行間生草種類對蘋果園春季土壤蒸發、空氣濕度和土壤貯水的影響
2023-02-07 02:22:00秦文利
草業學報 2023年1期
秦文利
(河北省農林科學院農業資源環境研究所,河北省肥料技術創新中心,河北 石家莊 050051)
果園生草是一種先進的果園土壤管理制度,能夠抑制土壤蒸發、增加空氣濕度、改善微域小氣候,對防治我國北方果樹因春季降水稀少、升溫快、空氣干燥引起的落花落果及裂紋病有重要作用[1-5],但牧草生長會與果樹競爭土壤水分[6-8],加劇果園用水矛盾。華北平原是我國蘋果(Malus pumila)的主產區,水資源短缺嚴重制約著該區蘋果產業的可持續發展。因此,探明蘋果園生草中土壤蒸發、空氣濕度、土壤水分等變化特征及相互關系與影響因素對高效利用果園土壤水分、穩定蘋果產量與質量、推廣果園生草具有重要意義。
土壤蒸發與生產過程無關,屬無效耗水,降低土壤蒸發是提高農田水分利用效率的一個重要途徑[9-10]。地表覆蓋物物理阻隔了土壤-大氣之間的能量傳輸和水分轉換,可增加土壤水分蒸發阻力[11-13]。研究表明,土壤蒸發阻力與秸稈覆蓋量、植被葉面積指數(leaf area index,LAI)呈正相關關系[14-16],與土壤含水量、地溫呈負相關關系[17-19]。果園生草后牧草阻隔太陽直射地表,生草區光合有效輻射(photosynthetically active radiation,PAR)透射率下降不利于地溫上升、耗水增加降低土壤含水量,二者皆抑制了土壤蒸發[4,20]。但生草種類不同PAR透射率、LAI、土壤含水量及地溫的變化不同,進而土壤蒸發的變化特征不同,目前關于這方面的研究甚少。
牧草群體蒸散的水分還可調控果園空氣濕度[4]。牧草種類及其生育進程對不同季節果園空氣濕度影響不同。李春波[21]的研究表明,果園生草能提高6至9月葡萄(Vitis vinifera)園空氣濕度。高方勝等[22]的研究發現,與白三葉(Trifolium repens)相比,魯西北蘋果園種植黑麥草(Lolium perenne)能提高5月10日左右的蘋果樹冠層空氣相對濕度,這與其返青早、生長快相關。目前,果園生草對果樹冠層空氣濕度影響的研究多集中于多年生牧草,而隨著果園觀光旅游業、飼草業的發展及果品生產化肥零增長的需求,景觀作物二月蘭(Orychophragmus violaceus)、優質牧草小黑麥(×TriticosecaleWittmack)和綠肥作物毛葉苕子(Vicia villosa)等越年生牧草越來越受到果樹種植戶的喜愛,但關于越年生牧草對果樹冠層空氣濕度影響的研究較少。牧草群體蒸散的水分來自于根系對土壤水分的吸收,而關于果園生草對土壤水分影響的各個研究結論截然不同,這可能與生草種類、年限及季節不同有關。惠竹梅等[8]的研究認為,行間生草降低了葡萄園土壤含水量,不同種類牧草對不同季節土壤含水量的降幅影響不同。