昆蟲神經肽tachykinin研究進展
2023-02-03 10:07:00劉曉光徐夢超王青鴿蘇健誠魏紀珍安世恒
植物保護 2023年1期
關鍵詞:研究
劉曉光, 徐夢超, 王青鴿, 蘇健誠, 魏紀珍, 安世恒
(小麥玉米作物學國家重點實驗室, 河南省害蟲綠色防控國際聯(lián)合實驗室, 河南農業(yè)大學植物保護學院, 鄭州 450002)
昆蟲體外信息交流和體內信號傳遞均離不開神經肽的參與[1-7]。神經肽是一類小分子活性多肽,主要由神經元(neurons)、神經內分泌細胞(neuroendocrine cells)或腸內分泌細胞(endocrine cells)分泌,其生成過程具有較高的相似性(圖1),即神經肽基因經過轉錄、翻譯后首先形成前體蛋白(precursor proteins,pro-proteins),前體蛋白在其N端具疏水結構的信號肽的引導下進入內質網(endoplasmic reticulum,ER)[8]。在內質網中,前體蛋白N端信號肽首先被切除,切除信號肽的其余肽段經歷一系列翻譯后修飾(糖基化、羥基化、N端酰基化和二硫鍵的形成)進入高爾基體,其序列中某些單個或者相鄰兩個位點氨基酸殘基被特異性蛋白酶識別并切割(C端堿性殘基由羧肽酶轉化酶切除形成C端酰胺化或N端焦谷氨酰殘基發(fā)生谷氨酰胺酰基環(huán)化)[9],最終形成具有生物活性的成熟多肽,這些活性多肽通過各種途徑分泌到細胞外,最終達到靶位點,與G 蛋白偶聯(lián)受體結合(G protein-coupled receptor, GPCR),激活下游信號通路[8,10]。隨著基因組、轉錄組和多肽質譜等技術的快速發(fā)展,諸多神經肽得以發(fā)現(xiàn)。僅昆蟲中就有30余種、數(shù)百個成熟肽得到鑒定[11-15]。其中tachykinin(TK)為眾多神經肽中的一類,也稱速激肽。TK 在進化上屬于較為保守的腦-腸多肽(brain-gut peptide),參與生物的生長發(fā)育和生理行為活動[16-18]。TK含量雖少,但在昆蟲生長發(fā)育過程中卻起著異常重要的作用,對于以TK為代表的神經肽的深入研究,有助于人們更全面地了解昆蟲生理功能及內分泌機制,為探索害蟲治理提供科學依據(jù)。

圖1 昆蟲速激肽成熟肽合成釋放過程
1 昆蟲速激肽的發(fā)現(xiàn)
無脊椎動物中,TK最早發(fā)現(xiàn)于昆蟲,Schoofs等收集并解剖9 000多頭飛蝗Locustamigratoria腦-心側體-咽側體-咽下神經節(jié)復合體,通過分離純化,首次獲得TK的多肽序列[19-20]。利用類似方法,Muren等從1 000頭馬德拉蜚蠊Leucophaeamaderae的腦組織中成功分離純化出7個TK多肽[21]。