南大洋浮游細菌群落結構研究進展
2023-01-07 08:16:04孫玉蓉馬玉欣曹叔楠羅光富藍木盛何劍鋒
極地研究 2022年4期
孫玉蓉 馬玉欣 曹叔楠 羅光富 藍木盛 何劍鋒,4,5
研究進展
南大洋浮游細菌群落結構研究進展
孫玉蓉1,2馬玉欣3,4,1曹叔楠2,3羅光富2,3藍木盛2,3何劍鋒2,3,4,5
(1上海海洋大學海洋科學學院, 上海 201306;2南極長城極地生態國家科學觀測研究站, 中國極地研究中心, 上海 200136;3自然資源部極地科學重點實驗室, 中國極地研究中心, 上海 200136;4上海交通大學海洋學院, 上海 200030;5同濟大學環境科學與工程學院, 上海 200092)
浮游細菌是海洋微食物環的重要組成部分, 在維持海洋生態系統穩定和物質循環中起著重要作用。本文對南大洋浮游細菌群落組成和分布特征以及影響因素進行概述, 結果顯示, 優勢類群為α-變形菌綱、γ-變形菌綱和噬纖維菌-屈撓桿菌-擬桿菌群, 非優勢類群組成則存在顯著的空間差異。浮游細菌群落存在明顯的季節變化, 溫度和溶解有機碳濃度是調控群落結構的主要環境因子, 浮游細菌與微食物環其他類群存在明顯的相互作用。氣候變化導致的海洋升溫、海洋酸化等環境變化會對浮游細菌產生深遠影響。未來應加強對極地上層海洋與海冰相關浮游細菌群落和深海浮游細菌群落的研究, 依托站基開展季節變化的現場調查和模式研究, 預測南大洋浮游細菌群落的潛在變化趨勢及調控機理。
南大洋 浮游細菌 群落組成 季節變化 環境因子
0 引言
海洋浮游細菌是海洋微食物環的重要組成部分, 具有極強的有機物攝入能力[1], 是海洋物質循環的重要一環[2-4], 而碳、氮等物質的生物地球化學循環在維持海洋生態系統穩定和影響全球氣候變化中起著重要作用[5-7]。對南大洋的研究同樣顯示, 微食物環是南極海域碳和能量流動的重要途徑[8]。
海洋浮游細菌群落中, 變形菌門的a-變形菌綱和γ-變形菌綱在生物量和多樣性上均占優勢, 而擬桿菌門也具有較高的生物多樣性[9-10]。南大洋浮游細菌、特別是海洋表層浮游細菌群落, 具有明顯的區域特異性[11]。盡管相較于其他海域, 南大洋和北冰洋的浮游細菌群落結構較為相近[12], 但有研究表明, 南大洋表層特有分類單元(Operational Taxonomic Units, OTUs)比例高達78%[13]。
近年來, 隨著南大洋局域升溫、混合加劇、海冰覆蓋面積波動和淡水輸入增加等環境變化, 南大洋生態系統得到了更多的關注, 對浮游細菌群落組成、空間分布、季節變化和環境調控機理等也進行了更多的研究[14-17], 但與低緯度海域的研究強度相比仍有較大的差距。我國對南大洋浮游細菌的研究極為有限[17-19]。本文對南大洋浮游細菌群落結構的最新研究進行總結, 分析其時空分布和季節變化特征, 并對今后該領域的研究進行了展望, 以期可以推進我國的相關研究。
1 南大洋浮游細菌優勢類群
南大洋上表層海域浮游細菌的優勢類群為α-變形菌綱、γ-變形菌綱和噬纖維菌-屈撓桿菌-擬桿菌群(Cytophaga-Flexibacter-Bacteroides, CFB), 中深層優勢類群則以γ-變形菌綱為主[20-21], 不同海域優勢類群詳見表1。其中,a-變形菌綱又以SAR11類群占優[22]。該類群在開闊大洋和近岸水域都具有較高的豐度[20], 但垂向分布上在真光層相對較高, 水平分布上在亞南極和極地鋒附近海域的豐度相對較高[23]。a-變形菌綱中另一個常見的類群為玫瑰桿菌屬, 但其所占的比例要低于全球其他海域[10,24]。a-變形菌綱中的SAR116類群在全球海洋中均有分布, 在亞南極和南極浮游細菌豐度較低的海域也檢測到了該類群[10]。
