茶樹異胡豆苷合酶基因CsSTR1克隆及表達分析
2023-01-07 01:40:18周棋贏李婭菲郭文利張馨月方志貞王儀佳
西南農業學報 2022年10期
周棋贏,李婭菲,郭文利,張馨月,葛 鑫,方志貞,孫 潔,王儀佳,陳 尚
(1.信陽師范學院生命科學學院/茶學與食品科技學院,河南 信陽 464000;2. 河南省茶樹生物學重點實驗室,河南 信陽 464000;3.河南省茶葉精深加工工程技術研究中心,河南 信陽 464000;4.大別山農業生物資源保護與利用研究院,河南 信陽 464000; 5. 信陽師范學院國際教育學院,河南 信陽 464000)
【研究意義】異胡豆苷合酶(Strictosidine synthase, STR)是催化色胺和裂環馬錢子苷通過Pictet-Spengler反應合成異胡豆苷的一類酶。研究表明,異胡豆苷是植物體內具有重要藥用價值的萜類吲哚生物堿(Terpenoid indole alkaloids,TIAs)合成的通用前體,STR常被用作TIAs代謝工程中的候選基因和作用靶點[1-3]。STR基因的表達受低溫、干旱等環境因子,植物激素與病蟲害侵染等因素影響,在植物抗逆反應中具有重要作用[4]。茶是世界公認的三大健康飲料之一,具有重要的健康功效。圍繞茶樹(Camelliasinensis)STR基因(CsSTR)開展研究,為進一步揭示茶葉的藥用價值及茶樹抗性育種具有重要意義。【前人研究進展】STR的三維結構呈六葉β-螺旋漿狀,每葉由4個反向平行的折疊片組成。β-螺旋漿狀結構中有3個α螺旋,其中外部和內部的α螺旋通過Cys-89和Cys-101間的二硫鍵進行連接,這一特征在物種進化過程中具有高度的保守性,當Cys-89被其他氨基酸替代后,STR就會失去活性[5]。因此,該二硫鍵對于維持酶的整體結構和活性具有重要作用。1988年,STR基因首先在蛇根木(Rauvolfiaserpentina)中得到克隆,被命名為STR1。STR1為單分子蛋白質,由344個氨基酸組成,分子量約為35.3 kD[5]。目前,STR基因已經在長春花(Catharanthusroseus)、日本蛇根草(Ophiorrhizajaponica)、短小蛇根草(O.pumila)、金雞納樹(Cinchonacalisaya)、蘿芙木(R.verticillata)、喜樹(Camptothecaacuminata)和臭味假柴龍樹(Nothapodytesfoetida)等植物中得到克隆[5-7]。STR基因在植物生長發育和抗逆反應中具有重要作用。在長春花中的研究發現,紫外、鹽脅迫、激發子、蟲害和MeJA處理都能誘導STR基因表達,而生長素和低溫處理則抑制STR基因表達;Ca2+、蛋白磷酸化和NO在植株響應這些處理及調節STR基因表達中具有重要作用[8]。PEG模擬干旱脅迫條件下,隨著脅迫時間的延長、脅迫強度增強,STR基因表達量逐漸增加[9]。