籍增順等[23]的研究表明,旱地蘋果園生草4年后能提高0~60 cm土層土壤含水量。李同川等[24]的研究發現,果園生草對土壤含水量的影響與牧草生長進程有關。牧草萌動以前,因覆蓋地表抑制蒸發可提高土壤含水量;萌動之后,隨生長進程加快土壤水分消耗增加,降低了土壤含水量。種植小冠花(Coronilla varia)、百脈根(Lotus corniculatus)、雞腳草(Dactylis glomerata)等會造成4月初至5月初蘋果樹行間土壤水分虧缺。果園生草雖能改善北方春季果樹冠層空氣濕度,但生草種類不同對果樹冠層空氣濕度和土壤水分消耗的影響不同,目前關于果園生草后果樹冠層空氣濕度變化與土壤水分消耗的關系及不同種類牧草對土壤蒸發的抑制能否彌補對土壤水分消耗的研究還鮮有報道。
本研究在蘋果樹行間種植越年生牧草(小黑麥、二月蘭、毛葉苕子)和多年生牧草[鴨茅(Dactylis glomerata)、白三葉],探討行間生草種類對華北平原蘋果園4-5月土壤蒸發、空氣相對濕度、土壤水分等環境因子的影響特征,旨在揭示果園生草后土壤蒸發、空氣相對濕度、土壤水分的變化規律及其相互關系,為華北蘋果主產區采取合理的果園生草模式、實現水分高效利用、促進蘋果產業的可持續發展提供理論依據和技術支撐。
1 材料與方法
1.1 試驗園區概況
試驗于2019-2021年在河北省農林科學院武羅綜合示范園區蘋果園(北緯37°49′8.94″-37°49′57.62″,東經115°56′7.36″-115°56′37.97″,海拔19.0 m)進行。該區屬于冀中南平原,暖溫帶半干旱季風氣候,年平均氣溫12.6℃,年平均降水量518.0 mm,年平均日照時間2575.6 h,無霜期195 d左右。土壤為壤質潮土,生草前0~60 cm土層土壤pH值為7.85,有機質含量為9.07 g·kg-1,全氮含量為0.55 g·kg-1,全磷含量為0.80 g·kg-1,堿解氮含量為32.14 mg·kg-1,有效磷含量為5.17 mg·kg-1,速效鉀含量為111.83 mg·kg-1。0~20 cm、20~40 cm、40~60 cm、60~80 cm、80~100 cm的土層土壤容重分別為:1.40、1.38、1.30、1.32、1.37 g·cm-3。蘋果樹品種為天紅2號,樹齡4年生,株行距2 m×4 m,2021年覆蓋度為45%。果樹采用小管出流滴灌技術進行水肥一體化管理,每年在果樹萌芽期(3月中下旬)、落花期(4月下旬)、果實膨大期(6月下旬)3個果樹關鍵生育期每隔10 d灌水1次,2019、2020、2021年總灌水量分別為1200、1200、1350 m3·hm-2,每年隨灌水施用金正大大量元素水溶肥[總氮(N)、五氧化二磷(P2O5)、氧化鉀(K2O)含量均為20%,肥料中含有微量元素]750 kg·hm-2。
1.2 試驗設計