南大洋中, γ-變形菌綱豐度很高, 其中SAR86類群是南極表層海水中的主要類群[25]。該類群可以代謝脂類和碳水化合物, 并且與SAR11類群具有不同的碳水化合物代謝途徑, 因而兩者的競爭性很小[9]。CFB在南大洋分布較廣, 其占比的高低與浮游植物水華出現與否密切相關[24], 其中擬桿菌中的黃桿菌屬細菌也是顆粒物附著菌的主要類群[26-27]。
β-變形菌綱占比較低但分布廣泛, δ-變形菌綱主要存在于深層海水中[28-29], 而藍細菌類群主要出現在亞南極水域[25,30]。其他出現的類群包括疣微菌門、浮霉菌門和放線菌門等。
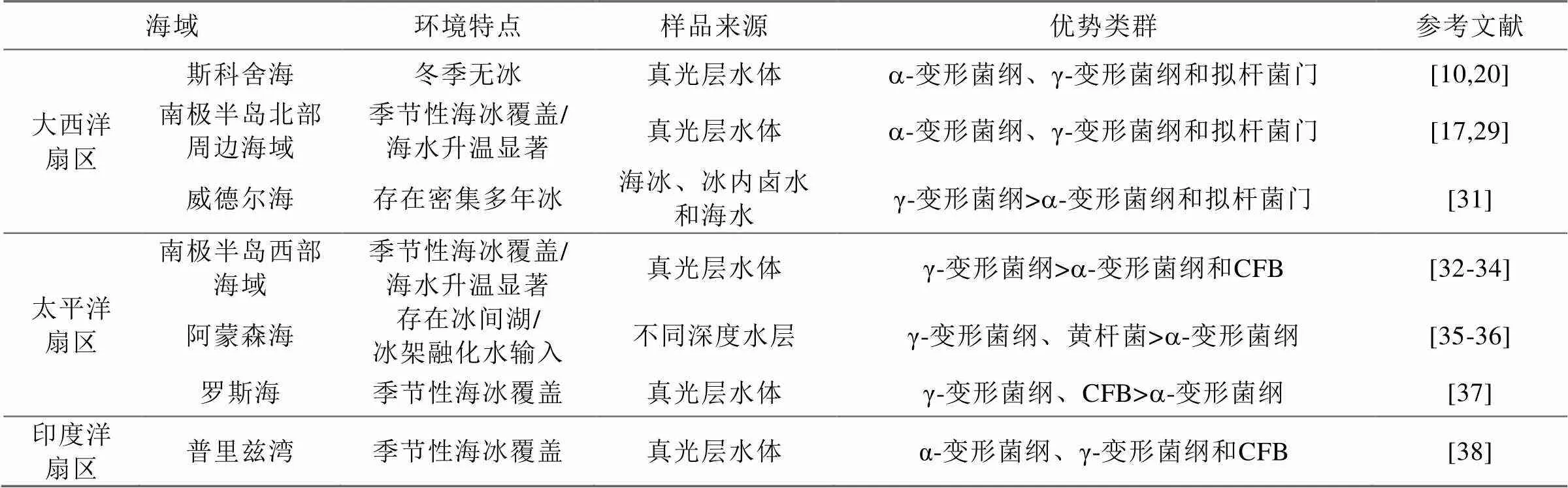
表1 南大洋不同海域浮游細菌優勢類群組成
2 南大洋浮游細菌空間分布
2.1 大西洋扇區
斯科舍弧鄰近海域表層海水浮游細菌優勢類群為α-變形菌綱、γ-變形菌綱和噬纖維菌-屈撓桿菌-擬桿菌群(Cytophaga-Flexibacter-Bacteroides, CFB), 不同站位間優勢類群組成較為一致, 但稀有類群組成存在明顯差異[20]。斯科舍海a-變形菌綱占比最高, 其次為γ-變形菌綱和鞘脂桿菌綱/黃桿菌綱[10]。其中,a-變形菌綱主要包括SAR11類群、玫瑰桿菌屬和SAR116類群, γ-變形菌綱主要為SAR86類群。
南設得蘭群島喬治王島珀特灣浮游細菌的優勢類群為變形菌門, 其次是桿菌門和放線菌門[15]。對該地區智利灣的研究則表明, 其優勢類群為變形菌門和桿菌門, 浮游植物水華期間較為常見的類群有α-變形菌綱中的玫瑰桿菌屬、遠洋桿菌屬和屬以及γ-變形菌門中的交替單胞菌屬、海桿菌屬、交替假單胞菌屬和海洋螺菌科[39]。
南極半島北端鄰近海域浮游細菌群落包括變形菌門、擬桿菌門、藍細菌門、放線菌門、浮霉菌門和疣微菌門細菌。其中a-變形菌綱, γ-變形菌綱和擬桿菌門為優勢類群[18]。對威德爾海冰緣區的研究表明, 浮冰內部和冰下海水中的細菌群落存在明顯差異。冰下海水中的可培養細菌以非發酵的革蘭氏陰性桿菌為主, 而部分海水細菌群落中, γ-變形菌綱可占到 25%以上[31]。
2.2 太平洋扇區
從澳大利亞霍巴特到東南極默茨冰川斷面的調查表明, 極地鋒以北和以南浮游細菌群落組成存在明顯差異。極地鋒以北海域優勢類群為SAR11/SAR116和藍細菌門, 而以南則為γ-變形菌綱、桿菌門、疣微菌門和紅細菌目[28]。
布蘭斯菲爾德海峽浮游細菌優勢類群為γ-變形菌綱和擬桿菌門, 其他包括α-變形菌綱、δ-變形菌綱和SAR406類群等。其中, γ-變形菌綱主要包括海洋螺菌目、弧菌目和交替單胞菌目, 擬桿菌門主要包括黃桿菌目, 而α-變形菌綱主要包括紅細菌目[32]。
南極半島美國帕爾默站近岸海域浮游細菌主要類群包括: γ-變形菌綱、α-變形菌綱、δ-變形菌綱和擬桿菌門, 其中γ-變形菌綱在冬季占有優勢[33]; 該海域另有研究表明, 浮游細菌以γ-變形菌綱為優勢類群, 占比為47%~53%, α-變形菌綱占22%~27%, 而擬桿菌占比為8%~17%, 其他類群占比不足6%[28]。而對南極半島英國羅瑟拉站附近瑞德灣的研究顯示, 其浮游細菌類群包括γ-變形菌綱、β-變形菌綱、α-變形菌綱、放線菌門和CFB簇群[34]。
阿蒙森海是南大洋太平洋扇區重要的邊緣海之一, 也是近年來我國新開辟的調查海域[40]。研究表明, 冰間湖表層海水優勢類群為黃桿菌門的極桿菌屬和γ-變形菌綱海洋螺菌科的巴氏發菌屬, 深層的優勢類群為a-變形菌綱的遠洋桿菌屬[35-36]。
羅斯海近岸海域浮游細菌優勢類群為γ-變形菌綱和 CFB, 與海冰細菌優勢類群類似[37]。
2.3 印度洋扇區
普里茲灣浮游細菌優勢類群主要包括CFB類群、γ-變形菌綱和α-變形菌綱細菌, 其他類群包括δ-變形菌綱、疣微菌目、浮霉菌門、芽單胞菌門和一個分類地位未定類群ANT4E12[38]。另有研究顯示, 夏季普里茲灣達恩利角附近海域可培養細菌分屬α-變形菌綱、γ-變形菌綱和擬桿菌門3個類群下的35個不同分類單元, 其中α-變形菌綱為優勢類群[41]。亞南極凱爾蓋朗群島近岸海域中, α-變形菌綱為最優勢類群(33%~38%), 其次是β-變形菌綱和γ-變形菌綱[29]。