此后,研究者又陸續(xù)從黑腹果蠅Drosophilamelanogaster[22]、家蠶Bombyxmori[23]、赤擬谷盜Triboliumcastaneum[12]、豌豆蚜Acyrthosiphonpisum[24]、褐飛虱Nilaparvatalugens[25]、吸血蝽Rhodniusprolixus[26]、意大利蜜蜂Apismellifera[13]、二化螟Chilosuppressalis[14]、麥蛾繭蜂Habrobraconhebetor[15]和稻綠蝽Nezaraviridula[27]等其他昆蟲中發(fā)現(xiàn)。
2 速激肽的命名與分類
分類命名上,在脊椎動物中發(fā)現(xiàn)的速激肽稱為典型的P物質(substance P,SP),主要包括神經激肽A(neurokinin A,NKA)和神經激肽B(neurokinin B, NKB)[28];在無脊椎動物包括昆蟲中發(fā)現(xiàn)的稱為TK或速激肽相關肽(tachykinin-related peptide, TKP),它們共同構成了速激肽超基因家族[29]。
3 速激肽的一般特征
目前研究結果表明,在編碼昆蟲TK蛋白的序列中一般含有多個TK成熟多肽,而單個成熟多肽通常由8~14個氨基酸殘基組成,且C末端均發(fā)生酰胺化。如利用分離純化方法,結合質譜技術研究者從飛蝗中鑒定出4種TK成熟肽[19-20],果蠅中有5種[30],家蠶和小地老虎Agrotisipsilon中有6種[23,31-32],意大利蜜蜂中鑒定出7種[13,33-34],赤擬谷盜中預測出8個[12],褐飛虱中預測出8個[25]。不同昆蟲體內TK成熟肽數(shù)量不盡相同,但它們均由一個基因開放閱讀框(open reading frame,ORF)編碼的前體蛋白產生。大多數(shù)昆蟲TK前體蛋白中,編碼其成熟肽的氨基酸序列有一定的差異,但也有一些昆蟲中存在一個前體蛋白所產生的若干個TK成熟肽氨基酸序列完全一致的現(xiàn)象(如意大利蜜蜂TK前體蛋白序列中有兩個位點同時產生APMGFQGMR-amide成熟肽),不同昆蟲產生的TK成熟肽也有個別序列完全一致,如馬德拉蜚蠊與赤擬谷盜中均存在APSGFMGMR-amide成熟肽。這些多肽的鑒定和預測也為發(fā)現(xiàn)和研究相關受體提供了很好的基礎。其次,除酰胺化外,這類成熟肽在 C端也相對保守,具有典型的FX1GX2R氨基酸基序(圖2)。
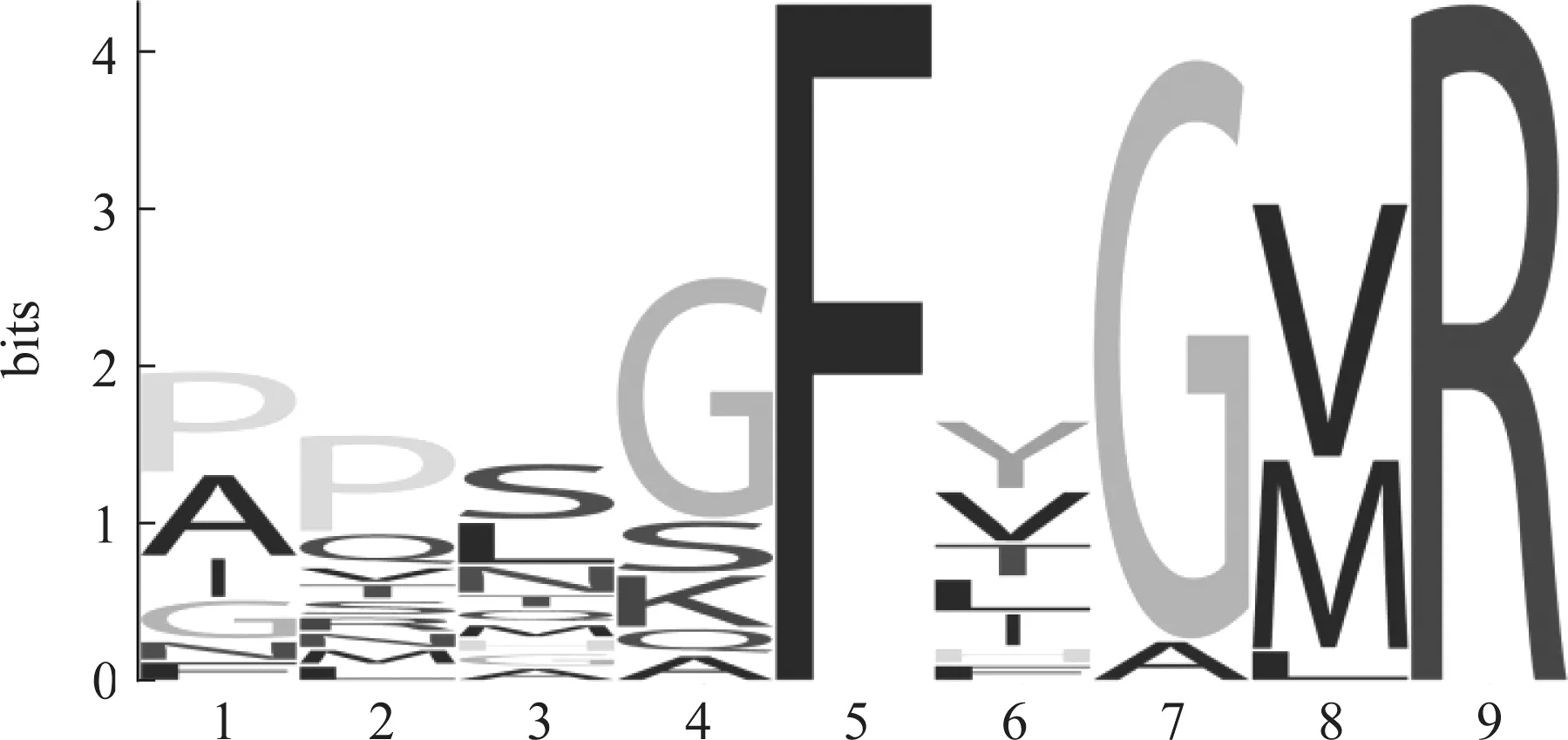
圖2 昆蟲速激肽成熟肽C端典型FX1GX2R基序
4 速激肽分離、鑒定和預測方法
昆蟲神經肽的分離與鑒定歷經了較為漫長的發(fā)展過程。自Stone等從飛蝗和沙漠蝗Schistocercagregaria心側體(corpus cardiacum)中分離并首次測序了神經肽脂動激素(adipokinetic hormone,AKH)開始[35],至Schoofs等成功分離并測定出TK部分序列,開啟了有關昆蟲TK研究的新征程,這也是在節(jié)肢動物中最早發(fā)現(xiàn)TK的正式報道,該傳統(tǒng)研究方法沿用至今[19-20]。