在日本蛇根草中的研究也發現,植物防御信號SA和MeJA能誘導STR基因的表達[10]。對擬南芥中STR基因的表達模式分析表明,擬南芥中STR基因SSL5、SSL6、SSL7都可以被植物防御信號(SA、MeJA、ET), 傷信號以及病原菌侵染誘導,暗示他們在植物的誘導防御反應中有重要作用[11]。近期研究還發現,在全球氣候變暖的趨勢下,STR同源基因SSL4增強了高溫環境下擬南芥對細菌性青枯病的抗性[12]。水稻中一個STR同源物OsSTRL2,在水稻花藥發育和花粉壁形成中具有重要作用[13]。近年來,茶樹基因組測序陸續在大葉種‘云抗10號’、中小葉種‘舒茶早’‘龍井43’和‘碧云’以及野生種‘DASZ’中完成[15-17]。河南省茶樹生物學重點實驗室茶樹功能基因組學課題組前期利用生物信息學分析方法,對茶樹基因組中的CsSTR基因進行了鑒定和分析[18]。目前為止,CsSTR基因的克隆和表達調控分析還鮮有報道。中國自古就有“神農嘗百草,日遇七十二毒,得荼而解之”的歷史記載。現代科學研究也發現,茶具有重要的健康功效[14]。茶樹作為一種固生、木本經濟作物,在生長中常遭受各種逆境脅迫,嚴重影響茶樹的生長、茶葉產量和品質。【本研究切入點】利用RACE克隆,本研究克隆了‘云抗10號’基因組數據庫中的一個茶樹STR基因(CsSTR1),并對其在非生物脅迫和激素處理條件下的表達量進行分析。【擬解決的關鍵問題】本研究為后續對CsSTR1基因的功能研究奠定基礎,為CsSTR1基因的表達調控及其在茶樹抗逆育種中的應用提供理論依據。
1 材料與方法
1.1 RACE克隆
利用“Strictosidine synthase”關鍵詞進行檢索,在‘云抗10號’基因組數據庫[19]中檢索到5個“Strictosidine synthase”基因(CsSTR),其中一個CsSTR基因(CSA035098.1,CsSTR1)的CDS包含不完整的開放閱讀框(ORF),采用RACE方法對其全長cDNA進行克隆。首先,利用RNAiso Plus (Takara)試劑提取‘舒茶早’茶樹葉片的總RNA,利用cDNA 反轉錄試劑盒( Takara) 合成第一鏈cDNA。設計CsSTR1基因片段的擴增引物(F:ATGGACATAGATGAGCACCA; R: AATCCACAACTTGCCATTT TTTTCCT),以第一鏈cDNA為模板,使用KOD FX DNA聚合酶(TOYOBO)進行PCR擴增。擴增程序為:94 ℃ 5 min; 94 ℃ 30 s, 55 ℃ 30 s, 68 ℃ 30 s, 循環30次; 68 ℃ 7 min。此步驟擴增產物連接到pEASY?Blunt克隆載體 (TransGen Biotech),轉化至大腸桿菌DH5α,挑取克隆,經初步PCR鑒定陽性克隆后,送克隆至安徽通用生物公司進行測序。