試驗開始于2019年9月中旬,共設6個處理,分別為二月蘭(Ory.)、小黑麥(Trit.)、毛葉苕子(Vic.)、白三葉(Trif.)、鴨茅(Dac.)5個生草區和1個清耕對照區(CK),其中二月蘭、小黑麥、毛葉苕子為越年生牧草,白三葉、鴨茅為多年生牧草。二月蘭、小黑麥、毛葉苕子均采用機械播種,播種量分別為22.5、120.0、90.0 kg·hm-2,播深分別為1.5、3.0、4.0 cm,行距分別為30、20、30 cm,播種后于翌年5月初機械切碎地上部分覆蓋還田。2020年二月蘭、毛葉苕子、小黑麥播種時間分別為9月18日、10月8日、10月12日,還田時間均為2021年5月6日;白三葉、鴨茅均采用人工播種,播種量分別為30.0、45.0 kg·hm-2,播深均為1.5 cm,行距均為20.0 cm,于2019年9月15-16日播種。2019年播種前,所有試驗地塊隨整地施入有機肥105×103kg·hm-2、復合肥(N∶P2O5∶K2O=15∶15∶15)150 kg·hm-2做基肥并灌溉造墑,灌水量為800 m3·hm-2,播種完后牧草不再追肥和灌溉。2020年秋季播種越年生牧草時不再施基肥,但整地時灌溉造墑,灌水量為800 m3·hm-2,同時白三葉、鴨茅多年生牧草和清耕對照處理灌溉等量的水。生草帶寬為2.2 m,樹行每側保留0.9 m清耕保護帶。每個處理地塊面積為150 m×160 m=24000 m2,每個處理重復3次,隨機布設。果樹田間管理一致。
1.3 測定項目與方法
1.3.1 土壤溫度 在2021年4月15日13:30-14:30時段內采用精確度為0.1℃的直角地溫計測定蘋果樹行間中線上5、10、15、20、25 cm不同深度土層土壤溫度。每處理生草區隨機選取4個采樣點,每個采樣點每個土層測定2次重復,取平均值計算。
1.3.2 牧 草 群 體LAI及PAR透射率 在2021年4月15日11:00-14:00時段內采 用SunScan冠 層分析系統(英國)測定不同處理生草區牧草群體LAI和冠層PAR傳輸特性。測定時,保持漫射系數傳感器(beam fraction sensor,BFS)水平且高度超過牧草冠層(避開果樹遮蔭),將SunScan探測桿置于冠層底部并離地面5 cm,垂直向上測量透射到底部的PAR(transmission of photosynthetically active radiation,PART),BFS將同步測量冠層頂部入射的PAR(incidence of photosynthetically active radiation,PARI),SunScan直接測出LAI。每處理生草區隨機選取4個采樣點,每個采樣點測定2次重復,取平均值計算。PAR透射率(Tr)=PART/PARI。因測定時白三葉未完全返青、植株低矮,故未測定該生草區處理。