3 南大洋浮游細菌季節性演替
受極晝/極夜交替和海冰生消的影響, 南大洋浮游細菌群落結構存在明顯的季節變化特征[33,38]。研究顯示, 南極半島近岸海域浮游細菌豐富度在冬季要明顯高于夏季, 細菌屬的數量在冬季要高出夏季時的3倍以上, 其中隸屬γ-變形菌綱的屬的數量在冬季為21個、夏季僅有7個[33]。
對南極半島和凱爾蓋朗群島近岸海域浮游細菌一整年的研究表明, 全年最優勢類群為γ-變形菌綱, 其中夏季優勢亞群為Ant4D3、冬季則為Ant10A4, 擬桿菌門黃桿菌綱豐富度在夏季明顯高于冬季, 而δ-變形菌和脫鐵桿菌門僅在冬季被檢測到[29]。
普里茲灣浮游細菌優勢類群呈現明顯的季節變化, 其中春初浮游細菌群落主要由CFB類群、γ-變形菌綱和α-變形菌綱細菌組成, 但春末夏初隨著春季浮游植物水華形成, CFB類群開始占據主導地位[38]。
總體而言, 冬季浮游細菌表現出更高的系統發育多樣性和功能潛力, 同時夏季細菌的代謝主要為化學異養、光合異養和好氧不產氧光合作用, 而冬季則具備化能無機自養能力[20,33]。
4 南大洋不同水團的浮游細菌
就空間分布而言, 上表層和中深層生態系統可能具有兩種不同的微生物群落發育和維持機制。上表層浮游細菌以變形菌門、擬桿菌門和藍細菌門為優勢類群, 由于易受短期環境變化的影響, 不同站位浮游細菌群落結構存在顯著差異。隨著深度的增加, 藍細菌門和a-變形菌綱相對豐度減少, γ-變形菌綱相對豐度增加。中深層受較長周期的水團駐留和較為穩定的環境影響, 群落結構較為一致, γ-變形菌等為優勢類群[12]。相比上表層浮游細菌群落, 中深層水團中的黃桿菌比例較低, 且檢測出未分類細菌序列的比例較高。
對阿蒙森海冰間湖浮游細菌的研究表明, 表層生物多樣性相對較低, 以富營養型類群占據優勢, 而中深層水團中多樣性較高, 以寡營養型類群占據優勢[35]。從表層到深層, 南極表層水、冬季殘留水、繞極深層水中的浮游細菌群落結構存在明顯差異: 表層水中的多樣性低, 優勢類群為黃桿菌科極桿菌屬以及海洋螺菌科海洋螺菌屬; 冬季殘留水和繞極深層水中浮游細菌群落的主要差別是優勢類群占比不同, 并且繞極深層水中含有其他兩個水團少有的豐富類群[35]。
水文過程同樣會影響浮游細菌的群落結構。對南極半島英國羅瑟拉站附近瑞德灣浮游細菌的研究表明, 在春夏季上層躍層被逐步混合的過程中, 浮游植物優勢藻種從輻環藻屬種轉變為海鏈藻屬種, 數天后, 浮游細菌優勢類群從α-變形菌綱和γ-變形菌綱更替為CFB簇群[34]。南大洋海冰凍結可以產生高鹽冷水團并下沉到海底, 形成溫度、鹽度和溶解氧等環境要素的垂直梯度, 導致浮游細菌群落呈明顯的垂直分布模式[12]。另有研究表明, 對浮游細菌群落結構的影響, 除環境和距離效應外, 對流可以長距離輸運浮游細菌并使其可能在新的環境下生長形成優勢[28]。
5 南大洋浮游細菌群落結構調控因子
5.1 環境因子
5.1.1 光照
光照可直接影響部分細菌類群, 除藍細菌的典型產氧光合作用外, 部分類群的光合營養可通過好氧不產氧光合作用(如紅細菌科)或依托細菌視紫紅質富集能量(如極桿菌屬、SAR11以及部分γ-變形菌綱種類)[42-43], 加上部分異養種群生長存在光抑制現象, 夏季光照顯著增強會改變海洋表層浮游細菌的群落結構。南大洋相關研究有限。對阿蒙森海浮游細菌群落的模擬實驗表明, 光照和對有機營養物質的競爭力是調控浮游細菌群落結構的關鍵因子[35]。但實驗研究同時表明, 春季較強的紫外輻射對群落結構的影響并不顯著[38]。
5.1.2 溫度和鹽度
