東革內酯通過InR-PI3K-mTOR-Bcl-2通路誘導斜紋夜蛾卵巢細胞凋亡
2023-02-03 10:07:12溫雪梅秦子昕黃建輝歐陽嘉敏邵雪花
植物保護 2023年1期
關鍵詞:檢測
溫雪梅, 秦子昕, 路 偉, 黃建輝,歐陽嘉敏, 萬 妍, 邵雪花
(1.廣東省農業科學院果樹研究所, 農業農村部南亞熱帶果樹生物學與遺傳資源利用重點實驗室, 廣東省熱帶亞熱帶果樹研究重點實驗室, 廣州 510640; 2.新疆農業大學農學院, 棉花教育部工程研究中心,自治區農林有害生物監測與安全防控重點實驗室, 烏魯木齊 830052; 3.廣東省中山市坦洲鎮農業服務中心, 中山 528400)
斜紋夜蛾Spodopteralitura(Fabricius)是一種世界性分布的農業害蟲,其幼蟲可為害十字花科、茄科、豆科以及花卉等300多種植物[1-2]。其繁殖力高、生命周期短和遷移能力強等特點增加了田間防治難度[3],化學防治為其主要防治手段,但隨著人們對農產品質量安全的重視,對綠色食品、有機食品的需求越來越多,這為生物農藥的發展帶來新的契機。因此,挖掘靶向斜紋夜蛾的高活性天然產物非常重要。
東革內酯(eurycomalactone,EC)來源于東革阿里Eurycomalongifolia(Jack)根部。該類化合物是一類具有苦味的降三萜類物質,不僅在醫學上具有抗瘧疾[4]、抗腫瘤、抗癌[5-7]、抗炎、抗病毒[8]等藥理作用,在農業上亦對多種害蟲具有拒食和毒殺等生物活性。從印度黃楝樹Samaderaindica種子和樹皮中分離的4個苦木苦味素類化合物(indaquassin C和samaderines A、B、C)對斜紋夜蛾幼蟲具有良好的拒食作用[9];從牛筋果Harrisoniaperforata(Blanco)枝葉中分離得到苦木苦味素類化合物perforalactone A和perforalactone B對蚜蟲具有較好的毒殺活性[10];從鴉膽子Bruceajavanica(Linn)中分離得到的苦木苦味素類化合物鴉膽因D對小菜蛾Plutellaxylostella的拒食活性為印楝素的6.2倍[11]。近期,作者從東革阿里中分離得到了東革內酯、寬纓酮(eurycomanone)、13,21-dihydroeurycomanone、11-dehydroklaineanone、15β-hydroxyklaineanone、eurycolactone F、6α-hydroxyeurycomalactone等7個苦木苦味素類化合物。研究表明,寬纓酮對草地貪夜蛾Spodopterafrugiperda、斜紋夜蛾、小菜蛾及甜菜夜蛾Spodopteraexigua等鱗翅目害蟲的拒食及抑制生長活性與印楝素相當或更高,同時兼具良好的植物內吸特性[12-18]。