在利用傳統(tǒng)方法研究TK等神經肽過程中,樣品前處理、活性組分的收集與分離等是獲取活性神經肽的關鍵和難點。前處理過程需要兩步,首先,組織解剖后樣品需要盡快進行熱變性或立即采用有機溶劑(如冰凍的色譜級甲醇)進行變性穩(wěn)定處理,或液氮迅速冷凍后暫時存放于-80℃,防止成熟肽的快速降解[36];其次是樣品除雜,樣品組織需經超聲破碎、離心后取上清液,上清液經超濾柱(膜)初步去除大分子蛋白。以上預處理樣品再借助填充不同材料的色譜柱分離,根據(jù)單一峰或若干組合峰對應的液相分離液,準確收集并進一步將其冷凍濃縮獲得活性組分。此步是獲得活性神經肽的關鍵,技術上有一定的難度。因此,利用色譜柱分離是多肽分離技術的核心,其在實際應用中經歷了較大改進與發(fā)展。該技術最初采用可控孔徑玻璃珠(controlled-pore glass,CPG)作為吸附載體,先將小分子多肽過柱分離、濃縮,再經過硅膠薄層層析板(silica gel thin-layer plate)分離獲得微量(μg)、純度較高且具有活性的神經肽[35];隨后又發(fā)展到利用高效液相色譜(highly performance liquid chromatography,HPLC)系統(tǒng),與不同非極性反相(reversed-phase)色譜柱組合,充分利用色譜柱極性差異,不斷更換色譜柱,依次反復純化洗脫。洗脫粗組分經生物活性測定初步確定多肽分離的保留時間(每種多肽具有特殊的生理功能,可通過生物活性測定初步鑒定多肽種類)[37];進一步地,學者們又將免疫學與色譜技術結合,用神經肽(TK等)多克隆抗體結合放射免疫(radioimmunoassay,RIA)[2]或酶聯(lián)免疫(enzyme-linked immunosorbent assay,ELISA)等技術[4]快速確定陽性組分,同時記錄具有陽性組分所對應的更準確的保留時間,利用同樣分離條件可在短時間內獲得大量分離液,將收集到的分離液進一步冷凍濃縮,提高天然神經肽的獲得率與產量,完善了TK神經肽等分離和鑒定技術[36]。由于該技術操作繁瑣,對儀器及操作過程要求較高,目前國際上僅有少數(shù)實驗室仍保留著以上傳統(tǒng)的分離鑒定方法。此外,還可借助簡單的反相色譜技術,前處理樣品經過一次反相色譜分離得到粗產物,該粗產物在快速冷凍濃縮后無需經過其他色譜填料的反相色譜柱組合進行反復地分離,即可直接上質譜系統(tǒng)檢測[32],還有將神經組織或腦組織切片直接上質譜獲得TK精確分子量的報道[33-34]。此外,隨著生物信息學和質譜技術的飛速發(fā)展,首先利用以上質譜技術,獲得多肽的精確分子量,或者利用同位素標記相對和絕對定量(isobaric tags for relative and absolute quantification, iTraq)技術獲得序列確定的多肽片段,再結合基因組、轉錄組,將收集到的組織樣品與相應的質譜數(shù)據(jù)與氨基酸序列自動匹配,預測出神經肽成熟肽序列[32]。