溫度和鹽度是影響微生物群落結構的重要因子[17]。海水溫度直接影響細菌的新陳代謝能力, 同時通過影響浮游植物初級生產和光合作用過程中溶解有機質(Dissolved Organic Matter, DOM)的釋放間接影響細菌生產力。對斯科舍海的調查顯示, 溫度是影響浮游細菌群落結構的第一影響因子[12]。模式研究則顯示, 全球變化導致表層海水升溫, 在21世紀后半葉對浮游細菌和古菌的影響是顯著而持續的[16]。
(9)Property Rules,Liability Rules,and Inalienability:One Viewof the Cathedral.
夏季近岸海域受到冰雪融化水注入的影響, 表層海水鹽度明顯下降。對西南極南設得蘭群島喬治王島珀特灣為期1年的研究表明, 鹽度、溫度和顆粒物是影響群落結構的主要環境因子[15]。該灣在夏季時的浮游細菌群落結構與河口附近的沿海地區類似, 而秋冬季的群落結構則更接近于大洋水域, 表明浮游細菌群落的結構受到鹽度的強烈制約[15]。
對南極的研究表明, 寒冷環境可能會提高浮游細菌的生物多樣性。對普里茲灣近岸海域的研究表明, 冰區浮游細菌具有相對較高的生物多樣性[37]。而對亞南極凱爾蓋朗群島和南極半島近海海域的研究同樣顯示, 冬季的浮游細菌多樣性更高, 其OTUs數量比全年其他季節的要多出約60%[29]。
5.1.3 海冰與淡水輸入
海冰內部存在著一個有別于海水的特殊微生物區系[31,44]。海冰融化會導致生長在其中的微生物釋放至表層海水, 從而影響表層海水浮游細菌群落結構。對羅斯海西海岸近岸海域表層海水浮游細菌的研究顯示, γ-變形菌綱和擬桿菌門占優, 其中變形菌綱中的優勢類群為桿嗜冷菌和嗜冷桿菌屬, 擬桿菌門的優勢類群為極桿菌屬和彎桿菌屬, 這些類群與海冰密切相關[37]。
對冰山追蹤和實驗研究顯示, 冰山漂移導致的水體混合和營養狀況改變, 會影響海洋浮游細菌群落的活性、組成以及物質利用能力[14]。氣溫上升導致冰山數量的增加, 可以影響局域碳和營養物質的生物地球化學循環。而夏季陸上冰雪融化導致淡水輸入近岸海域, 會把陸上細菌帶入海洋, 同樣會影響表層浮游細菌群落結構。羅斯海西海岸羅德灣表層海水中發現的β-變形菌和ε-變形菌與沉積和淡水相關, 推測與附近意大利站的人類活動有關[37]。
5.1.4 其他
對南大洋25個主要水團的研究顯示, 即使在環境要素和空間分割受控的情況下, 水團對流距離與微生物種類組成差異性存在正相關, 表明“對流效應”是塑造微生物群落結構的一個因素[45]。
對羅斯海表層海水浮游細菌的酸化實驗結果顯示, 海水pH下降會提高細胞的酶活性, 加速碳水化合物和脂肪的降解以及磷酸鹽的再循環, 導致細菌豐度增加和多樣性下降[46]。而對阿蒙森海冰間湖區的研究表明, 海水中的無機營養鹽主要影響浮游細菌的豐度而非多樣性[47]。
5.2 生物因子
5.2.1 浮游植物
浮游植物光合作用過程中釋放的DOM是浮游細菌的主要營養源, 因而浮游植物的生物量和組成會顯著影響浮游細菌的群落結構[48-49]。對南極半島近岸海域的研究表明, 細菌群落結構會隨著浮游植物組成的變化而變化, 但存在時間上的滯后[34]。與此同時, 浮游細菌與浮游植物間存在對海洋中限制性鐵(Fe)元素的競爭。實驗和模式研究均顯示, 增加Fe濃度和提高光照強度有利于浮游植物主導Fe的利用, 而提高溶解有機碳(Dissolved Organic Carbon, DOC)濃度和降低Fe濃度可促進浮游細菌對Fe的吸收[50]。在高DOC、低Fe濃度的環境下, 浮游細菌對Fe具有極強的競爭力, 浮游植物釋放的DOC可調節相互競爭的強度。
對南極南設得蘭群島智利灣的研究發現, 浮游細菌的多樣性與浮游植物豐度密切相關, 浮游植物水華期間浮游細菌群落的多樣性明顯較高, 主要類群為α-變形菌綱和擬桿菌門[39]。對普里茲灣的研究表明, 細菌多樣性與浮游植物量和生產力水平呈現正相關[41]。但對阿蒙森海的研究則顯示, 浮游植物水華通過顆粒物沉降等過程會提高真光層以下水層中浮游細菌的多樣性[47]。而對南極半島和亞南極凱爾蓋朗群島近海海域研究同樣表明, 浮游細菌群落結構和生物多樣性的變化與浮游植物水華密切相關, DOC是冬季期間支配浮游細菌群落的主要因子[29,51]。