1.3.3 土壤蒸發 2021年4-5月用自制小型蒸發器Micro-lysimeter(由PVC管制成,內徑8.5 cm,壁厚5.0 mm,高度15.0 cm)測定蘋果樹行間中線上土壤水分日蒸發量。測定時,將其垂直壓入距牧草10.0 cm的行間土壤內取原狀土(為防止土壤掉入蒸發器內,壓入時保持0.5 cm高出地面),用膠帶封底后套于外管(由PVC管制成,內徑10.5 cm,壁厚5.0 mm,高度15.0 cm)中并固定于行間。每天16:00用精度為0.1 g的電子天平稱重,2 d內重量的差值為土壤日蒸發量,每隔3~5 d更換蒸發器內的原狀土(下雨后立即更換)。土壤日蒸發量由兩次稱重之間的差值和小型蒸發器底面積換算得出。每處理生草區隨機選取4個采樣點進行監測。
1.3.4 空氣相對濕度 2021年4-5月采用路格溫濕度記錄儀(L92-1,中國杭州)監測蘋果樹冠層1 m高度空氣相對濕度。將記錄儀固定于果樹主干1 m處,每隔1 h記錄1次數據,取24 h平均值。每處理生草區隨機選取4個采樣點進行監測。
1.3.5 土壤含水量和貯水量 2021年4-5月每隔10 d采用烘干稱重法測定0~100 cm土層土壤重量含水量并計算土壤貯水量。土壤貯水量(W,mm)=土壤重量含水量(ω,%)×土壤容重(ρ,g·cm-3)×土層深度(h,cm)×10。每處理生草區隨機選取4個采樣點進行監測,每個采樣點每個土層重復測定2次,取平均值。
1.3.6 一年生牧草還田時的生物學特征 2021年5月5日在小黑麥、二月蘭、毛葉苕子越年生牧草生草區取樣測定生物量。每處理生草區隨機選取4個采樣點進行測定,每個采樣點重復測定2次。每個采樣點每次重復齊地皮刈割相鄰4行1 m長牧草地上部分并稱總鮮重,稱完后隨機取500 g鮮草樣放入恒溫烘箱中65℃烘干至恒重,稱取干重計算干鮮比,根據干鮮比計算牧草地上生物量和含水量。將烘干的植物樣品粉碎后測量植株全碳(C)含量、全氮(N)含量[25],計算牧草植株的C/N。
1.3.7 蘋果產量與果實品質 2021年10月26日進行采收、測產。每生草區隨機選取20株果樹進行單株稱重測產,重復2次,測產時避開授粉樹行。每株測產后隨機選取20個蘋果果實測定品質,分別采用PAL-1折光儀(日本)、蒽酮比色法、NaOH滴定法測定可溶性固形物含量、可溶性糖含量和含酸量[25]。
1.4 氣象數據
來自武邑縣氣象局設在武羅綜合示范園區的標準氣象站。
1.5 數據處理
采用SAS 9.1統計軟件進行數據統計分析,用Excel 2013進行數據整理和作圖。
2 結果與分析
2.1 試驗區日均溫度和降水量變化
圖1顯示了武羅試驗區2021年和2000-2020年4-5月多年平均日均溫度和降水量變化情況。2000-2020年4-5月多年平均日均最高溫度為24.48℃,日均最低溫度為11.93℃,二者相差12.55℃,表明春季北方氣溫變化劇烈。2021年4-5月日均最高溫度為25.78℃,日均最低溫度為10.57℃,二者相差15.21℃,表明2021年春季氣溫變幅大于多年平均值。2000-2020年多年平均4、5月累積降水量分別為29.47、35.97 mm,2021年4、5月累積降水量分別為7.40、2.90 mm,二者分別相差22.07、33.07 mm,表明2021年4、5月在歷史上屬于極度干旱月份。