東革內酯具有抗癌、抗炎及抗病毒等藥理作用,臨床上常被作為有效的核因子κB(NF-κB)抑制劑使用[19],但其是否可抑制昆蟲細胞增殖及毒理機制目前尚未見報道。基于此,本文以斜紋夜蛾卵巢細胞SL-221為研究對象,采用CCK-8法檢測不同濃度東革內酯對SL-221細胞的毒力,并通過流式細胞術測定該化合物對SL-221細胞周期、線粒體膜電位及凋亡的影響;最后,利用實時熒光定量PCR(qPCR)技術探究東革內酯處理SL-221細胞后相關基因的表達情況,旨在探究東革內酯誘導SL-221細胞發生凋亡的分子機制,為斜紋夜蛾的綠色防控和東革內酯的開發利用提供新的視角。
1 材料與方法
1.1 供試材料
1.1.1供試細胞
斜紋夜蛾卵巢細胞SL-221,由廣東省農業科學院果樹研究所資源與環境實驗室提供,用SIM SF昆蟲細胞培養基(含5%胎牛血清),在27℃恒溫箱中培養。
1.1.2供試藥劑
東革內酯(eurycomalactone,EC),分子量348,CAS號:23062-24-0,由廣東省農業科學院果樹研究所資源與環境實驗室提供,結構式見圖1。

圖1 東革內酯的結構式
1.1.3供試試劑和儀器
供試試劑及試劑盒:胎牛血清購自蒙馬生物科技有限公司;SIM SF Expression Medium(MSF1)、CCK-8(M128)、細胞周期與細胞凋亡檢測試劑盒(C1052)、Annexin Ⅴ-FITC/PI凋亡檢測試劑盒(CA1020)、線粒體膜電位檢測試劑盒(M8650)和二甲基亞砜(dimethyl sulfoxide,DMSO)均購自北京索萊寶科技有限公司;RNA提取試劑盒、呲溜極速反轉錄試劑盒和2×M5 HiPer Realtime PCR Super mix with Low Rox(SYBR green with anti-Taq)(MF797-05)均購自北京聚合美生物科技有限公司;PBS緩沖液購自博奧森生物技術有限公司;印楝素,純度≥98%,由華南農業大學亞熱帶農業生物資源保護與利用國家重點實驗室提供。
供試儀器:恒溫培養箱(DH-360)購自北京科偉永興儀器有限公司;多功能酶標儀(Spark)購自上海帝肯實驗器材有限公司;流式細胞儀(FACS Verse)購自上海土森視覺科技有限公司;熒光定量PCR儀(Quant Studio 3)購自賽默飛世爾科技有限公司。
1.2 試驗方法
1.2.1CCK-8法檢測東革內酯對SL-221細胞的毒力
將凍存的SL-221細胞置于37℃水浴迅速攪拌融化,在超凈工作臺內將細胞液轉入2 mL離心管,800 r/min離心5 min,棄上清,用2 mL含10%胎牛血清的SIM SF昆蟲細胞培養基重懸細胞,將細胞懸液轉于細胞培養瓶,27℃恒溫箱中培養。用DMSO將東革內酯配制成10 mg/mL的母液,并用無血清SIM SF培養基逐級稀釋至100、50、25、12.5、6.25、3.125 μg/mL和1.562 5 μg/mL。