預測表明南大洋浮游植物群落會受到氣候變化的顯著影響[52], 這將對包括浮游細菌在內的整個南大洋生態系統產生深遠影響。
5.2.2 原生動物
海洋原生動物是浮游細菌最主要的捕食者, 其捕食行為可以直接抑制細菌生物量的增長, 同時捕食過程也會導致DOM 的釋放和無機鹽的再生, 從而促進浮游細菌的生長。對南大洋的研究顯示, 原生動物可以消耗90%的日細菌產量[53], 但對于浮游細菌生物群落的影響, 目前尚缺乏相關研究。
5.2.3 內部競爭
浮游細菌群落結構和多樣性不僅受環境因素的影響, 也會受到不同細菌類群間的相互合作或競爭的影響[54-55]。例如, 亞硫酸鹽桿菌是一種亞硫酸鹽氧化細菌, 而海桿菌、交替單胞菌和食堿菌亞硫酸鹽桿菌能夠利用碳氫化合物, 它們之間可以很好地合作共生[56]。具有相同或者相似生態功能的浮游細菌優勢類群, 彼此間會為爭取相同或者相似的生境產生競爭。如α-變形菌中的SAR11類群存在于特定生境, 呈寡營養方式生活[57], 玫瑰桿菌類群則具有諸如厭氧光合[58]、一氧化碳氧化[59]以及對一系列有機物的異養利用[30]等多種代謝途徑, 而SAR116類群兼具SAR11類群和玫瑰桿菌類群的功能, 它們之間存在競爭關系。在寡營養海域, 具有多種代謝功能有利于細菌在競爭關系中占據優勢。另外, 具有自養能力的藍細菌在表層海水中廣泛分布, 但在黃桿菌豐度較高的表層海域卻很少能被檢測到[37]。對南極半島北端海域表層浮游細菌群落的研究表明, 在夏季其優勢屬間的相互作用是浮游細菌群落結構的主控因子[11]。
6 展望
浮游細菌在極區海洋生態系統中扮演著十分重要的角色。高通量測序技術的逐漸成熟、生物信息學的發展以及相關數據的積累, 在一定程度上增進了對于極地浮游細菌群落結構的認知。但目前對南極浮游細菌群落結構的研究, 主要集中在對優勢類群的分析, 對小眾類群的研究極為有限, 導致未能鑒定OTUs的比例偏高。與此同時, 盡管浮游細菌群落在南大洋發揮著至關重要的作用, 但對它們的分布和季節變化的研究仍然有限, 研究區域主要集中在西南極、特別是南極半島近岸海域, 而對開闊大洋的調查偏少、對東南極廣闊海域的研究很少。氣候變化導致局域海水升溫和海冰減少, 會對生態系統造成深遠影響[60-61], 但對浮游細菌群落的相關研究和預測仍極為有限[16]。為此, 今后應重點開展以下幾個方面的研究。
1.群落組成與空間分布研究。加強船基大洋調查與研究, 補充東南極海域和冰區浮游細菌群落研究, 開展小眾類群結構及調控機理研究, 提升對南大洋浮游細菌群落的系統了解。
2.群落季節變化研究。加強依托考察站的近岸浮游細菌群落研究, 重點分析海冰變化對群落結構的影響。加強對浮冰區冰下浮游細菌群落季節變化研究, 重點開展融冰期冰緣區浮游細菌群落及其與冰緣浮游植物水華的相關性分析。
3.深層群落結構及調控機理研究。開展深海浮游細菌群落結構研究, 探索群落空間分布特征及其調控機理, 與其他大洋深層群落的特異性對比, 分析水文過程和上層海域顆粒物沉降對深層海域群落結構的潛在影響。
4.氣候變化潛在影響研究。研究氣候變化導致的海水增暖、水團變異、海洋酸化等環境快速變化對浮游細菌群落的影響, 發展南大洋海洋微食物環模型, 預測在全球變化背景下浮游細菌群落的調控機理及潛在變化。
1 BLACKBURN N, FENCHEL T, MITCHELL J. Microscale nutrient patches in planktonic habitats shown by chemotactic bacteria[J]. Science, 1998, 282(5397): 2254-2256.