然后,根據SMARTer RACE 5′/3′試劑盒(Clontech)的說明進行:①以 ‘舒茶早’茶樹葉片總RNA為模板,分別反轉錄獲取5′ RACE和3′ RACE cDNA模板;②根據克隆測序得到CsSTR1基因的片段序列,設計CsSTR1基因的RACE克隆引物(5′ RACE引物:CCATTGGCAAAAGCAAGGCCTCTTAG;3′ RACE引物:ACAGACACGAGCACGAGCTTTCACAG);③分別對CsSTR1基因5′ RACE和3′ RACE 進行PCR擴增,將擴增產物分別連接到pEASY?Blunt克隆載體,轉化大腸桿菌DH5α后,挑取單克隆進行測序。最后,通過序列拼接得到CsSTR1基因的全長cDNA序列。
1.2 生物信息學分析
利用ProtParam、Plant-PLoc、CD-Search、SOPMA和SWISS-MODEL在線工具分別對蛋白理化性質、亞細胞定位、保守結構域、蛋白質二級結構和三級結構進行分析。使用MEGA X軟件構建蛋白同源關系進化樹。
1.3 材料及處理方法
取生長健康長勢一致、來源于同一株信陽群體種茶樹的枝條(長度約15 cm)為材料,浸入100 μmol/L脫落酸(ABA)水溶液中進行處理,分別于處理0、3、6、12、24、48 h后,將第2和第3片葉片迅速置液氮中速凍,然后放-80 ℃冰箱保存備用。以相同的方法,將茶樹枝條分別浸入1 mmol/L水楊酸(SA)水溶液、1 mmol/L茉莉酸甲酯MeJA溶液(0.5%乙醇溶解)中進行處理。ABA和SA處理組以滅菌水處理組為對照,MeJA處理組以0.5%乙醇處理組為對照。低溫處理是將茶樹枝條浸入滅菌水中,置于4 ℃冰箱處理;茶樹枝條浸入10% PEG-6000 溶液中模擬干旱處理;以常溫滅菌水處理組為對照。每個取樣時間點設置3個生物學重復。
取根,莖,芽,第1~3葉,花和果材料,迅速置液氮中速凍,然后放-80 ℃冰箱保存備用。每個組織樣品取3個生物學重復。
1.4 qPCR分析
利用RNAiso Plus (Takara)試劑提取上述各樣品的總RNA,通過瓊脂糖凝膠電泳檢測RNA的完整性后,利用cDNA 反轉錄試劑盒(Takara) 合成第一鏈cDNA。根據克隆得到的CsSTR1基因全長cDNA序列,設計基因特異的qPCR引物(CsSTR1-F: AACAACAGACAAGAGATGAAC;CsSTR1-R:GTGGA GGTGGTTATTGGA)。根據前述方法[20],以茶樹CsGAPDH為內參基因(CsGAPDH-F:TTGGCATCGTTGAGGGTCT;CsGAPDH-R:CAGTGGGAACACGGAA AGC),對CsSTR1基因的表達量進行分析。
2 結果與分析
2.1 CsSTR1基因RACE克隆
