發生頻率相仿(12.5%),以上高頻率的堿基刪除事件占比68.8%。較大片段(>20 bp)刪除的事件發生頻率較低,為6.3%。分析發生突變事件的堿基位置發現,距PAM序列14~22 bp的堿基發生突變頻率(6.38%~10.64%)高,發生突變頻率(10.64%)最高的位置距離PAM序列17 bp。而PAM序列附近發生突變頻率(0.71%)最低(圖5)。以上結果說明,本研究中CRISPR/Cpf1系統在蘋果基因組中傾向于引導小片段堿基刪除,堿基突變位置多于PAM序列遠端14~22 bp位置。

圖4 CRISPR/LbCpf1在蘋果愈傷組織基因組不同靶點位置的編輯事件統計
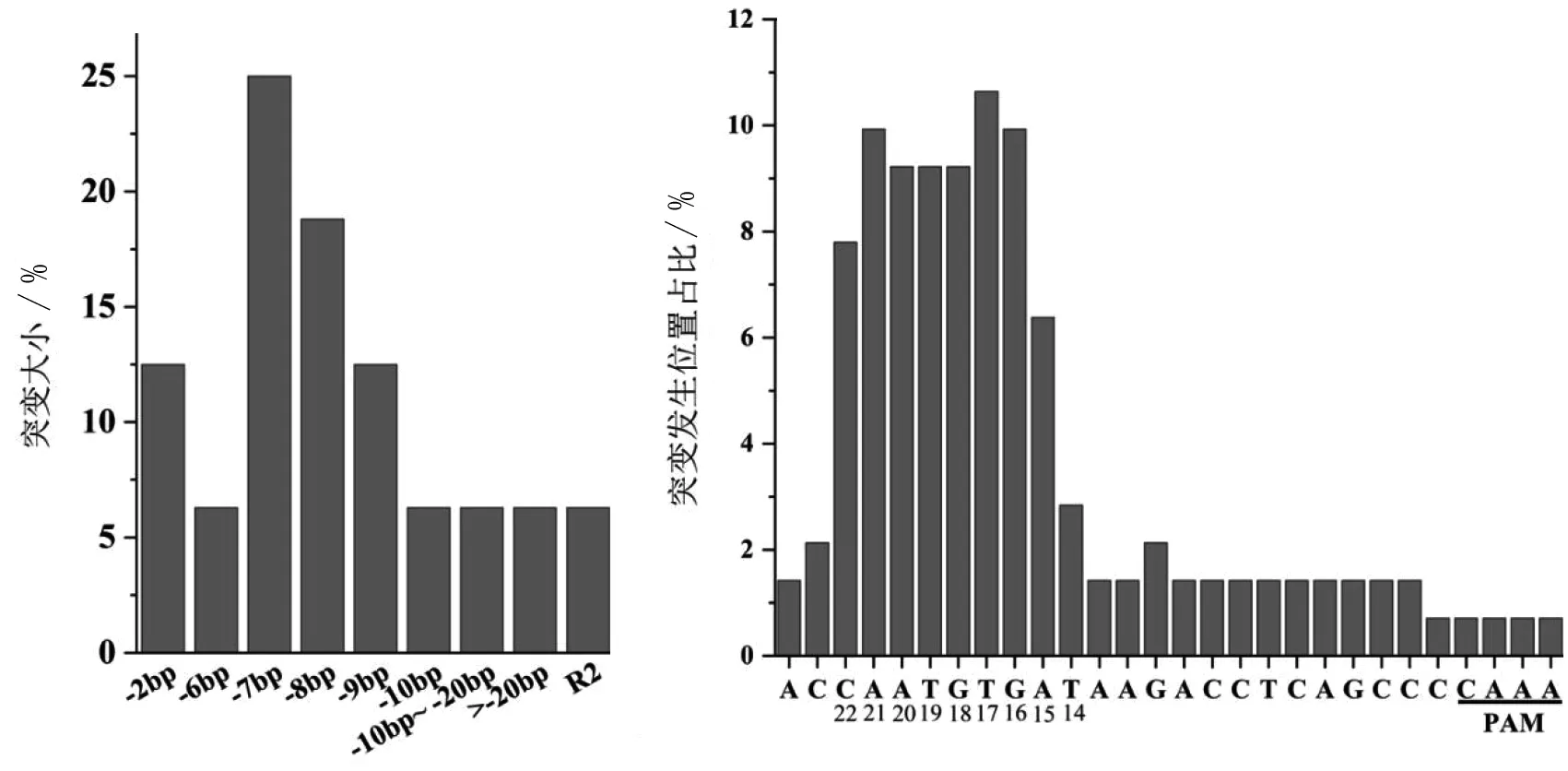
圖5 CRISPR/LbCpf1在蘋果愈傷組織中的基因編輯特征分析
2.4 MdMYB1基因編輯愈傷的表型驗證
為證實利用CRISPR/Cpf1基因編輯愈傷進行功能研究的可行性,選定生長狀態一致的野生型與發生基因編輯事件頻率較高(#2、#3和#5)的愈傷組織進行花青苷積累試驗。結果表明,經過12 d的不含氮誘導以及低溫培養,野生型王林愈傷組織已經有少量著色,但3個基因編輯愈傷都沒有著色(圖6)。限于王林愈傷組織的遺傳型特征,經過12 d加7 d的花青苷誘導處理后沒有更多花青苷形成,而基因編輯愈傷花青苷積累量始終沒有積累花青苷(圖中未展示)。以上結果說明,MdMYB1的CRISPR/Cpf1基因編輯愈傷抑制花青苷積累。

圖6 基因編輯愈傷花青苷積累試驗
3 小結與討論
為實現CRISPR/Cas9系統的多基因編輯,多轉錄單元提供了解決思路,但多個啟動子與sgRNA的組合使得載體長度過大且構建過程繁瑣[7, 21, 22]。而由于CRISPR/Cpf1所需sgRNA長度較短,且Cpf1蛋白的RNA酶活性可將單個sgRNA從串聯的CRISPR RNA中釋放,因此CRISPR/Cpf1的多基因編輯體系相對于CRISPR/Cas9系統更高效,組裝更簡便[15]。本研究所用多基因編輯CRISPR/Cpf1載體構建簡易,sgRNA支架僅19 bp,通過串聯sgRNA支架與sgRNA即可構建出單轉錄單元的多基因編輯表達盒,可極大降低蘋果中多基因編輯的成本。蘋果愈傷組織轉化效率較高,獲得轉基因愈傷周期短,適于對基因編輯載體效率以及基因功能的驗證。本研究在較短時間內獲得轉基因愈傷,轉化率約90%,較高的轉化率使CRISPR/Cpf1的基因編輯效率(44%)也較高,說明蘋果愈傷是驗證基因編輯效率的高效工具。基因編輯愈傷表現出MdMYB1突變體的性狀,說明利用CRISPR/Cpf1體系對基因進行功能研究是可行的。