5 速激肽的表達與分布
昆蟲TK與其他神經肽類似[1],最早從腦組織(腦-咽側體-心側體-咽下神經節(jié))中分離[19-20],后來也在中腸發(fā)現(xiàn),因此又稱腦-腸肽[2]。有關TK的研究當數(shù)黑腹果蠅中最為詳細,Veenstra等利用埃及伊蚊AedesaegyptiTK多肽制備的抗體,在黑腹果蠅成蟲不同組織中進行了TK神經元定量分析,結果發(fā)現(xiàn)腦部最多,約700個,中腸中次之,約360個,其次是后腸,約140個,胸腹神經約110個,馬氏管約10個,嗉囊最少,約4個[38]。類似地,有關TK神經元在煙草天蛾Manducasexta中的分布也有報道。在煙草天蛾成蟲期,檢測到TK神經元主要集中在中腦(midbrain),約有100個,其中視神經葉(optic lobe)中僅發(fā)現(xiàn)1個。而在5齡幼蟲中,TK神經元主要集中在腦神經細胞,約有60個;其次是腹神經索(ventral nerve cord),其中咽下神經(suboesophageal ganglion)5對,胸神經(thoracic ganglia)2對,第5未融合的腹神經(fifth unfused abdominal ganglia)各1對,融合末端神經(fused terminal ganglion)3對,與黑腹果蠅成蟲不同,在煙草天蛾成蟲與幼蟲的前腸和后腸中并未檢測到TK神經元[39]。此外,借助新興的單細胞轉錄組測序技術,Guo等首次研究了黑腹果蠅中腸腸內分泌細胞各類神經肽的分布特點,其中重點關注了TK神經元。結果表明,在中腸共富集并分離鑒定出10類腸內分泌神經元類群, 其中TK集中表達于4個主要細胞類群(即所分屬的1、3、4和7腸內分泌細胞類群)。同時發(fā)現(xiàn),TK成熟多肽具有與不同神經肽共表達的偏好性,即TK偏好在某些內分泌細胞中單獨存在,或與DH31、CCAP、CCH、ITP、Gbp5、sNPF、NPF、Mip、Nplp2等單個或多個神經肽共表達,而極少或決不與神經肽AstC共表達[40]。
6 速激肽受體研究
盡管昆蟲TK不斷被發(fā)現(xiàn),但有關其受體的發(fā)掘和研究較少,目前已發(fā)現(xiàn)的潛在受體可歸為3類。Li等利用哺乳動物同源TK受體(tachykinin receptor,TKR)基因作為探針,從黑腹果蠅cDNA文庫中篩選并首次發(fā)現(xiàn)了第一類無脊椎動物TK受體,該受體由CG7887基因編碼,也稱為DTKR(Drosophilatachykinin receptor)或TKR99D[41]。第二類TK受體由CG6515基因編碼,起初命名為果蠅神經激肽受體(neurokinin receptor fromDrosophila),簡稱NKD或TKR86C。