北極楚科奇海北部特征水團對浮游植物空間分布的調控
2023-01-12 09:02:24費淑怡李紀高源葛躍浩郭正東高詠卉
海洋學報 2023年1期
費淑怡,李紀, ,高源,葛躍浩,郭正東,高詠卉, *
( 1. 上海交通大學 海洋學院,上海 200030;2. 中國極地研究中心 自然資源部極地科學重點實驗室,上海 200136;3. 上海交通大學 海洋學院 上海市極地生命過程與環境重點實驗室,上海 200030;4. 國家海洋局南通海洋環境監測中心站,江蘇南通 226002;5. 中國人民解放軍海軍潛艇學院,山東 青島 266199)
1 引言
北極的氣溫增速約為全球平均值的兩倍,這造成海冰面積減小、多年冰減退、無冰區增加,對脆弱的極地生態環境產生深遠影響[1]。自1978年以來,北極夏季冰蓋面積大幅下降,2021年夏季海冰面積僅為1981-2010年平均面積的77%左右[2]。而且多年冰占總海冰面積的比例從40年前的70%降低至近年來的30%[3],導致冰層逐漸變薄,透光性增強,利于浮游植物生長。此外,海冰開始融化的時間提前,凍結得更晚,無冰期隨之延長[1]。
隨著北極海冰面積減小、透光增強、開放時間延長等變化,北極的水文特性和環流結構也發生變化,對營養物質的輸運、物種的分布、群落結構和生態環境等產生重要影響。北極浮游植物的生物量總體呈增加趨勢[4-5],1998-2012年,海冰面積減小是驅動生物量和生產力增加的主要原因。此后,海冰損失明顯減緩,營養供應增加可能是近10年來的浮游植物生物量增加的主要原因[1]。同時,藻華出現時間更早,秋季二次藻華的報道也有所增加[6-7]。浮游植物通過光合作用吸收CO2,對調節CO2的海氣交換、收支和存儲起著重要作用[2-3,7];作為初級生產者,它們也為魚類、浮游動物和底棲動物提供食物,對生態系統起到關鍵的支持作用[8]。
北極冰下藻華可以從冰邊緣一直延伸至海冰深處[4]。海冰中普遍存在藻類(如海鏈藻Thalassiosira,擬脆桿藻Fragilariopsis, 齒狀藻Odontella等)的休眠孢子,入水萌發,為夏季藻華的形成提供種源。也有少量報道表明,北極水中發現的硅藻能長時間在黑暗中存活[5]。當冰雪融化或者融池形成后,為冰下的浮游植物帶來適宜的光照、削弱紫外線輻射,楚科奇海冰層下的浮游植物就開始生長。冰下寒冷的水溫也造成浮游動物的攝食速率降低[9],這使得冰下區域存在適宜浮游植物生長棲息的區域。浮游植物對環境的適應性有利于它們在高緯度地區的生存,并影響春、夏季藻華中浮游植物的群落結構。
在北極,冰下浮游植物對海洋碳匯的貢獻量約占80%,它們的豐度和分布也造成光合效率、初級生產力和碳通量的區域性差異[3,7]。楚科奇海作為北極氣溫增速最快的海域之一,擴大的無冰區面積和延長的開放時間均居北極的前列,近20年葉綠素濃度和生產力也顯著增加[7]。浮游植物以硅藻為主,葉綠素a濃度(3~800 mg/m3)和初級生產力(以碳計,0.2~463 mg/(m2·d))均呈現明顯的時空變化[5]。然而,過去的研究多集中在楚科奇海南部無冰或少冰區域[10-12],北部邊緣海區的數據不足,缺少對冰下浮游植物及其生存環境的系統性分析。