取對數生長期的SL-221細胞,用含5%胎牛血清的SIM SF培養基將細胞濃度調節至5×105cfu/mL,接種于96孔板,每孔100 μL,置于27℃恒溫箱過夜。試驗前棄去96孔板中原有的含血清培養基,分別加入各濃度含藥培養基,每濃度4板重復,以含0.1% DMSO的無血清SIM SF培養基作為空白對照,以相同濃度印楝素處理作為陽性對照,置于27℃恒溫箱孵育48 h。48 h后向每孔中加入10 μL CCK-8溶液,繼續避光孵育2~3 h。最后用酶標儀測定其在450 nm處的吸光度,根據公式計算細胞抑制率,并利用SPSS軟件計算其抑制中濃度(IC50)。
細胞增殖抑制率=(A零加藥-A加藥)/(A零加藥-A空白)×100%,式中,A加藥為含有SL-221細胞、含藥培養基和CCK-8溶液孔的吸光度;A空白為含有含藥培養基和CCK-8溶液孔的吸光度;A零加藥為含SL-221細胞、CCK-8溶液但無藥劑孔的吸光度。
1.2.2東革內酯對SL-221細胞形態的影響
用DMSO將東革內酯配制成10 mg/mL母液,在無血清SIM SF昆蟲細胞培養基中添加東革內酯使其終濃度為4 μg/mL。用含5%胎牛血清的SIM SF培養基將SL-221細胞培養至對數生長期(5×105cfu/mL),取2 mL接種于細胞培養皿(d=6 cm)中,置于27℃恒溫箱培養。待細胞密度達到85%~90%時,棄去原有培養基,更換為含4 μg/mL東革內酯的SIM SF培養基,培養0、3、6、12、24 h和48 h后在倒置顯微鏡下觀察,其中0 h為對照。拍照記錄細胞形態變化。
1.2.3流式細胞術檢測東革內酯對SL-221細胞周期的影響
用DMSO將東革內酯配制成10 mg/mL母液,進而配制含1 μg/mL東革內酯的無血清SIM SF培養基。取對數生長期的SL-221細胞(5×105cfu/mL)接種至12孔板,每孔2 mL,于27℃恒溫箱培養。待細胞密度達到85%~90%時,棄去原有培養基,加入含1 μg/mL東革內酯的SIM SF培養基,培養0、3、6、12、24 h和48 h,每個時間點設置3個重復孔, 試驗重復3次,于27℃恒溫箱中分別孵育0、3、6、12、24 h和48 h,其中0 h為對照。之后收集細胞,800 r/min離心5 min,棄上清。用PBS洗滌細胞,然后加入預冷的70%乙醇置于4℃冰箱過夜。次日用PBS洗滌細胞2次,加入100 μL RNaseA溶液重懸細胞,37℃水浴30 min;之后加入400 μL PI染色液,4℃避光孵育30 min。完成后立即用流式細胞儀在激發波長488 nm處檢測紅色熒光,用FlowJo軟件統計分析處于G0/G1(DNA合成前期)、S(DNA合成期)和G2/M(DNA合成后期)的細胞數量。
1.2.4JC-1熒光標記法檢測東革內酯對SL-221細胞線粒體膜電位的影響