這兩類與哺乳動物神經激肽受體(mammalian neurokinin receptor)跨膜區(qū)序列有32%~48%的相似度[42]。第三類速激肽受體是從廄螯蠅Stomoxyscalcitrans中克隆獲得,稱為STKR,它與前兩類受體在跨膜區(qū)相似度高達80%[43]。早期,由于試驗方法和技術的限制,這些TK及其受體的研究一直是孤立的,無法建立聯(lián)系,甚至受體基因的克隆早于TK的發(fā)現(xiàn)。直到2000年之后,遺傳學家以果蠅為材料,利用轉基因技術構建了大量基因缺失或過表達品系[22],才開始基因功能研究,但有關TK的研究在果蠅中尚未開展廣泛研究,進展較為緩慢,其具體生理功能也尚不清楚。
配體/受體基因的獲得為研究TK/TKR信號轉導打開了一扇大門。目前,相關研究主要集中于基因組草圖已經完成的模式昆蟲。對于DTKR,Birse等利用不同體外細胞表達系統(tǒng),在人腎源細胞HEK293和果蠅S2細胞系中均成功表達了DTKR,隨后用人工合成的DTK1~DTK6等多肽分別處理以上兩種細胞,結果均能在短時間內提升細胞內Ca2+濃度及環(huán)磷酸腺苷(cAMP)水平,并且,這些信號隨多肽濃度的增大而顯著上升[44]。而對于果蠅NKD,研究者分別利用中國倉鼠Cricetulusbarabensis卵巢細胞CHO-PAM28和果蠅S2細胞系表達了該受體。同樣,分別加入人工合成的DTK1~DTK6、N端增加不同氨基酸殘基而仍保留C端基序不變的DTK6和其他昆蟲TK等神經肽刺激,結果除DTK6表現(xiàn)出較低的生物活性外,DTK1~DTK5等神經肽均無激活特性[45]。這些結果暗示了DTKR是DTK的潛在受體,而NKD則不是DTK的受體。在鱗翅目中也有類似研究,根據(jù)基因進化分析,發(fā)現(xiàn)家蠶TK受體BNGR-A24(Bombyxmorineuropeptide GPCR A24)與DTKR親緣關系更近;而BNGR-A32和BNGR-A33與NKD具有較近的親緣關系[46],這在一定程度上也表明TKR與NKD屬兩類不同的TK受體。此外,He等用2種不同細胞系即人胚胎腎細胞HEK293和草地貪夜蛾卵細胞系Sf21(Spodopterafrugiperdaovarian cell line 21)分別表達了BNGR-A24,隨后用人工合成的家蠶TK檢測生理活性,發(fā)現(xiàn)TK1~TK6(BmTK1~BmTK6)也分別能夠快速提升細胞內Ca2+濃度及cAMP水平,cAMP水平隨多肽濃度增加具有一定的濃度依賴性。這些結果也進一步表明了BNGR-A24是BmTK1~BmTK6潛在的受體[47]。