本文依托中國第12次北極科學考察,對楚科奇海北部邊緣海與加拿大海盆西側(74°~78°N,150°~170°W)進行了生態調查(圖1)。結合水團等環境特征,采樣并分析密集冰區中營養鹽和葉綠素a濃度的分布特征。通過厘清水團分布,探究環境因子變化與浮游植物分布之間的關系,構建北極在環流作用下的物理-化學-生物間的相互作用,有助于評估北極氣候變化對海洋生態系統的潛在影響。
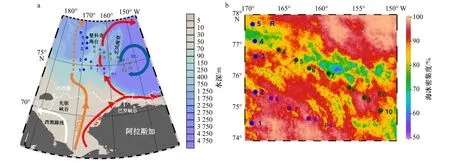
圖1 西北冰洋環流示意圖(a)和采樣區的海冰密集度在調查期間的均值(b)Fig. 1 Schematic diagram of the western Arctic Ocean circulation (a) and the average of sea ice concentration during the sampling time (b)
2 材料與方法
2.1 研究區域
2021年7月25日至8月1日,在北極75°N以北的高緯度冰區的3個斷面開展調查(圖1)。R斷面沿170°W經線,由南向北穿過楚科奇深海高原,其底部為典型的深海海溝和山脊,部分區域呈現平坦的高原狀峰頂。P1和P2斷面大致平行,跨越北風海脊,向東延展至加拿大海盆,水深從200 m驟增至4 800 m(圖1a)。各站位均位于海冰邊緣線以北(圖1,https://seaice.uni-bremen.de/databrowser)。
研究區域位于加拿大海盆邊緣,混合層以下的極地表層水包括夏季、冬季白令海水和阿拉斯加沿岸流(Alaskan Coastal Water,ACW),物理環境復雜[13]。太平洋夏季水(Pacific Summer Water,PSW)流經白令海峽后,攜帶的陸坡沉積物釋放營養鹽,形成營養鹽豐富的白令海夏季水(summer Bering Sea Water,sBSW),向北流向北極的中央海區[14]。PSW的東側支流沿著阿拉斯加沿岸向東北流動。在巴羅峽谷附近,受波弗特高壓和加拿大海盆地形的共同作用,大部分ACW沿加拿大海盆匯入順時針運動的波弗特流渦(Beaufort Gyre, BG)[15];其余沿阿拉斯加陸架繼續向東輸送[16]。由于波弗特高壓強度和位置的改變,BG影響的范圍和強度往往呈現明顯的時空變化[17]。每年的6-7月,阿拉斯加周邊的河流徑流量最大,海冰融水增加,往往造成BG覆蓋區域內的淡水儲量增加[18],BG影響范圍(70°~80°N ,130°~170°W)在8月達到最大[15],形成營養鹽濃度和鹽度略低的ACW暖水團向本研究區域輸送。
2.2 觀測和取樣
利用“雪龍2”號的破冰能力實現冰區采樣。站位水溫和鹽度通過溫鹽深剖面儀(CTD 911,Seabird InC.)測定。水樣在表層、10 m、20 m、50 m、75 m、100 m、150 m、200 m以及葉綠素a濃度最大層(ChlorophyllaMaximum Layer,CML)采集,用于營養鹽和葉綠素a濃度分析。
2.2.1 營養鹽濃度測定