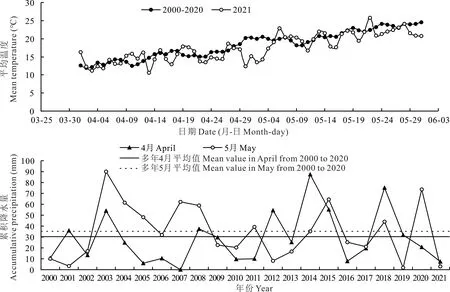
圖1 武羅試驗區2021年和2000-2020年4-5月多年平均氣溫和累積降水量Fig.1 Mean temperature and accumulative precipitation of April and May of 2021 and average from 2000 to 2020 in Wuluo experiment site
2.2 行間生草種類對果園土壤溫度的影響
從圖2可見,與清耕(CK)相比,越年生牧草和多年生牧草5、10、15、20、25 cm不同土層土壤溫度均顯著下降(P<0.05),說明行間生草阻隔了太陽光對果園地表的直接輻射,能顯著降低不同深度的土壤溫度。越年生牧草不同土層的降溫效應均大于多年生牧草,越年生牧草5、10、15、20、25 cm土層的降溫效應平均分別比多年生牧草高5.9、2.9、2.1、1.3、1.5℃。越年生牧草中毛葉苕子對5、10 cm土層土壤的降溫效應顯著低于二月蘭和小黑麥,這可能與毛葉苕子4月15日之前生長慢、地上生物量少、LAI顯著低于二月蘭和小黑麥、PAR透射率明顯增加有關。
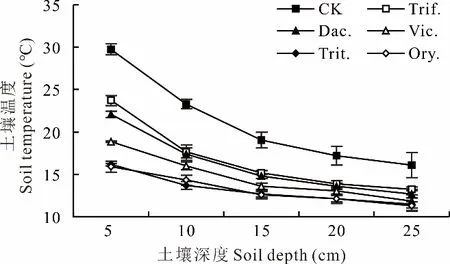
圖2 行間生草種類對果園0~25 cm土層土壤溫度的影響(2021年4月15日13:30-14:30)Fig.2 Effects of different grass species on the soil temperature of 0-25 cm profile in orchard(13:30-14:30,April 15,2021)
2.3 行間生草種類對牧草群體LAI、PAR透射率的影響
一定范圍內,作物群體LAI與冠層截獲的太陽輻射呈正相關關系[26-27],說明LAI越大,冠層PAR透射率越低,太陽對地表的輻射強度減弱,有可能阻止地溫上升、降低土壤蒸發。不同種類牧草因物候期不同,群體冠層LAI、PAR透射率不同。由圖3可知,越年生牧草二月蘭、小黑麥、毛葉苕子的LAI分別是8.43、6.77、4.67,PAR透射率分別是0.98%、1.72%、6.80%,多年生牧草鴨茅的LAI為2.87,PAR透射率為26.37%,越年生牧草群體LAI均顯著大于多年生牧草(P<0.05),而PAR透射率則顯著低于多年生牧草(P<0.05)。越年生牧草中二月蘭、小黑麥、毛葉苕子LAI差異顯著。毛葉苕子PAR透射率顯著高于二月蘭、小黑麥,二月蘭、小黑麥之間PAR透射率差異不明顯。