隨后一個重要試驗證明了果蠅、家蠶乃至其他一些模式昆蟲的各種受體與各自配體的生物學關系。Jiang等在果蠅、家蠶和赤擬谷盜等模式昆蟲中鑒定出一類C端相對保守、序列上類似TK的神經肽(C端為FxxxRa或YxxxRa motif),將其寓意為拉丁語 “出生”(natalisin,NTL)、并用拉丁化的NTL命名[48]。該團隊隨后進行了“受體/候選配體”體外細胞活性驗證,結果顯示果蠅DNTL1~DNTL5分別與受體NKD均有強烈的活性反應,相比而言,DTK6顯現(xiàn)出的活性反應遠低于DNTL1~DNTL5;而在家蠶中發(fā)現(xiàn)的12個NTL成熟多肽(BmNTL),其中BmNTL-1, BmNTL-3和 BmNTL-5(FxxxRa motif)可強烈激活BNGR-A33,而BmNTL-10和BmNTL-11(YxxxRa motif)強烈激活BNGR-A32;赤擬谷盜中發(fā)現(xiàn)的2個NTL對赤擬谷盜受體都具有生物活性反應[48]。自此,困擾了20多年有關TK與其受體的尋找終于“塵埃落定”,即DTKR(TkR99D)為DTK的受體,NKD(TkR86C)為果蠅NTL的受體;家蠶BNGR-A24為速激肽BmTK的受體,而BNGR-A32和BNGR-A33為BmNTL的受體;赤擬谷盜中也分別找到了TK和NTL及其相應受體。另外,在已知膜翅目昆蟲意大利蜜蜂和麗蠅蛹集金小蜂Nasoniavitripennis基因組中,并沒有發(fā)現(xiàn)NTL及其受體,而寄生于蜜蜂等膜翅目的常見螨蟲則存在相應的NTL及其受體,可以將螨蟲NTL及其受體作為靶標基因,這也為設計新的害蟲靶標基因提供了很好的思路[48]。
有關昆蟲TK/TKR信號轉導機制和功能研究才剛剛開始,在這些工作中,浙江大學周耐明教授、堪薩斯州立大學Park和哈佛大學Song等領銜或所在的研究團隊都做出了重要貢獻[10,17,47-48]。他們分別在家蠶、赤擬谷盜和黑腹果蠅等模式昆蟲中開展了相關研究,發(fā)現(xiàn)了一些重要的保守信號通路,即 TK與受體結合多數(shù)會引起cAMP、Ca2+等上升,可能通過PLC/Ca2+/PKC 或 AC/cAMP/PKA信號通路調控下游關鍵基因。同時,TK與受體結合后還可通過GRK5/PKC激酶途徑實現(xiàn)受體自身磷酸化,通過招募β-arrestin2/Kurtz蛋白復合體,經網格蛋白包被小窩(clathrin-coated pits)快速將受體內吞,隨后受體通過去磷酸化并最終恢復至細胞膜表面,形成完整信號轉導過程[10](圖3)。
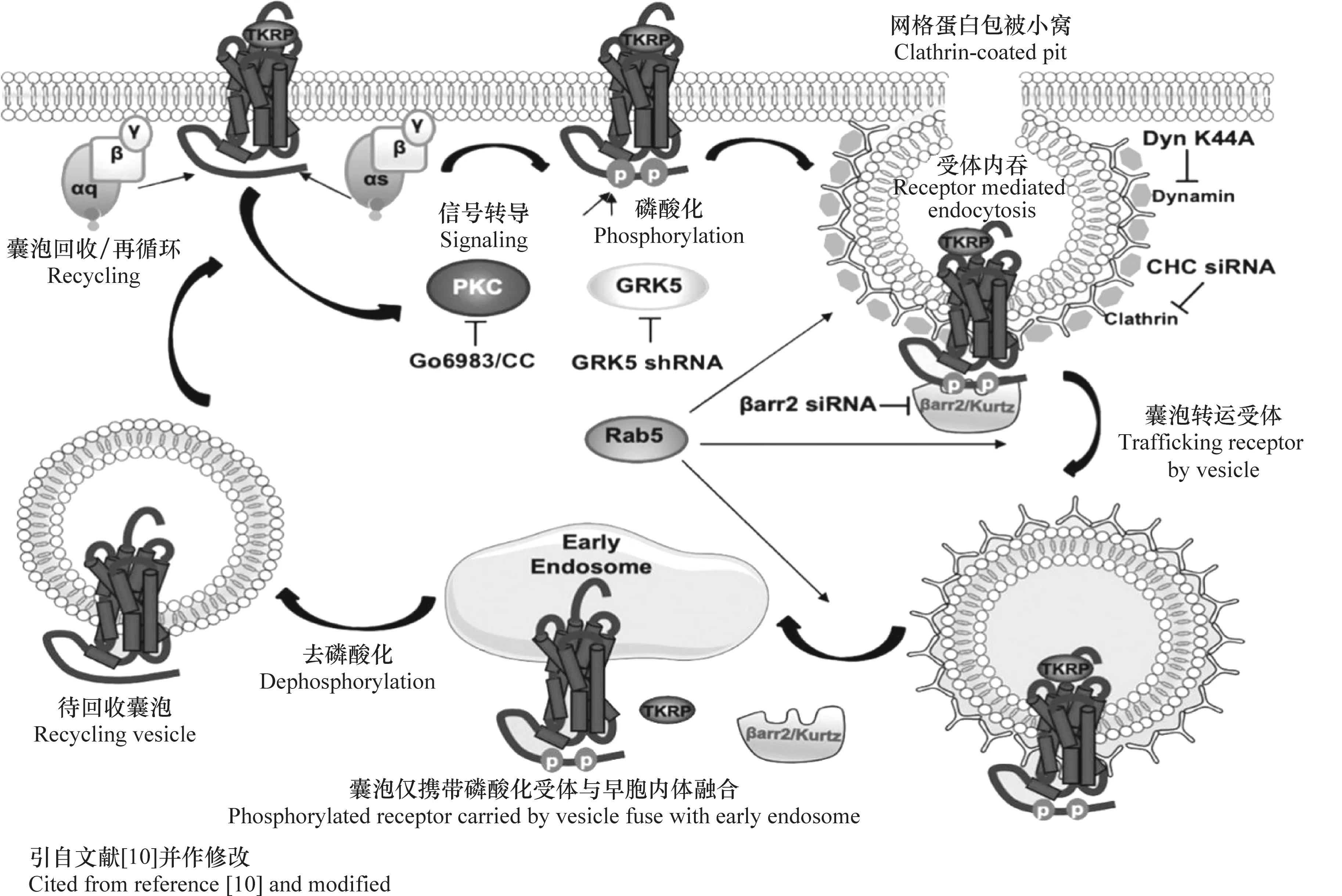
圖3 PKC,GRK5和β-arrestin2/BmKurtz參與TK/TKR信號通路中配體依賴的內吞過程
7 速激肽生理功能與行為調節(jié)研究
7.1 參與信息物質合成和感受