各層采集的水樣經孔徑0.45 μm的醋酸纖維濾膜過濾,冷凍存儲,用于生物可利用溶解態營養鹽的測定。其中,磷酸鹽(SRP)、硅酸鹽(Si (OH)4)、硝酸鹽( NO-3-N)和亞硝酸鹽( NO-2-N)濃度依據《海洋監測技術規程 第1部分:海水》(HY/T 147.1-2013)[19]流動分析法,由Skalar San++營養鹽自動分析儀(荷蘭,SKALAR)測定。氨氮( N H+4-N)濃度依據《海洋監測技術規程 第1部分:海水》(HY/T 147.1-2013)[19]次溴酸鹽氧化法測定。各溶解態營養鹽的檢出限分別為: NO-3-N 0.04 μmol/L、 NO-2-N 0.025 μmol/L、 NH+4-N 0.015 μmol/L、SRP 0.02 μmol/L、Si (OH)40.04 μmol/L。溶解無機氮(DIN)濃度為 NO-3- N、 N O-2- N和 N H+4-N濃度的總和。
2.2.2 葉綠素a濃度測定
依據《海洋監測技術規程 第1部分:海水》(HY/T 147.1-2013)[19]的熒光法測定葉綠素a濃度。先對各站位200 m以上水層的樣品進行粒度分級:量取250 cm3的水樣先經孔徑20 μm的篩絹進行過濾后(分離獲取小型浮游植物細胞,Netphytoplankton),再經孔徑為2 μm的 玻 璃 纖 維 素 酯 濾 膜 過 濾(分 離2~20 μm的微型浮游植物細胞,Nanophytoplankton),最后用Whatman GF/F玻璃纖維濾膜過濾(截留微微型浮游植物細胞,Picophytoplankton)。濾膜冷凍避光保存。以90%丙酮對濾膜上的浮游植物細胞萃取24 h后,在10AU型唐納熒光儀上測定并計算,得到Chla濃度[20]。
2.3 數據分析
運用SPSS 26.0軟件對數據進行統計學分析。R1、R3站位和其他站位之間的環境因子差異采用一元方差分析(One-way ANOVA)進行分析。環境因子和分級葉綠素濃度之間的相互關系用Spearman相關系數來表示。
3 結果
3.1 海區冰情、水文分布及水團特征
自1978年以來,雖然北極夏季冰蓋面積的總體變化趨勢大幅下降,但2021年夏季海冰覆蓋面積略高于2015-2020年[2]。調查期間,高緯度冰區剛開始融化。受西北風影響,海冰向研究區域移動和堆積。海冰密集度的均值約為75%,冰厚往往高達1 m左右。除了沿P2斷面零星出現海冰密集度約50%的區域,絕大多數區域由冰覆蓋(海冰密集度為90%~100%,圖1b)。
冰下0~150 m,水溫介于-1.75~0.8℃之間,鹽度為27~35,呈現復雜的層化現象(圖2)。海冰融化、混合形成低鹽的融冰水(Melting Water,MW)。融冰水以下,根據Steele等[14]和Wang等[21]對楚科奇海和加拿大海盆地區中夏季鹽躍層的劃分,可分為ACW和sBSW兩個暖水團,ACW水溫往往略高于sBSW。向下依次為低溫的白令海冬季水(winter Bering Sea Water,wBSW)和高溫高鹽的大西洋水(Atlantic Water,AW)。
本次研究區域存在相似的垂向和水平結構(圖2)。融冰水使表層形成水溫接近于海水冰點(-1.7℃)、鹽度小于30的低密度水層,從表層延伸至20 m上下。融冰水以下,鹽度隨深度增加而逐漸增大,溫度垂向分布存在明顯差異。在次表層(30~100 m),3個斷面均形成狹長的溫度略高的水舌。在R斷面北部、P1斷面、P2斷面(8-10站位)尤為明顯,水溫向加拿大海盆邊緣方向逐漸降低,該暖水團(溫度:-1.25~0.75℃,鹽度:30~32)符合受BG驅動的ACW特征。在R斷面南部的次表層(50~100 m)為sBSW暖水團(中心溫度約-1.5℃,鹽度為31~32),核心溫度呈隨緯度升高而降低的趨勢。在較冷的wBSW(70~125 m)下方,出現高溫高鹽的AW水團(圖2,表1)。