細胞處理方法和重復情況同1.2.3。收集1 μg/mL東革內酯分別處理0、3、6、12、24 h和48 h后的細胞,用PBS清洗細胞2次。之后加入0.5 mL JC-1染色工作液,37℃避光孵育20 min;孵育結束后,600 r/min離心4 min,棄上清,用JC-1染色緩沖液(1×)清洗細胞2次;再加入1 mL JC-1染色緩沖液(1×)重懸細胞。最后置于流式細胞儀檢測激發波長585 nm,發射波長590 nm處的PE(紅色)熒光;激發波長515 nm,發射波長529 nm處的FITC(綠色)熒光,并計算紅綠熒光的相對比例來衡量線粒體膜電位變化情況(紅綠熒光的相對比例=紅色熒光百分比/綠色熒光百分比)。
1.2.5Annexin Ⅴ-FITC/PI雙染檢測東革內酯對SL-221細胞凋亡的影響
Annexin Ⅴ-FITC/PI凋亡檢測試劑盒是一種采用Annexin Ⅴ-FITC與PI雙染法進行細胞早期凋亡分析的檢測試劑盒。正常細胞具有完好的細胞膜,此時細胞膜中的磷脂酰絲氨酸(phosphotidylserine,PS)位于胞膜內側。細胞早期凋亡時,細胞膜結構發生改變,使胞膜內側的PS外翻到胞膜表面。根據Annexin Ⅴ易于結合PS并對其高度親和的特性可檢測凋亡的細胞。熒光染料碘化丙啶(propidiumiodide,PI)不能通過正常細胞,但能對凋亡晚期的細胞進行染色,因此將Annexin Ⅴ與PI配合染色,以區別凋亡早期細胞和凋亡晚期細胞。細胞處理方法和重復情況同1.2.3。收集1 μg/mL東革內酯分別處理0、3、6、12、24 h和48 h后的細胞,用預冷的PBS清洗細胞2次,800 r/min離心5 min,棄上清;加入250 μL Binding Buffer重懸細胞;取100 μL細胞懸液放入流式管中,按照試劑盒說明書加入5 μL Annexin Ⅴ-FITC和10 μL PI溶液,輕搖混勻,避光孵育15 min。置于流式細胞儀檢測激發波長535 nm,發射波長615 nm處的PI(紅色)熒光;激發波長488 nm,發射波長525 nm處的FITC(綠色)熒光,并用FlowJo軟件分析細胞凋亡情況。
1.2.6RT-qPCR法檢測東革內酯對SL-221細胞相關基因表達情況的影響
細胞處理方法同1.2.3,以SL-Actin作為內參基因,引物序列見表1,由生工生物工程(上海)股份有限公司合成。收集1、2 μg/mL和4 μg/mL東革內酯分別處理0、3、6、12、24 h和48 h后的SL-221細胞,其中0 h為對照,利用RNA提取試劑盒提取其總RNA,再通過反轉錄試劑盒合成cDNA。反應體系(20 μL):5×Sprint gDNA Remover Mix 2 μL、RNA模板1 μL、DEPC-ddH2O 7 μL,將反應液混勻,42℃溫育2 min,冰上冷卻;再加入5×M5 Sprint RT Mix 4 μL、DEPC-ddH2O 6 μL,50℃ 5 min、85℃ 5 s。