2 LE B P J WILLIAMS. The balance of plankton respiration and photosynthesis in the open oceans[J]. Nature, 1998, 394(6688): 55-57.
3 AZAM F. Microbial control of oceanic carbon flux: The plot thickens[J]. Science, 1998, 280(5364): 694-696.
4 KARL D M. Nutrient dynamics in the deep blue sea[J]. Trends in Microbiology, 2002, 10(9): 410-418.
5 LOREAU M, NAEEM S, INCHAUSTI P, et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges[J]. Science, 2001, 294(5543): 804-808.
6 GIOVANNONI S J, STINGL U. Molecular diversity and ecology of microbial plankton[J]. Nature, 2005, 437(7057): 343-348.
7 MURRAY A E, GRZYMSKI J J. Diversity and genomics of Antarctic marine micro-organisms[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2007, 362(1488): 2259-2271.
8 DELILLE D. Abundance and function of bacteria in the Southern Ocean[J]. Cellular and Molecular Biology, 2004, 50(5): 543-551.
9 DUPONT C L, RUSCH D B, YOOSEPH S, et al. Genomic insights to SAR86, an abundant and uncultivated marine bacterial lineage[J]. The ISME Journal, 2012, 6(6): 1186-1199.
10 TOPPING J N, HEYWOOD J L, WARD P, et al. Bacterioplankton composition in the Scotia Sea, Antarctica, during the austral summer of 2003[J]. Aquatic Microbial Ecology, 2006, 45(3): 229-235.
11 CAO S, ZHANG W, DING W, et al. Structure and function of the Arctic and Antarctic marine microbiota as revealed by metagenomics[J]. Microbiome, 2020, 8(1): 47.
12 YU Z, YANG J, LIU L, et al. Bacterioplankton community shifts associated with epipelagic and mesopelagic waters in the Southern Ocean[J]. Scientific Reports, 2015, 5(1):12897.
13 GHIGLIONE J F, GALAND P E, POMMIER T, et al. Pole-to-pole biogeography of surface and deep marine bacterial communities[J]. PNAS, 2012, 109(43): 17633-17638.
14 DINASQUET J, RICHERT I, LOGARES R, et al. Mixing of water masses caused by a drifting iceberg affects bacterial activity, community composition and substrate utilization capability in the Southern Ocean[J]. Environmental Microbiology, 2017, 19(6): 2453-2467.
15 HERNáNDEZ E A, LOPEZ J L, PIQUET A M T, et al. Changes in salinity and temperature drive marine bacterial communities’ structure at Potter Cove, Antarctica[J]. Polar Biology, 2019, 42(12): 2177-2191.
16 TONELLI M, SIGNORI C N, BENDIA A, et al. Climate projections for the Southern Ocean reveal impacts in the marine microbial communities following increases in sea surface temperature[J]. Frontiers in Marine Science, 2021, 8: 636226.
17 LIU Q, JIANG Y. Application of microbial network analysis to discriminate environmental heterogeneity in Fildes Peninsula, Antarctica[J]. Marine Pollution Bulletin, 2020, 156: 111244.
18 CAO S, HE J, ZHANG F, et al. Diversity and community structure of bacterioplankton in surface waters off the northern tip of the Antarctic Peninsula[J]. Polar Research, 2019, 38: 3491.
19 WANG Z B, SUN Y Y, LI Y, et al. Significant bacterial distance-decay relationship in continuous, well-connected Southern Ocean surface water[J]. Microbial Ecology, 2020, 80(1): 73-80.
20 JAMIESON R E, ROGERS A D, BILLETT D S M, et al. Patterns of marine bacterioplankton biodiversity in the surface waters of the Scotia Arc, Southern Ocean[J]. FEMS Microbiology Ecology, 2012, 80(2): 452-468.
21 MILICI M, VITAL M, TOMASCH J, et al. Diversity and community composition of particle-associated and free-living bacteria in mesopelagic and bathypelagic Southern Ocean water masses: Evidence of dispersal limitation in the Bransfield Strait[J]. Limnology and Oceanography, 2017, 62(3): 1080-1095.
22 MORRIS R M, RAPPé M S, CONNON S A, et al. SAR11 clade dominates ocean surface bacterioplankton communities[J]. Nature, 2002, 420(6917): 806-810.
23 GIEBEL H A, BRINKHOFF T, ZWISLER W, et al. Distribution ofRCA and SAR11 lineages and distinct bacterial communities from the subtropics to the Southern Ocean[J]. Environmental Microbiology, 2009, 11(8): 2164-2178.
24 BUCHAN A, GONZáLEZ J M, MORAN M A. Overview of the marinelineage[J]. Applied and Environmental Microbiology, 2005, 71(10): 5665-5677.
25 ABELL G C J, BOWMAN J P. Ecological and biogeographic relationships of class Flavobacteria in the Southern Ocean[J]. FEMS Microbiology Ecology, 2005, 51(2): 265-277.
26 DELONG E F, FRANKS D G, ALLDREDGE A L. Phylogenetic diversity of aggregate-attached vs. free-living marine bacterial assemblages[J]. Limnology and Oceanography, 1993, 38(5): 924-934.