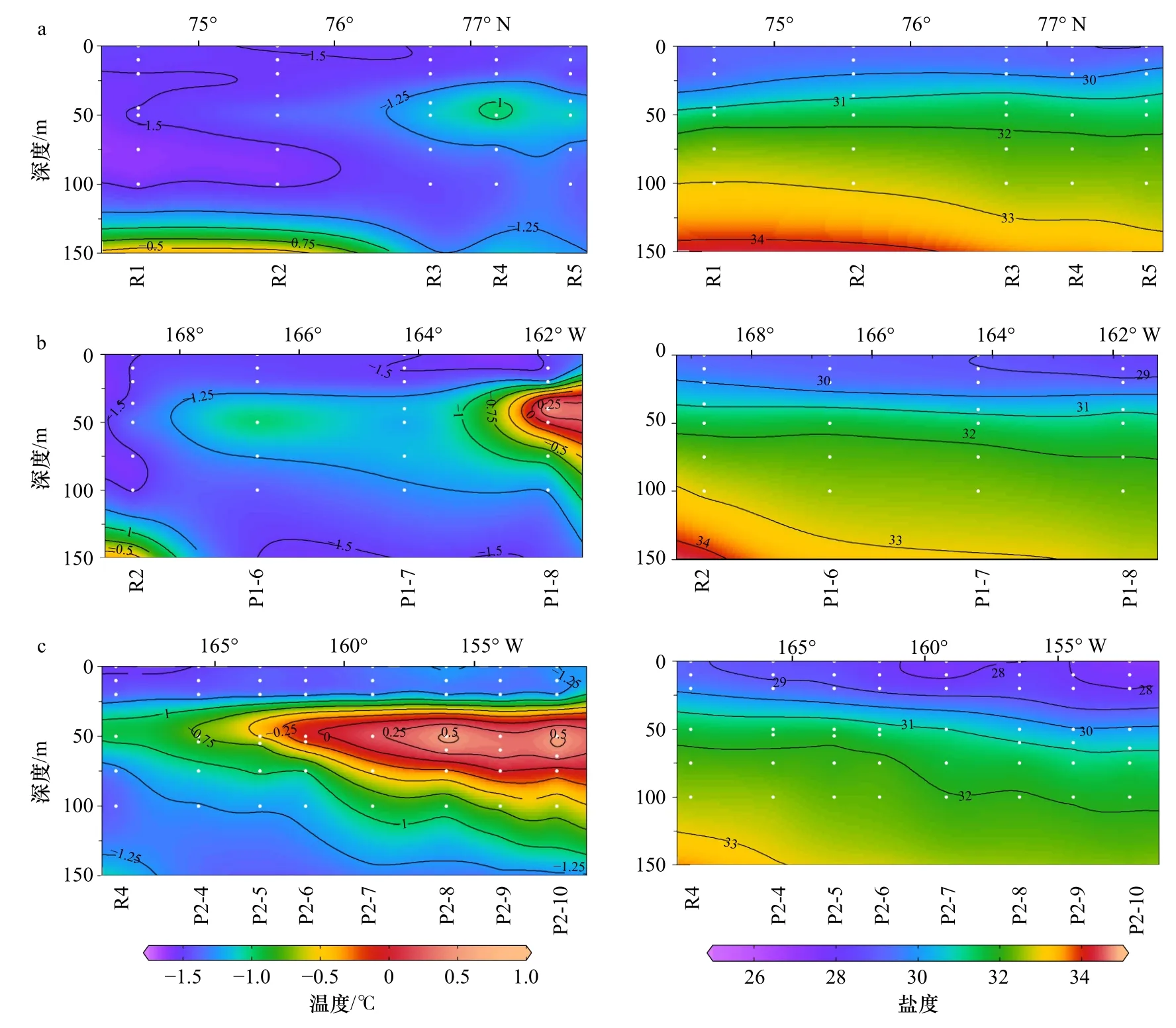
圖2 在 R(a)、P1(b)和 P2(c)斷面水溫和鹽度的垂向分布Fig. 2 Vertical distribution of temperature and salinity in the R (a), P1 (b), and P2 (c) transections
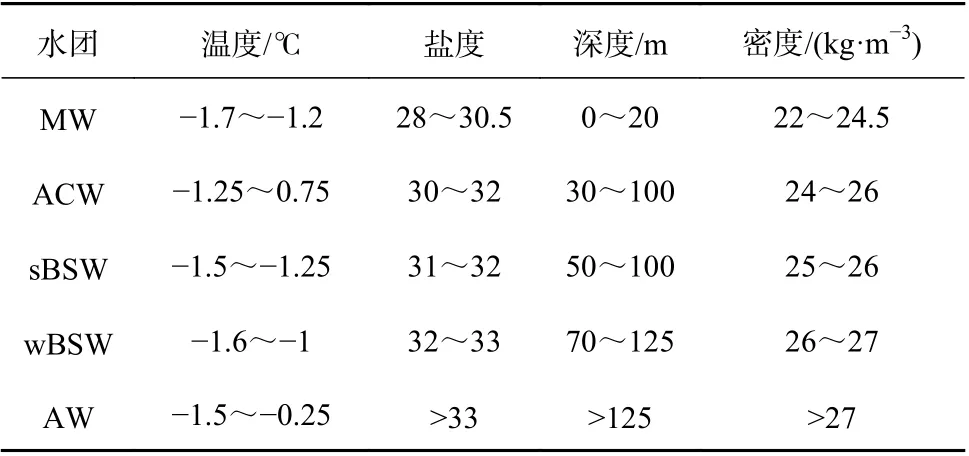
表1 不同水團溫度、鹽度、密度的分布Table 1 Distribution of temperature, salinity and density in different water mass
3.2 營養鹽的分布
生物可利用溶解態營養鹽的濃度變化范圍分別為DIN 0.47~18.19 μmol/L,SRP 0.23~2.65 μmol/L和Si(OH)41.08~39.06 μmol/L(圖3)。總體而言,營養鹽濃度隨著深度增加而逐漸升高。在0~20 m的MW中生物可利用的營養鹽濃度最低(DIN濃度<2.5 μmol/L、SRP濃度<1.5 μmol/L、Si(OH)4濃度<5 μmol/L)。50~100 m為營養鹽濃度的躍層,營養鹽的濃度均隨深度迅速增加。受BG影響,在P1、P2斷面呈現DIN等營養鹽躍層從深海向加拿大海盆邊緣地帶(從東向西)逐漸變淺的現象。R斷面(1~3站)在sBSW影響下,自南向北營養鹽濃度逐漸降低。100 m以下營養鹽濃度普遍較高,在靠近AW的125~150 m出現營養鹽濃度的最高值。DIN濃度與SRP濃度之比多低于16,存在氮的相對限制。
3.3 葉綠素分布
Chla的濃度為0~0.69 mg/m3,垂向變化明顯(圖4)。分布規律與水團的密度層化一致性良好。低營養鹽的表層MW中的Chla濃度低,部分區域幾乎為0(圖3,圖4)。在40~60 m的次表層暖水團區域內,尤其是ACW和sBSW的上邊緣,往往出現葉綠素a濃度最大值(平均值±標準差=(0.327±0.163)mg/m3)。Chla濃度在R斷面普遍高于P1和P2斷面(圖4)。在P1斷面,Chla濃度從西向東向加拿大海盆邊緣逐漸增加。在60 m以下,光強隨著深度增加而減弱,Chla濃度也逐漸降低,100 m以下Chla濃度幾乎為0。
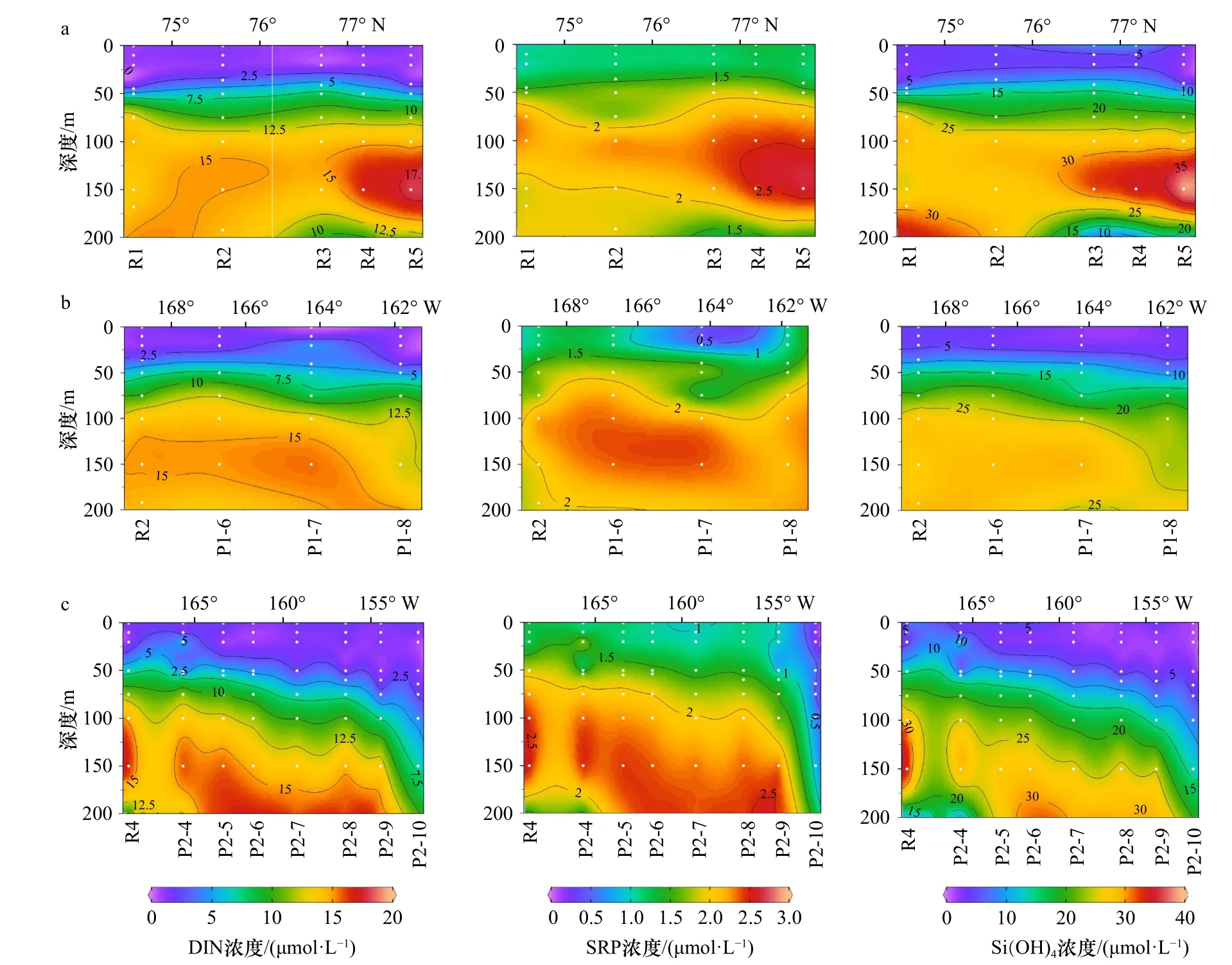
圖3 溶解態無機氮(DIN)、磷酸鹽(SRP)和硅酸鹽(Si(OH)4)濃度在 3 個斷面的垂向分布Fig. 3 Vertical distribution of dissolved inorganic nitrates (DIN), soluble reactive phosphates (SRP) and silicates (Si(OH)4) concentration in three transections