利用SYBR green with anti-Taq試劑盒進行PCR擴增,反應體系(10 μL):cDNA 1 μL、2×M5 HiPer Realtime PCR Super mix with Low Rox 5 μL、Primer 0.5 μL,RNase Free ddH2O 3.5 μL。反應程序為:95℃預變性 30 s;95℃ 5 s,60℃ 30 s,40個循環;95℃ 15 s,60℃ 1 min,95℃ 15 s。每樣品重復3次,通過熒光定量PCR得到目的基因與內參基因的Ct值,利用2-ΔΔCt法(△Ct=Ct目的基因-Ct內參基因)計算不同處理時間下SL-221細胞相關基因的相對表達量。

表1 斜紋夜蛾生長發育相關基因 RT-qPCR 引物序列
1.3 數據分析
本試驗結果均為3次以上重復試驗所得,運用IBM SPSS Statistics 19軟件分析數據,各處理組所得平均值之間的差異通過單因素方差分析及Duncan氏多重比較進行檢驗評估,P<0.05表示在統計學上具有顯著性差異,使用ImageJ軟件處理細胞圖片,FlowJo軟件分析細胞流式數據,Origin 2017進行繪圖。
2 結果與分析
2.1 CCK-8法檢測東革內酯對SL-221細胞的毒力
檢測結果(表2)顯示,東革內酯對SL-221細胞48 h的IC50為1.98 μg/mL,而陽性對照印楝素48 h的IC50為7.51 μg/mL,東革內酯對SL-221細胞有明顯的抑制細胞增殖的活性。濃度為1.562 5、3.125、6.25、12.5、25、50 μg/mL和100 μg/mL的東革內酯處理48 h的細胞增殖抑制率分別為43.87%、58.96%、66.52%、74.42%、79.41%、81.51%和86.97%(圖2)。東革內酯對SL-221細胞的抑制增殖活性與濃度呈正相關。

表2 東革內酯對SL-221細胞的毒力
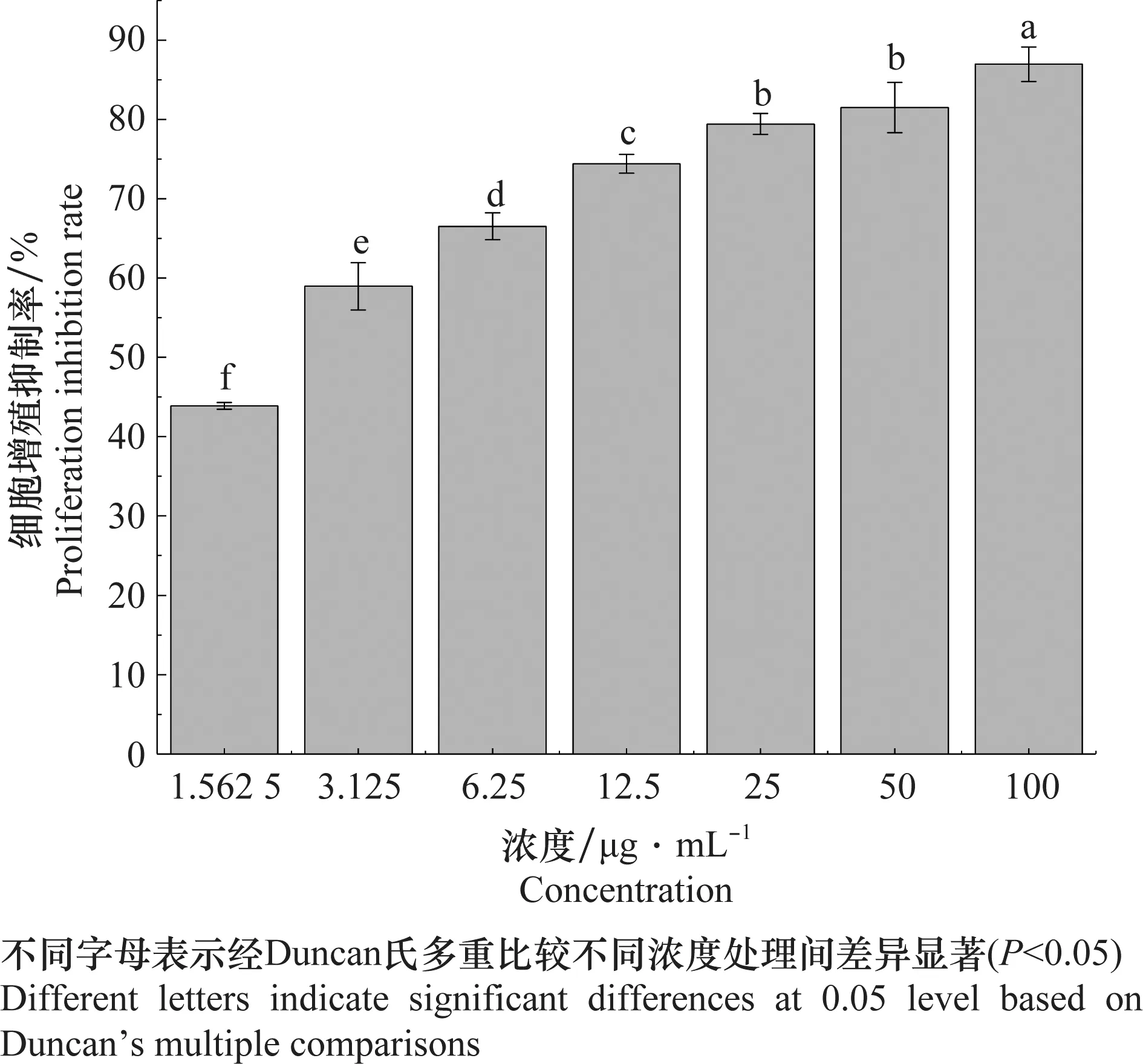
圖2 東革內酯處理SL-221細胞48 h的細胞增殖抑制率
2.2 東革內酯處理后SL-221細胞形態變化
經4 μg/mL東革內酯處理后SL-221細胞形態出現了明顯變化。從圖3可以看出,0 h(對照)時,SL-221細胞形態飽滿光滑,呈圓形或橢圓形;處理 3 h 和6 h后,細胞膜光滑度下降,部分細胞膨脹;12 h后,細胞膨脹明顯,細胞膜表面粗糙;處理24 h后,細胞膜粗糙,細胞腫脹破裂,并可見凋亡小體(圖3e);48 h后,細胞內容物溢出,凋亡小體數量明顯增多,大部分細胞不能維持貼壁(細胞培養瓶底)生長。綜上,隨著藥劑處理時間延長,細胞凋亡現象明顯,細胞活力下降,東革內酯對SL-221細胞增殖的抑制活性呈現時間依賴性。