在MdMYB1基因的編碼區分別設計了兩個靶點,兩靶點距離較遠,設計一對引物無法有效地擴增以檢測突變情況。因此不能排除Cpf1蛋白在兩靶點上共同發揮功能的情況,測序結果中也發現自靶點1后信號混亂,以及無法讀出的情況,或許能說明兩靶點間發生大片段刪除事件。靶點2相對靶點1有著更高的突變率(33.3% >22.2%),說明靶點2的sgRNA對于靶標基因有著更高的結合效率,而靶點2中出現較高頻率的小片段刪除也說明sgRNA的高結合率使Cpf1蛋白在靶點2位置有著更高的切割效率。sgRNA選擇影響CRISPR/Cas系統效率[23]。而靶點1 sgRNA 3’端較高的GC含量可能是其相對較低的突變率的原因之一[24]。因此,對于蘋果中基因編輯高效的靶點的選擇應同時結合軟件篩選以及轉化愈傷組織驗證。統計所有株系的突變類型發現,CRISPR/Cpf1傾向于7~9 bp的小片段刪除,且突變位置集中在PAM序列遠端的14~22 bp,符合典型的CRISPR/LbCpf1體系的基因編輯特征[9, 25]。發生突變的幾個株系中出現了較多類型的突變情況,說明在繼代過程中,Cpf1可能持續發揮功能對靶點進行切割。鑒于愈傷組織可能為單細胞或多細胞起源,所以基因編輯愈傷可能為雜合體或純合體,因此呈現了多種的突變類型。如何獲得單細胞起源的純合體基因編輯愈傷是亟待解決的問題。
基因編輯效率受多種因素影響,包括Cas蛋白、載體遞送效率以及基因編輯元件的表達效率[26, 27]。通過優化載體中基因編輯元件的啟動子可以提高基因編輯效率[28],蘋果中通過應用MdU3啟動子顯著提高MdPDS基因的編輯效率[29]。因此,為提高CRISPR/Cpf1載體的效率,可將sgRNA啟動子替換為蘋果MdU6/MdU3啟動子。sgRNAs的GC含量高于50%有相對較高的編輯效率[21],葡萄中發現GC含量高于60%時顯著提高基因編輯效率。通過挑選較高GC含量的sgRNAs也是提高CRISPR/Cpf1載體的舉措之一。
猜你喜歡
瘋狂英語·初中天地(2021年5期)2021-07-21 02:24:28
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數理化(高中版.高考數學)(2020年5期)2020-06-02 09:19:08
商周刊(2017年9期)2017-08-22 02:57:49
遼寧經濟(2017年6期)2017-07-12 09:27:16
中國衛生(2016年9期)2016-11-12 13:27:54
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國洗滌用品工業(2015年7期)2015-02-28 19:02:38
電子設計工程(2015年12期)2015-02-27 12:06:10
中國衛生(2014年11期)2014-11-12 13:11:32