結合垂向分布(圖2至圖4)和溫-鹽-深的點聚圖(圖5a,表1),可以看出該區域的層化現象除了長期存在的白令海冬季水(wBSW)和深層的大西洋水(AW)外,主要受融冰水(MW)、阿拉斯加沿岸流(ACW)和白令海夏季水(sBSW)入侵水團的影響。以對浮游植物生長有潛在限制的DIN為例,結合溫-鹽-葉綠素a濃度(圖5b,圖5c)進行點聚分析:DIN濃度在MW中最低(0~2.5 μmol/L之間)。在次表層暖水團中,DIN在ACW水團中濃度近似于sBSW水團,與葉綠素a濃度高值區相吻合。在125 m以下的AW水團中則呈現高DIN濃度和低Chla濃度的特征。
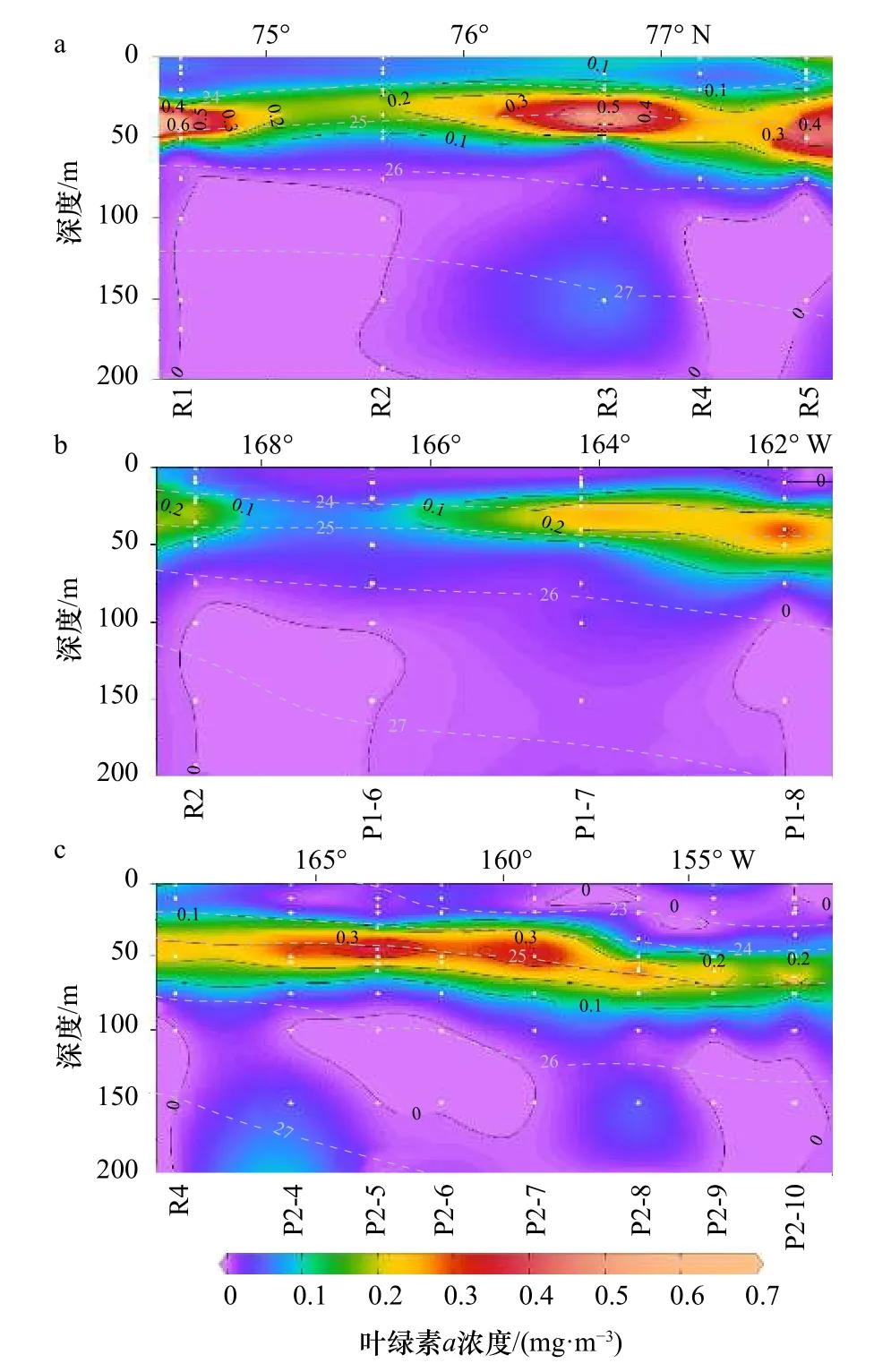
圖4 在 R(a)、P1(b)和 P2(c)斷面葉綠素a濃度的垂向分布(灰色虛線為等密度線)Fig. 4 Vertical distribution of chlorophyll a concentration in the R (a), P1 (b), and P2 (c) transections (gray dotted lines are equal density lines)
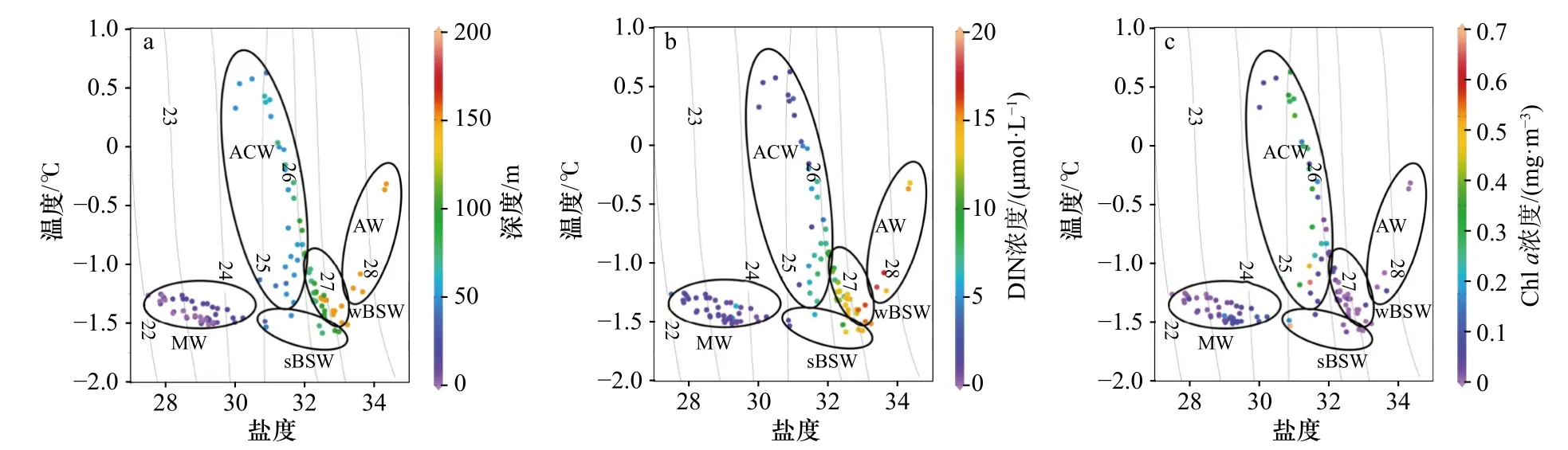
圖5 150 m 以上水體的溫-鹽-深度點聚圖(a)、溫-鹽-DIN 濃度點聚圖(b)和溫-鹽-葉綠素 a濃度點聚圖(c)Fig. 5 Depth distribution in temprature-salinity-depth scatter diagram (a), temperature-salinity-DIN concentration scatter diagram (b)and temperature-salinity-Chl a concentration scatter diagram (c) above 150 m
3.4 不同粒徑葉綠素a濃度的分布及其與環境因子的關系
各站位葉綠素a濃度最大層處的分級Chla濃度分布如圖6所示。sBSW影響下的R1、R3站位小型浮游植物葉綠素a濃度占比70%以上,這兩個站位的溫度明顯低于其他站位(One-way ANOVA,p=0.053<0.1)。在其他站位,浮游植物以微微型為主,微微型浮游植物Chla濃度范圍為0.03~0.28 mg/m3,占總葉綠素a濃度的40%~80%(均值約為65%)。微型浮游植物葉綠素a濃度在所有站位中比例均低于30%。
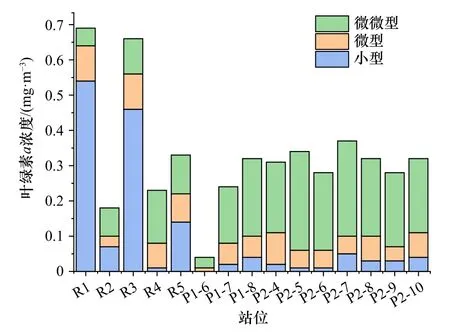
圖6 葉綠素a濃度最大層的浮游植物葉綠素a濃度的分級分布Fig. 6 Size-fraction distribution in Chl a concentration of phytoplankton in Chl a maximum concentration layer
各粒級浮游植物葉綠素a濃度與環境因子的Spearman相關性分析(表2)表明:小型浮游植物Chla濃度受多種環境因子的影響,與鹽度、DIN濃度、SRP濃度、Si (OH)4濃度和N/P值都呈顯著性負相關;微微型浮游植物Chla濃度與溫度呈顯著正相關,受鹽度和營養鹽的影響較弱。
4 討論
4.1 環境因子對浮游植物的影響