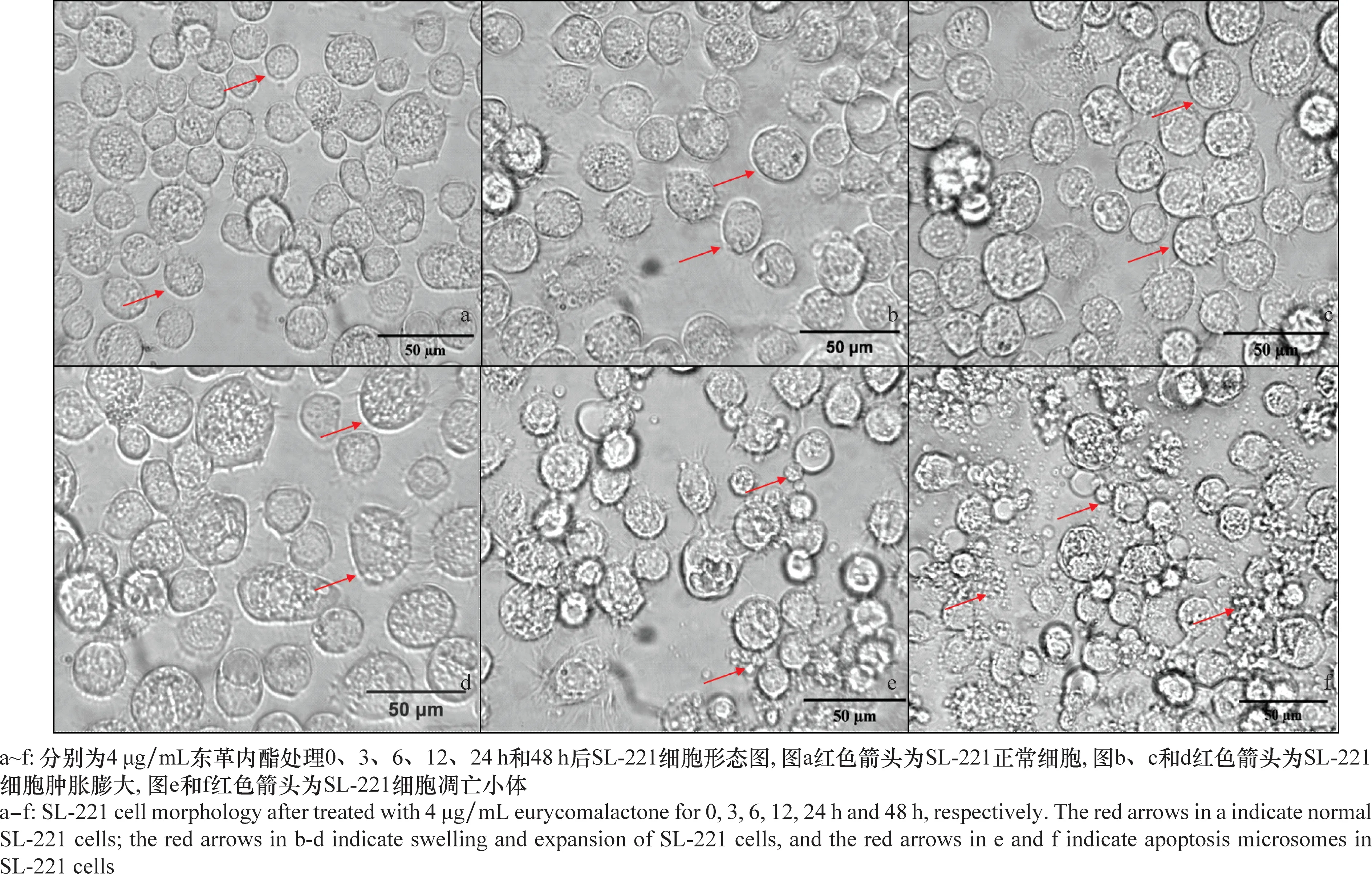
圖3 4 μg/mL東革內酯處理后SL-221細胞形態圖
2.3 流式細胞術檢測東革內酯對SL-221細胞周期的影響
經1 μg/mL東革內酯處理0、3、6、12、24 h和48 h的SL-221細胞中處于不同周期的細胞百分率的分布情況見圖4 a,其統計結果見圖4 b。從結果可以看出,與0 h(對照)相比,12、24 h和48 h的G0/G1期細胞百分率均顯著下降P<0.05),而G2/M期細胞百分率均顯著上升(P<0.05)。其中,處理48 h的G0/G1期細胞所占百分率從(35.80±3.58)%下降至(20.67±2.10)%,S期細胞所占百分率由(33.47±8.34)%下降至(16.90±3.03)%,而G2/M期細胞所占百分率從(30.47±5.43)%增加至(62.67±0.67)%。由此可見,東革內酯可將SL-221細胞周期阻滯于G2/M期。

圖4 東革內酯對SL-221細胞周期的影響
2.4 JC-1熒光標記法檢測東革內酯對SL-221細胞線粒體膜電位的影響
經1 μg/mL東革內酯處理不同時間的SL-221細胞線粒體膜電位的分布情況見圖5a。在線粒體膜電位較高時,JC-1聚集形成聚合物,可以產生紅色熒光;在線粒體膜電位較低時,JC-1為單體,可以產生綠色熒光。SL-221細胞線粒體膜電位變化情況的統計結果見圖5 b,隨著處理時間的增加,SL-221細胞線粒體紅/綠熒光比例顯著下降(P<0.05)。0 h(對照)SL-221細胞線粒體紅/綠熒光比例為11.55,處理3、6、12、24 h和48 h時,SL-221細胞線粒體紅/綠熒光比例分別為3.51、1.40、0.33、0.29和0.19,與0 h(對照)相比下降了69.61%、87.88%、97.14%、97.49%和98.35%。可見,東革內酯可顯著降低SL-221細胞的線粒體膜電位,且呈現時間依賴性。
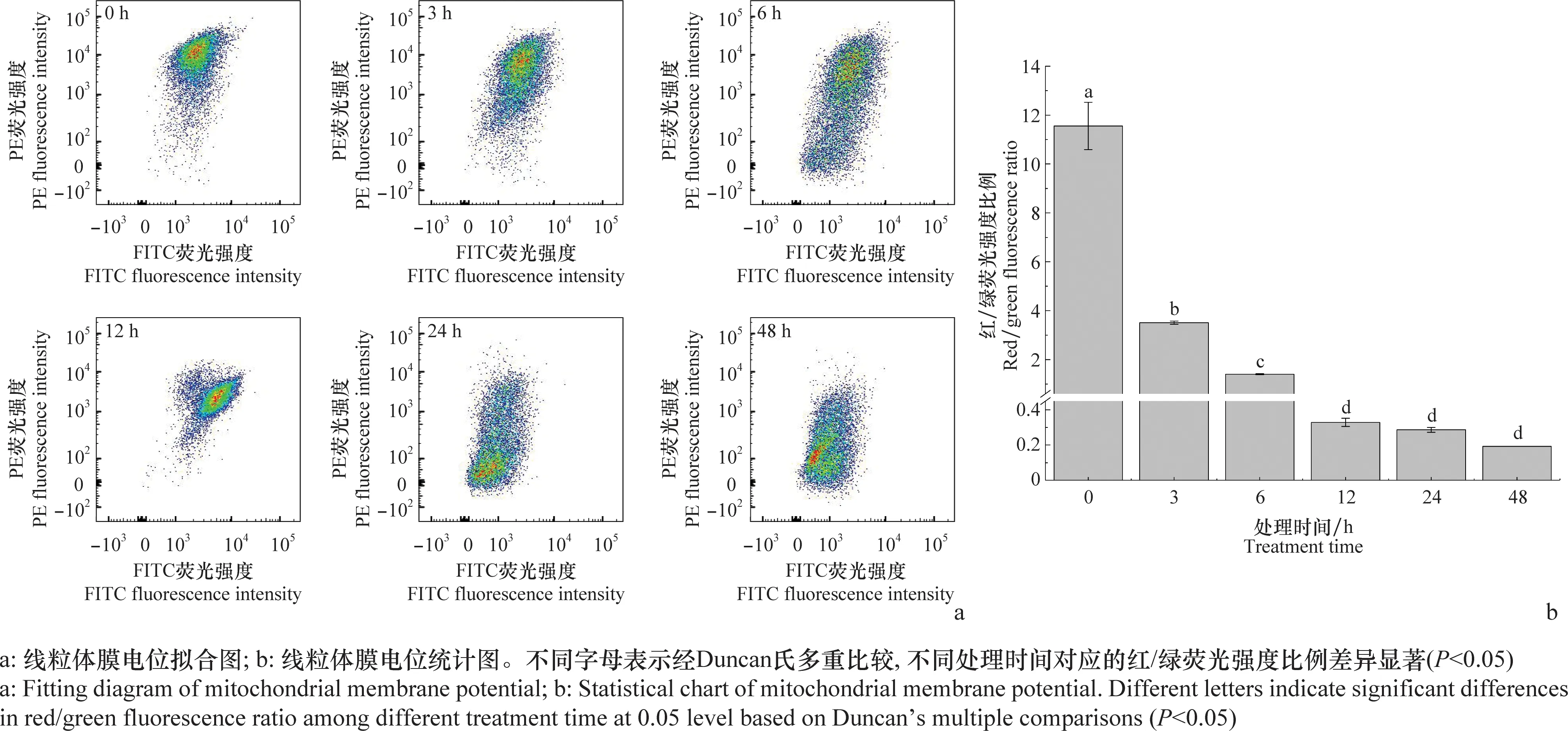
圖5 東革內酯對SL-221細胞線粒體膜電位的影響
2.5 Annexin Ⅴ-FITC/PI雙染檢測東革內酯對SL-221細胞凋亡的影響
Annexin Ⅴ-FITC/PI雙染檢測1 μg/mL東革內酯處理不同時間下,不同類型的SL-221細胞的分布情況見圖6 a。其中Q1象限為壞死細胞,Q2和Q3象限分別為晚期凋亡細胞和早期凋亡細胞,Q4為正常細胞。Annexin Ⅴ-FITC/PI雙染統計結果見圖6 b。結果顯示,東革內酯處理不同時間下,SL-221正常細胞百分率與0 h(對照)相比顯著下降(P<0.05),而早期凋亡細胞百分率顯著上升(P<0.05)。其中,處理48 h與0 h相比,SL-221正常細胞百分率由75.47%降低至32.80%,早期凋亡細胞百分率由8.42%增加到32.03%,晚期凋亡細胞百分率由11.43%增加到33.13%。由此可見,東革內酯可誘導SL-221細胞凋亡,且隨著時間的延長凋亡加劇。