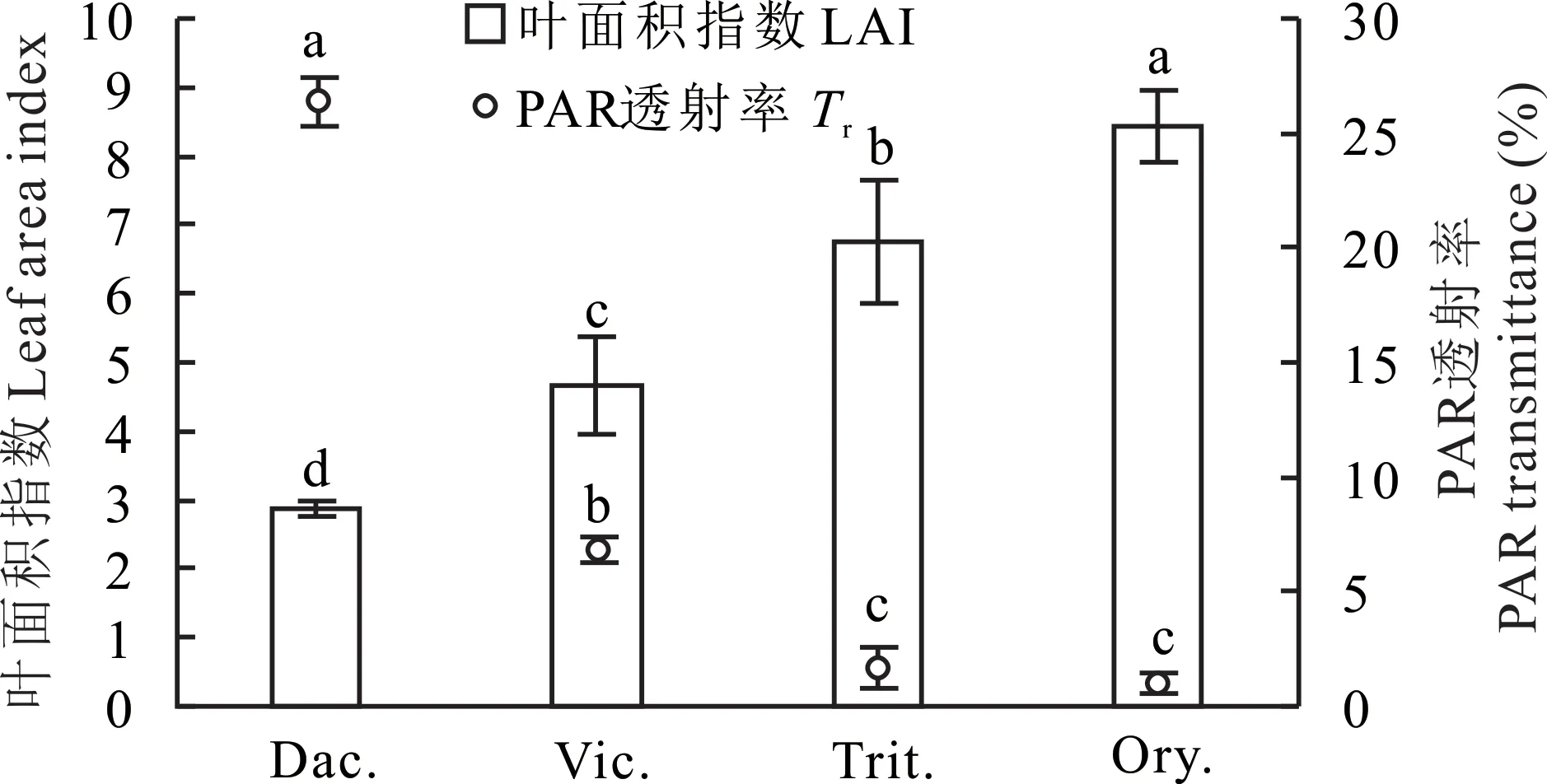
圖3 行間生草種類對牧草群體LAI、PAR透射率的影響(2021年4月15日11:00-14:00)Fig.3 Effects of different grass species on the grass population’s LAI and PAR transmittance(11:00-14:00,April 15,2021)
2.4 行間生草種類對果園土壤蒸發的影響
土壤蒸發強度與溫度呈正相關關系[19]。隨時間的推進和氣溫的上升,2021年4-5月果樹行間土壤日蒸發量和累積蒸發量均呈增加的趨勢(圖1和圖4)。行間生草均能顯著抑制土壤蒸發,生草種類不同對土壤蒸發抑制的效果不同(圖4)。與CK相比,越年生牧草二月蘭、小黑麥、毛葉苕子4-5月日蒸發量分別降低0.44、0.47、0.40 mm·d-1,累積蒸發量分別降低26.67、28.71、24.17 mm,多年生牧草白三葉、鴨茅4-5月日蒸發量分別降低0.50、0.47 mm·d-1,累積蒸發量分別降低30.4、28.9 mm。總體來看,多年生牧草土壤的日蒸發量、累積蒸發量的降幅均大于越年生牧草,平均分別降低0.06 mm·d-1、3.14 mm,該結果與越年生牧草切碎覆蓋還田(2021年5月6日)有關。
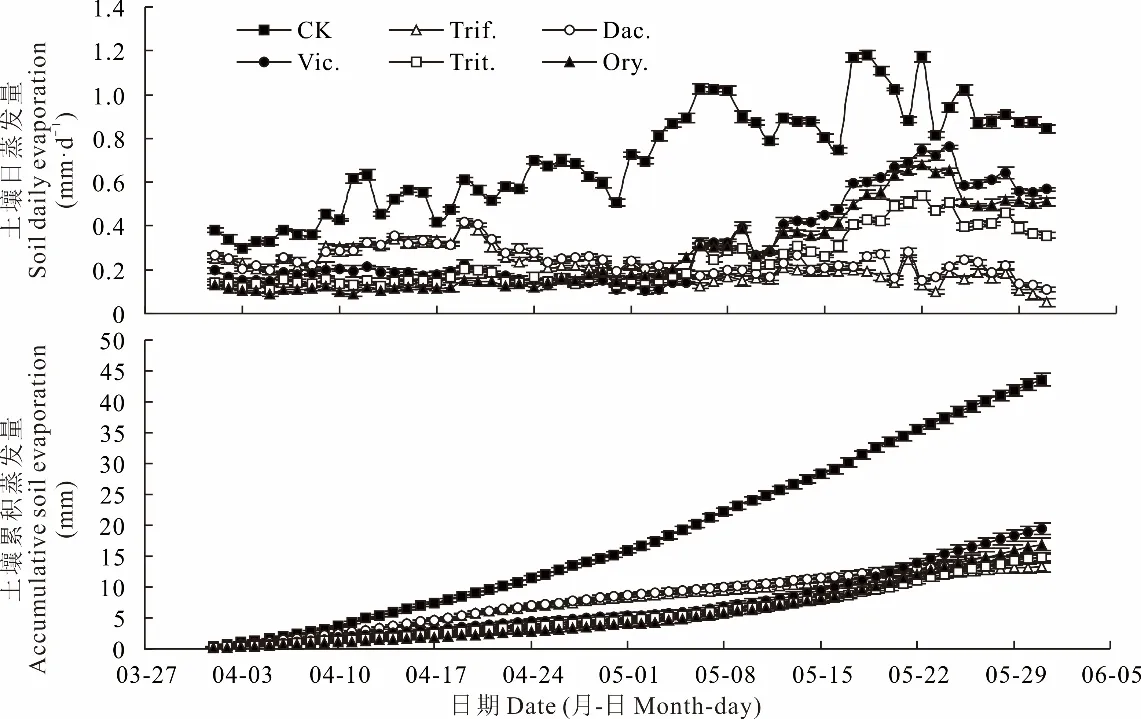
圖4 行間生草種類對蘋果樹行間土壤日蒸發量和累積蒸發量的影響Fig.4 Effects of different grass species on the soil daily evaporation and accumulative soil evaporation between apple tree rows
還田之前,與CK相比,越年生牧草二月蘭、小黑麥、毛葉苕子土壤日蒸發量分別減少0.41、0.40、0.38 mm·d-1,累積蒸發量分別減少14.42、13.90、13.38 mm,多年生牧草白三葉、鴨茅土壤日蒸發量分別減少0.29、0.28 mm·d-1,累積蒸發量分別減少10.15、9.76 mm,越年生牧草土壤的日蒸發量、累積蒸發量的降幅平均分別比多年生牧草高0.11 mm·d-1、3.94 mm,說明還田之前越年生牧草抑制土壤蒸發的效果比多年生牧草強,這可能與越年生牧草生長快、群體LAI大、PAR透射率低、地溫上升慢相關[28-29]。
還田后,與CK相比,越年生牧草二月蘭、小黑麥、毛葉苕子土壤日蒸發量分別減少0.47、0.57、0.42 mm·d-1,累積蒸發量分別減少26.67、28.71、24.17 mm,多年生牧草白三葉、鴨茅土壤日蒸發量分別減少0.78、0.74 mm·d-1,累積蒸發量分別減少30.36、28.95 mm,多年生牧草土壤的日蒸發量、累積蒸發量的降幅平均分別比越年生牧草高0.27 mm·d-1、3.14 mm,說明還田后越年生牧草抑制土壤蒸發的效果明顯小于多年生牧草。