葉綠素濃度的垂向分布與2003年[20]、2008年[22]我國在北風海脊附近的觀測相似。由于太陽輻射受海冰阻擋,海水中光照隨深度增加而減弱。各站位的葉綠素a濃度最大層位于50 m上下,光合有效輻射(PAR)范 圍 為1~27 E/(m2·d),平 均PAR為(8.60±8.08)E/(m2·d)。雖然葉綠素a濃度最大層光較弱,但是越來越多的觀測結果揭示了北極浮游植物在冰下和極夜的低光照環境中,以消耗脂類、碳水化合物和蛋白質來提供能量,降低代謝速率和細胞分裂等機制,維持Chla濃度增長和光合作用,形成了獨特、長期的暗適應能力[23-25]。浮游植物在PAR小于10 E/(m2·d)受 到 光 限 制,但 仍 能 在PAR低 至1 E/(m2·d)的環境下存活[6,23-24]。葉綠素a濃度最大層處微弱的光強也能支持浮游植物生長。由此可見(表2),光照對葉綠素a濃度最大層處浮游植物的分級Chla濃度的影響不顯著。
浮游植物的葉綠素a濃度最大層與營養鹽的分布密切相關。雖然表層光照更適宜浮游植物生長,但寡營養的融冰水在表層融冰層造成營養鹽(尤其是氮)的限制。太平洋入流水中經阿拉斯加沿岸進入加拿大海盆的ACW,富含營養鹽、相對溫暖,在次表層創造了相對適宜浮游植物生長的環境。在葉綠素a濃度最大層處,DIN∶SRP在1.0~4.1之間,而Si(OH)4∶DIN在1.9~3.7之間。與Redfiled Ratio(C∶N∶P∶Si=106∶16∶1∶16)相比,雖然N存在相對限制,但仍高于限制浮游植物生長的閾值(0.1 μmol/L)[26-27],對浮游植物不構成絕對限制。在葉綠素a濃度最大層處,浮游植物中耐受性強的物種會對N產生明顯的生物吸收。
從浮游植物粒徑分布來看,除了R1和R3站位的小型浮游植物占比較高(約70%)外,微微型浮游植物在冰區占主導地位。這一結果與Joo等[28]在2008年楚科奇海夏季浮游植物中的發現相符。Smith[29]也發現浮游植物的粒徑與冰覆蓋面積和營養鹽濃度有關:在冰覆蓋超過50%的海區浮游植物以微微型為主,而在層化強、營養鹽充沛的區域則以小型浮游植物為主。由Spearman分析(表2)可以看到,微微型浮游植物多出現在溫度略高、營養鹽濃度較低的水層,并成為該區域的優勢種。在低溫但營養鹽濃度較高的R斷面,以R1和R3為代表的站位中,小型浮游植物占比較高,形成了以小型浮游植物為主的浮游植物群落。這些粒徑不同的浮游植物群落在北極生態系統的群落結構和營養關系中起著重要作用。微微型浮游植物的粒徑更小,具有更大的比表面積,代謝和能量轉換效率高,通過冰下和融冰初期生物碳吸收、食物鏈間的傳遞利用和生物泵作用,在楚科奇邊緣海區的物質循環與能量流動中起著不可替代的作用[20]。
4.2 生物作用對葉綠素a濃度最大層處營養鹽去除的影響
本文通過水團分析發現葉綠素a濃度最大層與太平洋入流水的傳輸和區域性分布密切相關。葉綠素a濃度最大層中,不同粒徑的Chla濃度與營養鹽濃度呈負相關(表2),浮游植物對營養鹽的吸收造成了營養鹽濃度降低。
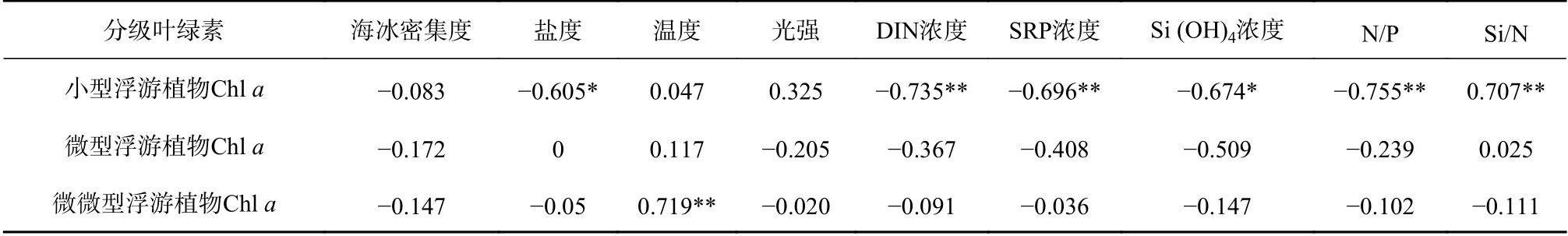
表2 分級葉綠素a濃度與環境因子的相關性Table 2 Correlations of size-fraction chlorophyll a concentration and environmental factors