TK不僅具有促進成蟲對信息化合物的感受,而且在性信息素合成、調控兩性交配行為等方面具有重要功能。將轉基因果蠅腦神經和咽下神經節(jié)中TK轉錄水平調低后,與對照品系相比,其對丁醇(butanol)、醋酸異戊酯(isoamyl acetate)和苯甲醛(benzaldehyde)等揮發(fā)性化合物的趨向選擇性顯著降低[7]。此外,研究還發(fā)現(xiàn)TK也參與昆蟲對性信息素的感受。位于雄蟲前足的味覺神經元受體Gr68a,能夠通過味覺系統(tǒng)參與感知雄性同類射精管球(ejaculatory bulb)釋放的抑性欲信息素成分CH503[(3R,11Z,19Z)-3-acetoxy-11,19-octacosadien-1-ol],該性信息素成分是雄性交配過程中留給雌性的信息物質,可以降低交配過的雌性對其他同種雄性的吸引力,從而避免該雌蟲與其他雄蟲發(fā)生重復交配。這一重要功能的實現(xiàn),首先是通過配體即性信息素CH503與Gr68a受體結合后快速激活該信號通路,進一步通過肽能細胞(peptidergic cell)將該信號傳導至雄性中央腦(central brain),而咽下區(qū)域(subesophageal zone,SEZ)正是作為感受信息、參與抑制其他雄蟲與該雌蟲重復交配行為的黑匣子。該黑匣子接收到上游轉導信號后,其區(qū)域分布的8~10個能夠分泌TK的特異性神經元并快速響應,釋放TK成熟多肽,其中TK參與的一條信號可通過P1神經元簇(P1 neuron cluster),最終表現(xiàn)出抑制雄蟲與該雌蟲交配行為。利用RNAi將果蠅雄成蟲的SEZ區(qū)域內TK轉錄水平定點下調,這些特定區(qū)域TK下調后的雄蟲在與已交配過的雌蟲接觸后,其感知CH503能力顯著降低,不能有效避免重復交配[49]。將人工合成的飛蝗TK(Lom-TK-Ⅲ和 Lom-TK-Ⅵ)[19],注射到家蠶體內或利用含有TK成熟肽的培養(yǎng)液離體培養(yǎng)雌蛾的性信息素腺體,結果發(fā)現(xiàn),兩種方式均可促進家蠶雌成蟲性信息素的合成,而且隨著TK劑量的增加,雌性性信息素合成量具有顯著的濃度依賴性[50]。此外,研究還發(fā)現(xiàn)由TK祖先進化形成的另一分支神經肽natalisin,具有促進雙翅目黑腹果蠅、橘小實蠅Bactroceradorsalis和鞘翅目赤擬谷盜交配的功能[18,48,51]。但因物種差異性,鱗翅目蛾類與鞘翅目或雙翅目性信息素合成機制差異較大。目前研究已證明鞘翅目[51]和蜚蠊目性信息素合成由保幼激素(juvenile hormone,JH)調控[52-53],雙翅目由蛻皮激素(20-hydroxyecdysone,20E)調控[54],而鱗翅目蛾類主要由性信息素合成激活肽(pheromone biosynthetic activating neuropeptide,PBAN)調控[55]。隨著性信息素合成途徑研究的深入,我們總結發(fā)現(xiàn),自Fónagy首次報道TK參與性信息素合成之后[50], TK如何參與性信息素合成尚無新的報道,其作用機制也依然未知,而TK在參與家蠶等鱗翅目性信息素合成過程中扮演何種角色值得進一步探索。
7.2 參與取食及其他生理行為