根據CSA035098.1的CDS序列,通過PCR擴增,得到500~600 bp的條帶(圖1-A);經測序確認,表明克隆序列與目標序列基本一致,但克隆序列長度為591 bp, 與目標序列相比有17個堿基不同,且在對應CSA035098.1 107 bp后有74 bp的堿基插入,在對應CSA035098.1 136 bp后有個G堿基插入。根據591 bp的測序序列設計CsSTR1基因的5′ 和3′ RACE克隆引物。CsSTR1基因的5′ 和3′ RACE克隆分別得到長度為813和 803 bp的片段(圖1-B);經測序分析,表明RACE克隆成功。將前面得到的591 bp擴增序列與5′ 和3′ RACE克隆序列進行拼接,得到CsSTR1基因的全長cDNA序列。CsSTR1基因全長1382 bp,其中CDS 1131 bp,編碼376個氨基酸;5′ UTR 長92 bp,3′ UTR 長159 bp(圖2)。

1. DNA分子質量標準;2. 基因部分片斷擴增產物;3. 5′ RACE擴增產物;4. 3′ RACE 擴增產物

圖2 茶樹CsSTR1基因的cDNA序列及推導的氨基酸序列
2.2 CsSTR1蛋白特征及結構
CsSTR1蛋白理化分析表明,該蛋白相對分子量為41.5 kD,理論等電點為 6.36,不穩定指數為29.74,脂肪族氨基酸指數為88.94,親水性平均系數為-0.077。亞細胞定位預測分析表明,該CsSTR1蛋白為液泡定位蛋白。保守結構域分析顯示,該蛋白在157~252 位氨基酸處具有異胡豆苷合酶Str_synth保守結構域(圖3)。CsSTR1蛋白二級結構主要由無規卷曲、延伸鏈、α螺旋和β轉角組成,占比分別為42.82%、29.79%、16.76%、10.64%(圖4)。三維結構預測結果顯示,CsSTR1蛋白由β折疊片組成六葉β-螺旋漿狀,中間有3個α螺旋(圖5),與結構模板6n5v.1.A間的同源性達35.25%。
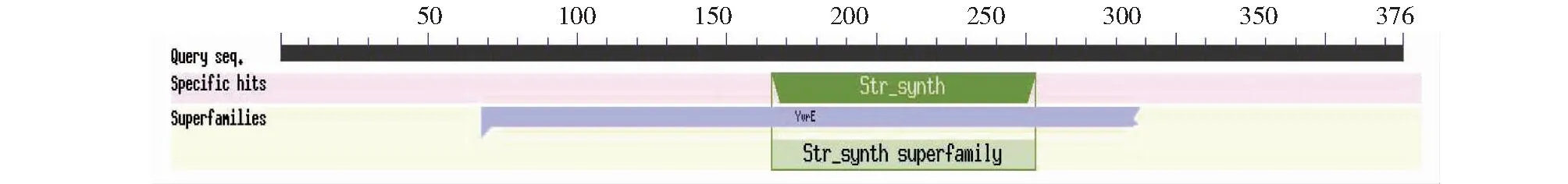
圖3 茶樹CsSTR1蛋白的保守結構域鑒定

圖4 茶樹CsSTR1蛋白的二級結構預測
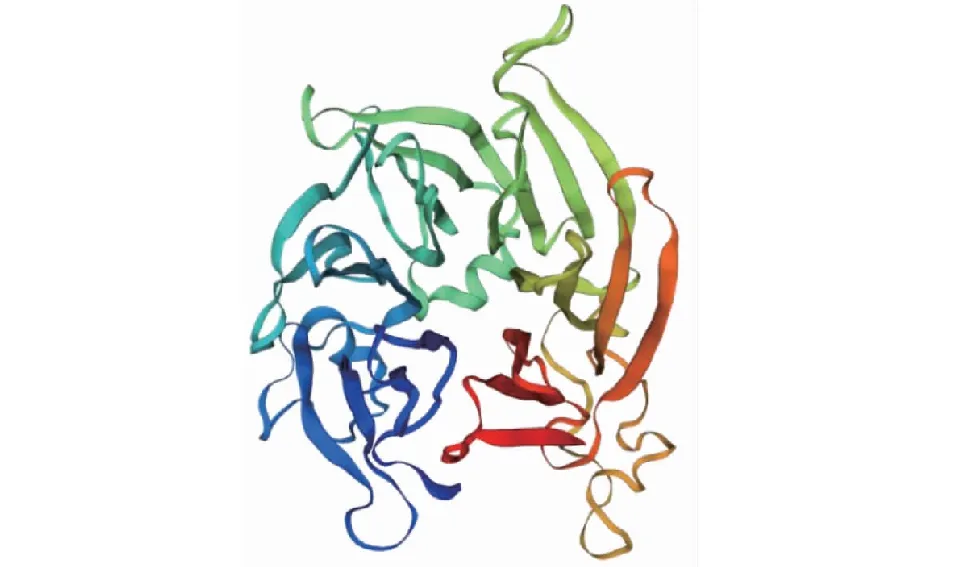
圖5 茶樹CsSTR1蛋白的三級結構預測
2.3 CsSTR1系統進化分析
將CsSTR1與已報到的植物STR蛋白構建系統進化樹,結果顯示CsSTR1與水稻STR蛋白OsSTRL2的親緣關系最近(圖6),而與蘿芙木、蛇根木、長春花等夾竹桃科植物中的STR以及擬南芥中的STR蛋白SSL親緣關系較遠。
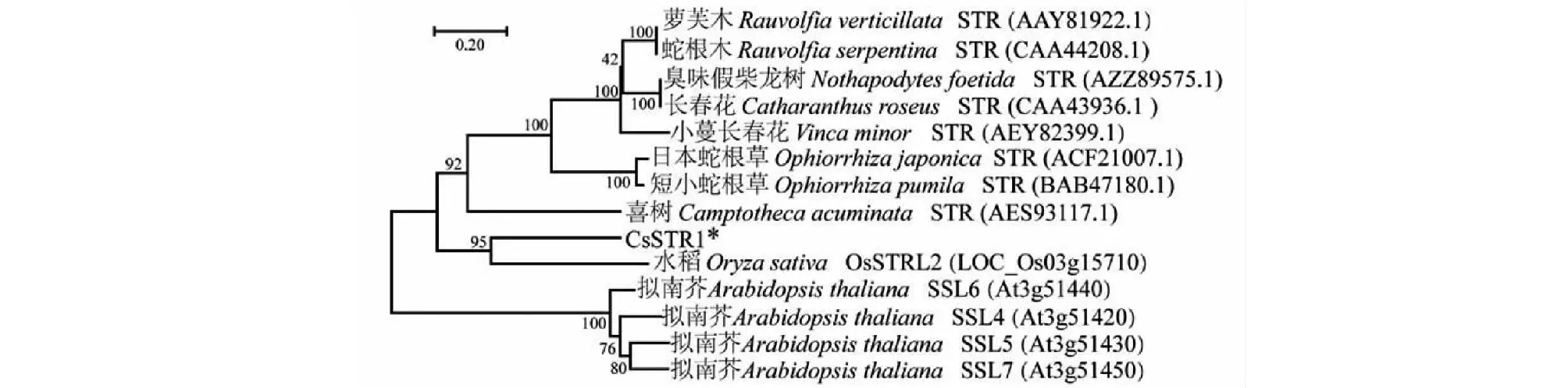
圖6 茶樹CsSTR1蛋白與其他物種STR蛋白的系統進化樹
2.4 CsSTR1基因的表達模式