為區分水團混合和生物作用對營養鹽濃度的改變,在接近葉綠素a濃度最大層,營養鹽吸收較旺盛的50 m處作圖(圖7)。由于在氮限制的水體中,DIN濃度與鹽度的關系優于SRP濃度與鹽度的關系。假如在ACW與sBSW水團物理混合的理想狀態下,DIN濃度與鹽度保守混合線呈線性關系[30]。然而,實測DIN濃度與鹽度關系呈凹形,并且DIN濃度低于混合線(虛線)。一般而言,浮游植物對營養鹽吸收量越高,DIN缺失量(ΔDIN)越高。這從葉綠素a濃度分布看,P2斷面與R斷面相差不大,P1斷面濃度較小(圖4), 而在對氮去除效率上,P1斷面與R斷面相差不大,但是稍遜P2斷面。受ACW暖水團影響的P1和P2斷面,Chla濃度與ΔDIN成正比。因而,浮游植物的豐度越高,對營養鹽吸收越強。由于ACW平均溫度高于sBSW,浮游植物的氮吸收效率往往隨溫度升高而增強。這導致ΔDIN在Chla濃度近似的P2斷面均高于R斷面,ΔDIN在低Chla濃度的P1斷面近似于R斷面(圖7)。這也進一步證明了在以小型浮游植物為主的R斷面,其浮游植物的營養鹽吸收效率低于以微微型浮游植物為主的P斷面。
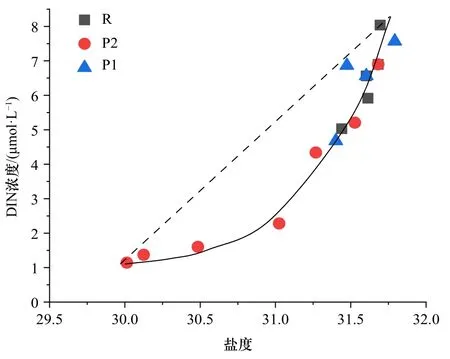
圖7 溶解無機氮(DIN)濃度與鹽度關系Fig. 7 Relationship between dissolved inorganic nitrate (DIN)concentration and salinity
4.3 浮游植物種群結構
在Ekman輸送和地轉力的共同作用下,BG上層的密度結構呈現中央低四周高的形態,營養鹽濃度從外圍向環流中心降低[16]。Zhuang等[31]也報道,從BG邊緣到中心區域,以硅藻為主的生物量逐漸減少。這與我們在P斷面觀測到的營養鹽濃度和藻類豐度從BG西邊緣區域向中心減低的空間分布趨勢相吻合。以P2斷面東側(8~10站位)為例,營養鹽濃度的等密度線逐漸向下延伸,暖水ACW和冷水wBSW間的層化加劇,提高了氮限制對藻類生長的脅迫效應。營養鹽濃度在真光層的降低,不利于浮游植物生長,導致Chla濃度從西向東逐漸降低(圖3)。浮游植物粒徑越小,在DIN限制下的適應能力越好[9,32],因而在BG影響范圍內,以微微型浮游植物為主。
4.4 浮游植物的發展趨勢
在北極變暖和北極放大效應的驅動下,北極生態環境趨向于向利于海洋浮游生物生存與繁衍的方向演變[33-34]。在近10年間,浮游植物的葉綠素a濃度年均增速約為0.013 mg/m3,初級生產力(以碳計)增速約為7 Tg/a[7]。BG海洋熱含量在過去的30余年間增加1倍,加速了海冰消融,對生物地球化學過程產生了重要影響[35]。此外,浮游植物增加與營養鹽供給增加密切相關。一方面,太平洋入流水持續增加,增加了sBSW攜帶的營養鹽輸入[36]。另一方面,融冰促進了阿拉斯加沿岸河流和陸源的有機物輸入。BG淡水含量和洋流速度也穩步增加[37],比20世紀70年代升高了40%[38],進而增加了ACW向海盆區的營養鹽供給[39-40]。以上原因都將緩解北極營養鹽限制,推動浮游植物生物量的進一步提高[28]。