昆蟲在取食過程中,不僅受JH和20E這兩大重要激素的影響,同時還受到諸多肽類激素的調節(jié)。研究表明,饑餓可誘導昆蟲體內TK激素水平快速上升[56]。而增加TK量能夠促進果蠅幼蟲后腸收縮蠕動[57],減少體內脂肪含量[17],這可能與其促進心側體內AKH的釋放、增加cAMP的水平具有一定的相關性[6]。同時,TK還可減少體重增加量,顯著縮短幼蟲饑餓后搜尋食物時間[47],這一現(xiàn)象與其他肽類激素如胰島素(insulin)和 sNPF(short neuropeptide F)類似[58]。進一步研究發(fā)現(xiàn),果蠅腦和中腸部分胰島素信號通路受到TK信號的嚴格調控,而且共同參與了維持體內海藻糖水平、抵御蟲體饑餓和氧化應激反應[56,59]。此外,TK還參與了痛覺敏感信號通路(hedgehog signaling),該環(huán)路中TK所在的神經元發(fā)生突變后可以提升對熱及紫外線的耐受性[60]。昆蟲在整個生命階段,時刻面臨著各種環(huán)境壓力,TK的分泌有助于昆蟲提升抗逆能力。自然界中,昆蟲取食各種寄主植物或其他食物的同時,從環(huán)境中獲得大量有益環(huán)境微生物[61],取食后的食物在腸道微生物的參與下易于蟲體吸收與利用, 更重要的是,某些有益微生物產生的短鏈脂肪酸乙酸鹽可有效激活蟲體先天免疫通路,進而通過促進TK大量分泌,在維持腸道脂肪穩(wěn)態(tài)乃至機體健康方面發(fā)揮著重要作用[16]。
8 展望
神經肽作為JH和20E之外的重要激素,在昆蟲生長發(fā)育及生理行為方面同樣發(fā)揮著重要的作用。研究發(fā)現(xiàn),TK與其他30多類神經肽一樣,均屬于典型的腦-腸多肽。總結目前國內外已有的神經肽功能研究文獻,發(fā)現(xiàn)其具有以下顯著特征,有關腦部神經肽的試驗側重于行為功能研究,有關中腸組織內神經肽的試驗則側重代謝或免疫研究。因此,基于神經肽組織特異性和表達的時序性,對于昆蟲TK的研究,已開始從傳統(tǒng)的分離鑒定及基因克隆研究逐步轉入基因功能與演化關系研究。一方面,可通過研究TK在不同類群昆蟲中的共性特征,如參與脂質穩(wěn)態(tài)[17]與饑餓下食物搜尋[56,59],了解昆蟲的取食與能量代謝機制。另一方面,發(fā)掘TK在不同類群中的特有功能,如具有聚集習性的切葉蟻Acromyrmexechinatior工蟻(worker caste)體內TK基因轉錄水平要顯著高于獨居型的蟻后(queen),可能與膜翅目特殊的社會性、或其蛋白序列中TK成熟肽數(shù)量較多具有一定的相關性[13,62]。此外,對昆蟲TK現(xiàn)有功能的研究和未知功能的挖掘,有助于探索昆蟲在取食寄主植物及逃避或適應天敵寄生過程中適合度權衡具有重要生態(tài)學意義,可進一步深入地闡釋TK在微生物-昆蟲-植物三級營養(yǎng)關系層面所扮演的重要角色[61,63]。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