27 PINHASSI J, SALA M M, HAVSKUM H, et al. Changes in bacterioplankton composition under different phytoplankton regimens[J]. Applied and Environmental Microbiology, 2004, 70(11): 6753-6766.
28 WILKINS D, LAURO F M, WILLIAMS T J, et al. Biogeographic partitioning of Southern Ocean microorganisms revealed by metagenomics[J]. Environmental Microbiology, 2013, 15(5): 1318-1333.
29 GHIGLIONE J F, MURRAY A E. Pronounced summer to winter differences and higher wintertime richness in coastal Antarctic marine bacterioplankton[J]. Environmental Microbiology, 2012, 14(3): 617-629.
30 BRINKHOFF T, GIEBEL H A, SIMON M. Diversity, ecology, and genomics of theclade: A short overview[J]. Archives of Microbiology, 2008, 189(6): 531-539.
31 DELILLE D. Marine bacterioplankton at the Weddell Sea ice edge, distribution of psychrophilic and psychrotrophic populations[J]. Polar Biology, 1992, 12(2): 205-210.
32 SIGNORI C N, THOMAS F, ENRICH-PRAST A, et al. Microbial diversity and community structure across environmental gradients in Bransfield Strait, Western Antarctic Peninsula[J]. Frontiers in Microbiology, 2014, 5: 647.
33 WILLIAMS T J, LONG E, EVANS F, et al. A metaproteomic assessment of winter and summer bacterioplankton from Antarctic Peninsula coastal surface waters[J]. The ISME Journal, 2012, 6(10): 1883-1900.
34 PIQUET A M T, BOLHUIS H, MEREDITH M P, et al. Shifts in coastal Antarctic marine microbial communities during and after melt water-related surface stratification[J]. FEMS Microbiology Ecology, 2011, 76(3): 413-427.
35 RICHERT I, DINASQUET J, LOGARES R, et al. The influence of light and water mass on bacterial population dynamics in the Amundsen Sea Polynya[J]. Elementa: Science of the Anthropocene, 2015, 3(2): 000044.
36 KIM J G, PARK S J, QUAN Z X, et al. Unveiling abundance and distribution of planktonic Bacteria and Archaea in a polynya in Amundsen Sea, Antarctica[J]. Environmental Microbiology, 2014, 16(6): 1566-1578.
37 GENTILE G, GIULIANO L, D’AURIA G, et al. Study of bacterial communities in Antarctic coastal waters by a combination of 16S rRNA and 16S rDNA sequencing[J]. Environmental Microbiology, 2006, 8(12): 2150-2161.
38 PIQUET A M T, BOLHUIS H, DAVIDSON A T, et al. Seasonal succession and UV sensitivity of marine bacterioplankton at an Antarctic coastal site[J]. FEMS Microbiology Ecology, 2010, 73(1): 68-82.
39 FUENTES S, ARROYO J I, RODRíGUEZ-MARCONI S, et al. Summer phyto- and bacterioplankton communities during low and high productivity scenarios in the Western Antarctic Peninsula[J]. Polar Biology, 2019, 42(1): 159-169.
40 李丙瑞. 中國第35次南極科學考察簡報[J]. 極地研究, 2019, 31(3): 364-367.
41 吳月紅, 韓正兵, 張心齊, 等. 南極普里茲灣可培養浮游細菌群落結構研究[J]. 極地研究, 2014, 26(2): 222-229.
42 GIOVANNONI S J, BIBBS L, CHO J C, et al. Proteorhodopsin in the ubiquitous marine bacterium SAR11[J]. Nature, 2005, 438(7064): 82-85.
43 COTTRELL M T, KIRCHMAN D L. Photoheterotrophic microbes in the Arctic Ocean in summer and winter[J]. Applied and Environmental Microbiology, 2009, 75(15): 4958-4966.
44 BECQUEVORT S, DUMONT I, TISON J L, et al. Biogeochemistry and microbial community composition in sea ice and underlying seawater off East Antarctica during early spring[J]. Polar Biology, 2009, 32(6): 879-895.
45 WILKINS D, VAN SEBILLE E, RINTOUL S R, et al. Advection shapes Southern Ocean microbial assemblages independent of distance and environment effects[J]. Nature Communications, 2013, 4(1): 2457.
46 MAAS E W, LAW C S, HALL J A, et al. Effect of ocean acidification on bacterial abundance, activity and diversity in the Ross Sea, Antarctica[J]. Aquatic Microbial Ecology, 2013, 70(1): 1-15.
47 RICHERT I, YAGER P L, DINASQUET J, et al. Summer comes to the Southern Ocean: How phytoplankton shape bacterioplankton communities far into the deep dark sea[J]. Ecosphere, 2019, 10(3): e02641.
48 DELMONT T O, HAMMAR K M, DUCKLOW H W, et al.blooms strongly influence bacterial community structures in the Amundsen Sea polynya[J]. Frontiers in Microbiology, 2014, 5(5): 646.
49 LANDA M, BLAIN S, CHRISTAKI U, et al. Shifts in bacterial community composition associated with increased carbon cycling in a mosaic of phytoplankton blooms[J]. The ISME Journal, 2016, 10(1): 39-50.