在本次調查的高密度冰區,我們發現葉綠素a濃度高于同地區的歷史記錄。劉子琳等[22]在2008年中國第三次北極考察時,測得楚科奇海北部區域的Chla濃度在0.004~0.468 mg/m3間變化。其最大值低于本次考察的Chla濃度最大值(0.69 mg/m3)。我們也對研究區域內2003-2016年間國際公開數據統計分析(航 次:32H120030911、32H120040910、49NZ20020-822、49NZ20040901、18SN20110721、18SN20120802、18SN20130724、18SN20140921、18SN20160922,https://explore.webodv.awi.de/ocean/hydrography/glodap/),發現該區域13年間的Chla濃度范圍為0.025~0.54 mg/m3,平均濃度為(0.126±0.093)mg/m3。歷史數據的Chla濃度最大值低于本次觀測,平均值約為本次觀測結果((0.327±0.163)mg/m3)的一半。該區域浮游植物的增長與北極圈內的變化趨勢相一致[7]。從采樣時間看,公開數據和劉梓琳等[22]的數據多在8-9月獲取,本次調查采樣早,冰覆蓋下的藻華仍處于生長初期。預計隨氣溫季節性升高,海冰融化,光限制削弱和無冰區擴大,浮游植物生物量將更高[1,41]。這意味著隨著氣候的變化,以浮游植物對碳吸收為主的生物碳匯將呈現增加的趨勢,有利于北極作為碳匯區域的發展。
5 結論
(1)在楚科奇北部海區-加拿大海盆西側海區,MW、ACW和sBSW等水團通過調控營養鹽濃度和溫度等因素,間接影響了浮游植物的分布和群落結構。
(2)葉綠素a濃度在40~50 m處的次表層暖水團(ACW、sBSW)中達到最大值,濃度在0.04~0.69 mg/m3之間,平均為(0.327±0.163)mg/m3。ACW相對于sBSW而言,水溫高但營養鹽濃度略低。由于浮游植物的環境適應性,在ACW中以粒徑小于2 μm的微微型浮游植物為主(約65%),在sBSW影響的站位中,以粒徑大于20 μm的小型浮游植物為主(約74%)。
(3)營養鹽濃度、溫度等因素對浮游植物的分布和群落結構影響較大。受北極極夜和冰下弱光影響,冰下浮游植物在長期演變中的暗適應性較強,光強對浮游植物分布的影響較弱。盡管融冰表層光照強,但寡營養鹽和低溫的表層冰融水會限制微藻生長。浮游植物在營養鹽相對充裕和溫度較高的ACW和sBSW暖水團附近(約50 m)聚集。根據保守混合模型和環境因子分析,葉綠素a濃度最大層中浮游植物的豐度和群落結構影響了DIN的吸收量和分布。
(4)浮游植物粒徑越小,在DIN限制下的適應能力越好。在BG影響范圍內,DIN限制明顯,浮游植物多以微微型為主。
(5)在海冰融化初期,葉綠素a濃度高于以往的報道。這與北極變暖、融冰加劇、BG持續增強、太平洋入流水攜帶的營養鹽增加等利于浮游生物生長的因素密切相關。浮游植物的增加有助于碳吸收,有望促進北冰洋碳匯增加。
猜你喜歡
海洋通報(2021年3期)2021-08-14 02:20:38
今日農業(2021年11期)2021-08-13 08:53:34
成都信息工程大學學報(2021年2期)2021-07-22 07:23:32
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
作文周刊·小學一年級版(2017年17期)2017-06-27 19:32:55
電子技術與軟件工程(2016年24期)2017-02-23 19:13:41
廣東第二課堂·小學(2017年2期)2017-02-20 15:10:08
海峽姐妹(2016年5期)2016-02-27 15:20:20
中國學術期刊文摘(2016年8期)2016-02-13 13:04:44