2.5 土壤蒸發量與土壤溫度、牧草群體LAI、PAR透射率、土壤含水量的相關性分析
由表1可見,土壤蒸發量與牧草群體LAI呈極顯著的負相關關系,與PAR透射率和5、10、15、20、25 cm土層土壤溫度均呈極顯著的正相關關系,說明隨牧草生長,牧草群體LAI增加能顯著降低PAR透射率,減弱太陽對地表的輻射強度,阻止春季地溫上升,進而抑制土壤蒸發。土壤含水量也是影響土壤蒸發的一個重要因素[17]。由圖5可見,果園土壤日蒸發量與0~20 cm土層土壤含水量呈顯著的線性正相關關系,說明行間生草消耗土壤水分的同時會降低土壤蒸發量。行間生草種類不同對土壤水分消耗和土壤蒸發的影響不同。還田之前,越年生牧草抑制土壤蒸發的效果大于多年生牧草,可能與其對土壤水分消耗較多、降低土壤含水量有關。
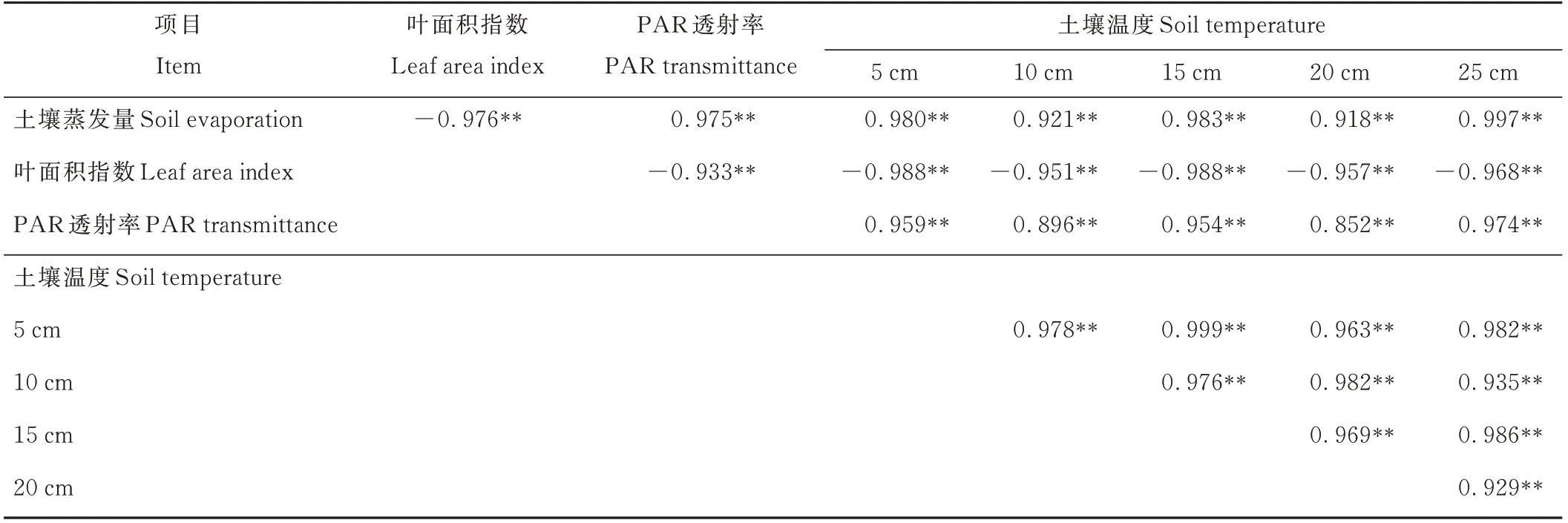
表1 土壤蒸發量、土壤溫度、牧草LAI及PAR透射率之間的相關性Table 1 The correlation between the indicators of soil evaporation,soil temperature,grass LAI and PAR transmittance
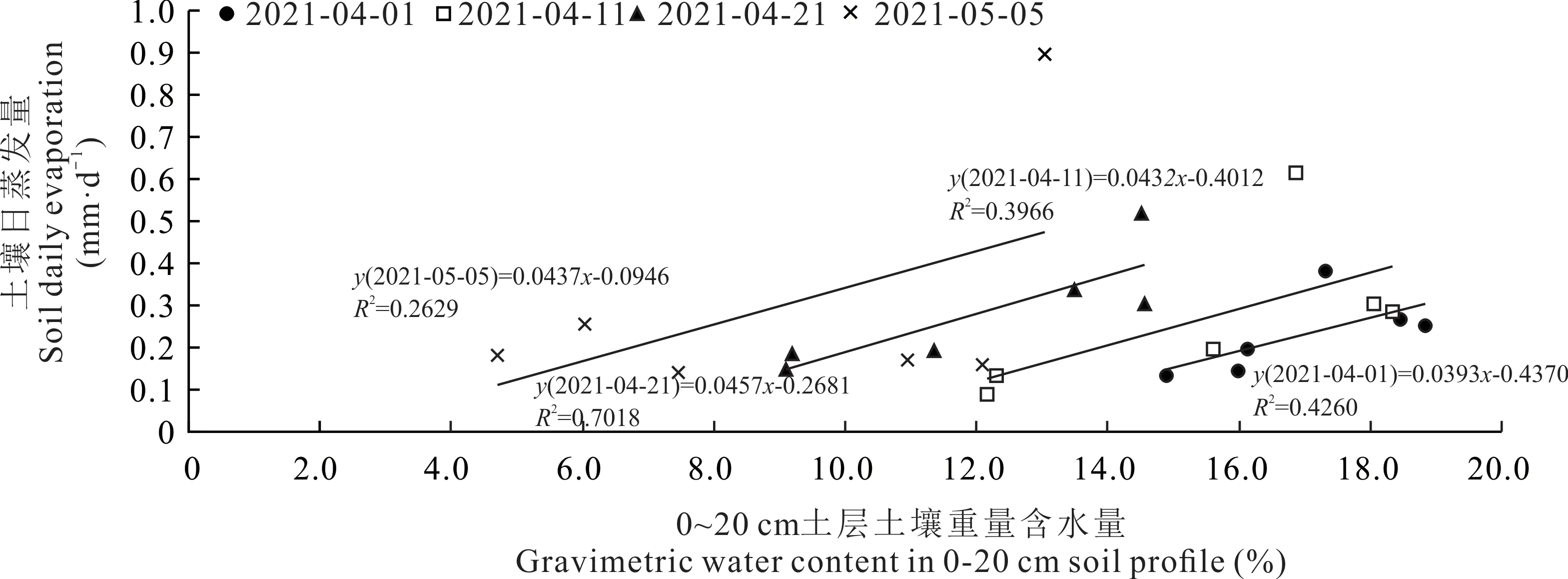
圖5 2021年4月1日-2021年5月5日土壤日蒸發量與0~20 cm土層土壤重量含水量的關系Fig.5 Relation of soil daily evaporation and gravimetric water content of 0-20 cm soil profile from 2021-04-01 to 2021-05-05
2.6 越年生牧草粉碎覆蓋還田時生物學性狀的比較
由表2可知,越年生牧草被切碎覆蓋還田時,禾本科作物小黑麥干重比豆科作物毛葉苕子、十字花科作物二月蘭分別多8.20、9.25 mg·hm-2,C/N分別高23.40、15.58,差異顯著(P<0.05)。3種越年生牧草還田時植株體內絕對含水量顯著不同,其中,小黑麥植株絕對含水量最低,分別比毛葉苕子、二月蘭低280.92%、70.66%,還田后小黑麥植株因含水量低、C/N高不易腐解,導致植株體內水分散失慢,有可能對果園空氣濕度的影響小于毛葉苕子、二月蘭。
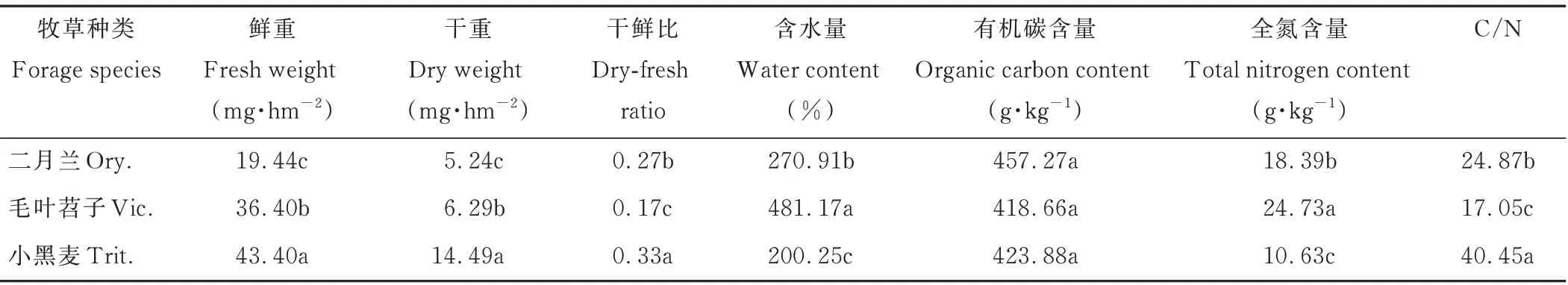
表2 越年生牧草還田時生物學性狀的比較Table 2 Comparison of biological characters of annual grasses when mulching returning
2.7 行間生草種類對蘋果樹冠層相對空氣濕度的影響