50 RATNARAJAH L, BLAIN S, BOYD P W, et al. Resource colimitation drives competition between phytoplankton and bacteria in the Southern Ocean[J]. Geophysical Research Letters, 2021, 48(1): e2020GL088369.
51 LANDA M, BLAIN S, HARMAND J, et al. Major changes in the composition of a Southern Ocean bacterial community in response to diatom-derived dissolved organic matter[J]. FEMS Microbiology Ecology, 2018, 94(4): fiy034.
52 DEPPELER S L, DAVIDSON A T. Southern Ocean phytoplankton in a changing climate[J]. Frontiers in Marine Science, 2017, 4(40): 1-28.
53 BECQUEVORT S, MENON P, LANCELOT C. Differences of the protozoan biomass and grazing during spring and summer in the Indian sector of the Southern Ocean[J]. Polar Biology, 2000, 23(5): 309-320.
54 DANG H Y, LOVELL C R. Microbial surface colonization and biofilm development in marine environments[J]. Microbiology and Molecular Biology Reviews, 2015, 80(1): 91-138.
55 KOSTKA J E, PRAKASH O, OVERHOLT W A, et al. Hydrocarbon-degrading bacteria and the bacterial community response in gulf of Mexico beach sands impacted by the deepwater horizon oil spill[J]. Applied and Environmental Microbiology, 2011, 77(22): 7962-7974.
56 MAS-LLADó M, PI?A-VILLALONGA J M, BRUNET-GALMéS I, et al. Draft genome sequences of two isolates of thegroup,sp. strains 3SOLIMAR09 and 1FIGIMAR09, from harbors of Mallorca Island (Mediterranean Sea)[J]. Genome Announcements, 2014, 2(3): e00350-14.
57 BROWN M V, LAURO F M, DEMAERE M Z, et al. Global biogeography of SAR11 marine bacteria[J]. Molecular Systems Biology, 2012, 8(1): 595.
58 BIEBL H, ALLGAIER M, TINDALL B J, et al.gen. nov. sp. nov., a new aerobic phototrophic bacterium isolated from dinoflagellates[J]. International Journal of Systematic and Evolutionary Microbiology, 2005, 55: 1089-1096.
59 KING G M. Molecular and culture-based analyses of aerobic carbon monoxide oxidizer diversity[J]. Applied and Environmental Microbiology, 2003, 69(12): 7257-7265.
60 SALLéE J-B. Southern Ocean warming[J]. Oceanography, 2018, 31(2): 52-62.
61 PINKERTON M H, BOYD P W, DEPPELER S, et al. Evidence for the impact of climate change on primary producers in the Southern Ocean[J]. Frontiers in Ecology and Evolution, 2021, 9: 592027.
Progress in the study of bacterioplankton community structures in the Southern Ocean
Sun Yurong1,2, Ma Yuxin3,4,1, Cao Shunan2,3, Luo Guangfu2,3, Lan Musheng2,3, He Jianfeng2,3,4,5
(1College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China;2Antarctic Great Wall Polar Ecology National Scientific Observation and Research Station, Polar Research Institute of China, Shanghai 200136, China;3MNR Key Laboratory for Polar Science, Polar Research Institute of China, Shanghai 200136, China;4School of Oceanography, Shanghai Jiao Tong University, Shanghai 200030, China;5College of Environmental Science and Engineering, Tongji University, Shanghai 200092, China)
Bacterioplankton is an important part of the marine microbial loop, playing a role in maintaining the stability of the marine ecosystem and material circulation. This paper summarizes the community composition of bacterioplankton and its influencing factors in different parts of the Southern Ocean. The results show that the dominant groups are α-proteobacteria, γ-proteobacteria, and Cytophaga-Flexibacter-Bacteroides (CFB), while less abundant groups exhibit strong spatial variability. Temperature and dissolved organic carbon concentrations are two of the main environmental factors that regulate the community structure. There is an obvious interaction between bacterioplankton and other compnents of the microbial food web. Environmental changes such as increasing water temperature and ocean acidification caused by climate change may affect the bacterioplankton community. In the future, research on the bacterioplankton in the pelagic waters, pack ice zone, and deep waters, should be strengthened. In addition, research on seasonal community changes in nearshor waters should be conducted, as should attempts to predict the potential changes and regulation mechanisms of the bacterioplanktonic community in the Southern Ocean via both field investigations and modeling.
Southern Ocean, bacterioplankton, community structure, seasonality, environmental factors
2020年12月收到來稿, 2022年3月收到修改稿
極地調查專項(IRASCC-01-02)和國家自然科學基金(41976230)資助
孫玉蓉, 女, 1997年生。碩士, 主要從事海洋微生物學和海洋化學研究。E-mail:1018445900@qq.com
何劍鋒, E-mail: hejianfeng@pric.org.cn
10.13679/j.jdyj.20200079
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中華詩詞(2020年1期)2020-09-21 09:24:52
趣味(語文)(2020年3期)2020-07-27 01:42:46
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學生作文(中高年級適用)(2018年5期)2018-06-11 01:22:56
中學生數理化·七年級數學人教版(2017年11期)2017-04-23 07:18:00
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
數學大王·中高年級(2016年12期)2016-12-26 21:37:36