組織表達模式分析結果表明,CsSTR1基因在芽和嫩葉(第1~3葉)中的相對表達量較高,在老葉、花和果中的相對表達量較低(圖7)。低溫脅迫條件下,CsSTR1基因在3 h后表達量顯著上調,6~24 h 表達量呈下調趨勢,12 h 后CsSTR1基因呈顯著下調表達,48 h后表達量恢復到處理前水平。PEG 模擬干旱處理條件下,CsSTR1基因在24~48 h后呈現顯著上調表達,在48 h后其表達量上調近10倍。MeJA處理后,CsSTR1基因呈現上調表達,在24~48 h后呈現顯著上調。SA處理后,CsSTR1基因呈現顯著上調表達,3 h后其表達水平上調3.3倍。ABA處理后,CsSTR1基因在3~12 h表達變化不大,但在24~48 h后呈顯著下調表達。
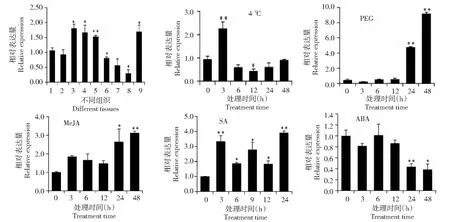
1:根; 2:莖; 3:第1葉;4:第2葉;5:第3葉;6:老葉;7:花;8:果;9:芽.*表示在0.05水平上差異顯著,**表示在0.01水平上差異顯著
3 討 論
STR是植物體內調控TIAs合成的重要酶類,還在植物的生長發育和逆境反應中具有重要作用[1-4, 13]。近年來,茶樹基因組的破譯為茶樹優質基因資源的挖掘利用以及茶葉藥理作用的進一步研究提供了資料。前期研究發現茶樹基因組中具有17個STR基因[18],但目前還沒有一個CsSTR基因得到克隆。河南省茶樹生物學重點實驗室茶樹功能基因組學課題組對CsSTR基因的克隆起始于最早破譯的‘云抗10號’茶樹基因組數據,由于其基因組中一個CsSTR基因(CsSTR1)的CDS包含不完整的開放閱讀框(ORF),利用RACE技術,對CsSTR1基因進行了克隆。結果表明,克隆的CsSTR1基因CDS包含完整的編碼框,比‘云抗10號’基因組數據注釋的對應CDS序列(CSA035098.1)長615 bp,且有17個堿基位點不同。這些堿基位點的不同是否由于‘舒茶早’與‘云抗10號’之間的物種差異造成的,還需要進一步研究。與2018年公布的‘舒茶早’茶樹基因組[21]中對應的STR基因(TEA012295.1)CDS序列相比,除在912位有個A與T堿基的顛換,在299 bp后有258 bp的插入,其余序列與該STR基因(TEA012295.1)的CDS序列完全一致。進一步分析顯示,CsSTR1基因CDS序列與新公布的‘舒茶早’高質量基因組數據庫中對應STR基因(CSS0030692.1)的CDS序列完全一致,表明本研究RACE克隆的準確性。同時,通過RACE克隆,本研究還得到CsSTR1基因的5′ 和3′ UTR序列,為設計CsSTR1基因特異引物進行基因表達研究以及CsSTR1基因上游調控因子的分離鑒定和調控研究奠定了基礎。