圖6 東革內酯對SL-221細胞凋亡的影響
2.6 RT-qPCR法檢測東革內酯對SL-221細胞凋亡相關基因表達情況的影響
1、2、4 μg/mL東革內酯處理SL-221細胞后,SL-p53、SL-CytochromeC和SL-InR基因表達均呈現上調趨勢(圖7),處理48 h,SL-p53基因表達與對照相比分別上調17.53、67.92倍和90.10倍;SL-CytochromeC基因表達與對照相比分別上調38.03、65.15倍和74.15倍;SL-InR基因表達與對照相比分別上調2.34、4.19倍和4.34倍。SL-Bcl-2、SL-mTOR和SL-PI3K基因的表達量隨處理時間的延長均呈現下降趨勢。其中,1、2、4 μg/mL東革內酯分別處理48 h后,與對照相比,SL-Bcl-2基因表達分別下調94.76%、96.49%和96.94%;SL-mTOR基因表達與對照相比分別下調96.10%、97.30%和98.14%;SL-PI3K基因表達與對照相比分別下調86.55%、87.01%和93.05%。
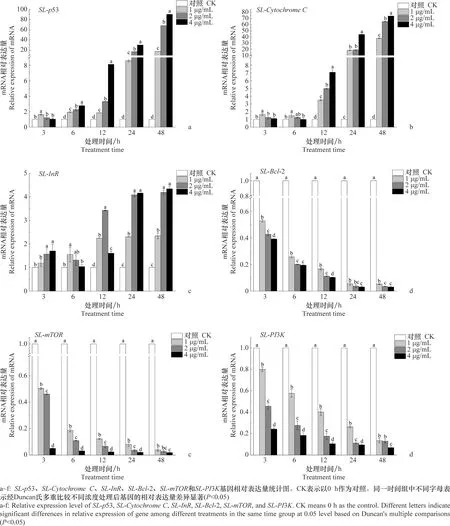
圖7 東革內酯對SL-221細胞凋亡相關基因表達情況的影響
綜上,1、2、4 μg/mL東革內酯可誘導凋亡標志基因SL-p53、SL-InR和SL-CytochromeC顯著上調表達(P<0.05),同時抑制凋亡抑制因子SL-Bcl-2的表達。細胞凋亡信號通路上游的SL-mTOR和SL-PI3K基因表達均受到顯著抑制(P<0.05)。表明東革內酯可通過InR-PI3K-mTOR-Bcl-2信號通路誘導SL-221細胞凋亡。
3 結論與討論
斜紋夜蛾屬于重大農業害蟲,目前主要依靠化學農藥進行防控,但監測表明其已對多種化學殺蟲劑產生了抗性[20-21]。因此,尋找高效低毒的藥物及靶點是農藥領域研究的熱點和難點。從天然產物中挖掘新活性物質,研發替代防治產品至關重要。本文比較了印楝素與東革內酯對SL-221細胞增殖的抑制作用,發現東革內酯能以更小的劑量(IC50為1.98 μg/mL)抑制SL-221細胞增殖,隨著東革內酯濃度增高,SL-221細胞數目減少,細胞膨脹、核固縮(細胞核染色質DNA濃聚、皺縮,使核體積減小)破碎,細胞碎片增多,提示其可能具有開發為防控斜紋夜蛾的藥物的潛力。東革內酯能否通過誘導凋亡來抑制SL-221細胞增殖,目前仍不清楚。我們通過流式細胞術發現,東革內酯能使SL-221的細胞周期停滯于G2/M期,降低線粒體膜電位,且能顯著誘導細胞凋亡。