CsSTR1蛋白三維結構預測分析表明,CsSTR1蛋白呈六葉β-螺旋漿狀;亞細胞定位預測分析表明,CsSTR1定位于液泡,這與已有的研究一致[5]。系統進化分析顯示,CsSTR1與水稻OsSTRL2的同源性最高,而與蘿芙木、蛇根木、長春花等植物中的STR同源性較低,暗示CsSTR1可能不具有催化異胡豆苷合成的功能。研究表明,OsSTRL2主要在花器官中表達,在花藥發育和花粉形成中有重要作用,OsSTRL2基因敲除突變導致水稻雄性不育;另外,OsSTRL2突變體還呈現出花藥外表皮上不正常的蠟質晶體,可能還影響了水稻對病原菌的抗性[13]。CsSTR1基因在芽和幼葉中表達量較高,而在花和果中表達量較低。表明CsSTR1與OsSTRL2的表達調控不完全一致。CsSTR1可能在茶樹芽葉發育和病蟲害防御反應中發揮作用。根據最近的研究結果,在利用更大范圍STR蛋白構建的系統進化樹中,CsSTR1與擬南芥中STR同源蛋白(AT3G57030.1)的親緣關系最近,與水稻OsSTRL2蛋白位于同一進化分枝[18]。研究表明,擬南芥中該STR (AT3G57030.1)參與調控類黃酮物質的合成[22]。茶葉中類黃酮是具有重要健康功效的內含成分,而且是茶樹參與抗逆反應的重要次生代謝物[19, 23]。
STR在應答逆境脅迫和激素信號中有重要作用。CsSTR1基因受低溫和PEG模擬干旱處理誘導表達,結果提示CsSTR1參與茶樹對低溫和干旱的響應。前期研究顯示,STR基因表達受干旱脫水脅迫誘導,但低溫處理下調STR基因表達[8-9]。雖然CsSTR1受低溫誘導表達,但上調表達只出現在低溫處理3 h后,在其他檢測時間段(6~48 h)呈現下調表達。已有研究表明,植物對低溫的應答途徑包括ABA非依賴途徑和不依賴CBF兩條途徑[24]。ABA處理后,CsSTR1基因在24~48 h后顯著下調表達,暗示CsSTR1基因是以ABA非依賴途徑參與對低溫的應答。MeJA和SA處理后,CsSTR1基因都呈現上調表達,這與已有的報道一致[8, 10-11]。MeJA和SA是植物體內重要的信號物質,在調控植物脅迫反應和生長發育中都具有重要作用[25]。MeJA還參與調節植物體內的次生代謝[26]。因此,CsSTR1可能通過參與調控茶樹次生代謝,在茶樹應對逆境脅迫中發揮作用,但其具體的作用機制還需要后續進一步研究。
4 結 論
本研究克隆了茶樹中一個異胡豆苷合酶基因CsSTR1。其CDS長1131 bp, 編碼376個氨基酸,分子量41.5 kD;5′和3′UTR長分別為92和159 bp。CsSTR1具有異胡豆苷合酶Str_synth保守結構域,其三維結構呈六葉β-螺旋漿狀。系統進化分析表明,CsSTR1與水稻OsSTRL2具有較近的親緣關系。CsSTR1在茶樹芽和第1~3葉中表達量較高,在花和果中表達量較低;低溫、PEG、MeJA和SA處理均誘導CsSTR1上調表達,ABA處理后CsSTR1下調表達,表明CsSTR1在茶樹響應生物和非生物脅迫反應中具有重要作用。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