細胞凋亡(apoptosis)是一種特殊的細胞死亡方式,又被稱為程序性細胞死亡(programmed cell death, PCD),指為維持內環境穩定,由基因控制的細胞自主有序的主動死亡過程[22]。在形態方面,細胞凋亡表現為:細胞質濃縮、染色質聚集、核酸內切酶活化和出現凋亡小體等特征。發生過程包括:凋亡的起始、凋亡小體形成和凋亡小體被鄰近細胞吞噬并消化[23]。通常情況下,細胞凋亡是機體為了維持各器官的穩定性而進行的一種自我調控,但當細胞凋亡調控失衡時,可引起細胞過度增殖或過度凋亡,導致相關疾病的發生[24]。細胞凋亡主要有內源性和外源性兩種途徑。內源性途徑主要包括細胞應激、DNA損傷、發育信號、存活因子缺失等[25]。這個途徑是由Bcl-2家族介導,該家族存在兩種類型的蛋白,促凋亡蛋白(Bax、Bad等)和抑凋亡蛋白(Bcl-2)[26]。在內源性途徑中,CytochromeC的表達對細胞凋亡起著關鍵作用[27]。CytochromeC從線粒體釋放到胞質中,形成凋亡復合體,進而激活核酸內切酶切割DNA,促使細胞凋亡[28]。p53是一種腫瘤抑制因子,它可以對細胞應激或DNA損傷作出響應,被激活后的p53可誘導細胞的周期停滯、凋亡和衰老[29-30]。當蛋白表達異常時可能促使細胞進入不可逆的凋亡階段。我們發現東革內酯可抑制凋亡因子SL-Bcl-2的表達,促進SL-p53、SL-InR和SL-CytochromeC基因的表達。InR-PI3K/mTOR是一條經典的胞內信號通路,主要調節細胞的增殖、自噬和凋亡等,其異常活化或被抑制均會誘導下游信號途徑的紊亂。PI3K是該信號通路的重要組成因子,可在多種生長因子的刺激下被激活,參與細胞自噬的調節,降低細胞自噬力[31-32]。mTOR位于PI3K/mTOR信號通路的下游,屬于磷酸肌醇激酶家族成員,PI3K激活后,其下游的mTOR也被激活,mTOR激活可抑制細胞自噬[33]。本研究表明SL-mTOR和SL-PI3K基因表達均顯著下調(P<0.05),進一步證實了東革內酯可誘導SL-221細胞凋亡,這與印楝素可誘導SL-1細胞凋亡的結果一致[34]。
近年來研究表明,活性成分可通過上調InR,負反饋抑制PI3K/Akt通路來誘導細胞凋亡[35]。本試驗中,我們發現東革內酯可顯著上調InR基因的表達,同時抑制PI3K和mTOR基因的表達,因此,我們推測東革內酯誘導SL-221細胞發生的凋亡可能與抑制PI3K和mTOR的磷酸化、抑制Bcl-2的表達,促使細胞凋亡有關。
綜上所述,東革內酯可通過InR-PI3K-mTOR-Bcl-2信號通路誘導SL-221細胞發生凋亡,后續將進一步研究東革內酯是否還通過誘導其他死亡方式發揮抑制昆蟲細胞增殖及其具體機制,為東革內酯的開發和利用提供新的依據和思路。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