果園生草對果樹冠層空氣濕度的調控主要來源于下墊面牧草群體的蒸騰散水,而不同種類牧草群體蒸騰散水能力不同,對果樹冠層空氣濕度的調控效果表現不同。由圖6可見,行間種植越年生牧草和多年生牧草均能增加蘋果樹冠層1 m高度的空氣相對濕度。與CK相比,2021年4-5月越年生牧草二月蘭、小黑麥、毛葉苕子的每日空氣相對濕度平均分別增加24.51%、22.47%、19.36%,多年生牧草白三葉、鴨茅平均分別增加5.32%、7.99%,越年生牧草的每日空氣相對濕度比多年生牧草平均多增加15.46%,說明行間種植越年生牧草對春季果樹冠層1 m高度空氣相對濕度的改善效果大于多年生牧草。不同時間段,不同種類牧草對蘋果樹冠層1 m高度的空氣相對濕度影響不同。4月12日之前,與CK相比,白三葉、鴨茅和毛葉苕子對蘋果樹冠層1 m高度空氣相對濕度影響不明顯,而二月蘭、小黑麥影響顯著(P<0.05),二月蘭分別比白三葉、鴨茅、毛葉苕子的空氣相對濕度平均增加24.58%、24.28%、21.62%,小黑麥分別平均增加27.10%、26.81%、24.14%,這可能與4月12日之前白三葉、鴨茅、毛葉苕子返青慢、蒸騰作用弱相關。4月12日之后,毛葉苕子生長加快,與二月蘭、小黑麥蘋果樹冠層1 m高度空氣相對濕度差距顯著縮小。與CK相比,5月6日-5月8日小黑麥、二月蘭、毛葉苕子果樹冠層1 m高度空氣相對濕度變化較小,這與越年生牧草被切碎覆蓋還田不能蒸散水分相關。5月9日-5月25日,與清耕、多年生牧草相比,越年生牧草能顯著增加蘋果樹冠層1 m高度的空氣相對濕度。與CK相比,二月蘭、小黑麥、毛葉苕子處理平均分別增加34.64%、26.92%、37.90%;與白三葉相比,平均分別增加25.88%、18.16%、19.14%;與鴨茅相比,平均分別增加21.86%、14.14%、25.12%,這可能與越年生牧草切碎還田后植株殘體水分大量散失有關。越年生牧草中,小黑麥對蘋果樹冠層1 m高度空氣相對濕度的影響小于二月蘭和毛葉苕子。
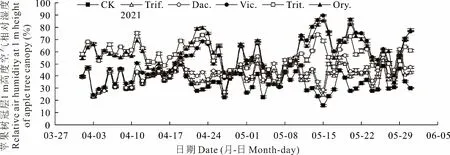
圖6 行間生草種類對蘋果樹冠層1 m高度空氣相對濕度的影響Fig.6 Effects of different grass species on the relative air humidity at 1 m height of apple tree canopy
2.8 行間生草種類對土壤含水量、貯水量的影響
牧草群體通過蒸騰散水改善果樹冠層空氣濕度,蒸騰散水來自于牧草根系對土壤水分的吸收,因此生草種類不同對果樹冠層空氣濕度的調控效果不同,進而對土壤含水量和貯水量變化的影響不同。圖7表明,隨時間推進,不同處理0~100 cm土層土壤含水量呈下降的趨勢,這與氣溫升高牧草生長加快消耗土壤水分相關。其中,白三葉根系吸水層主要發生在0~40 cm土層,鴨茅、毛葉苕子、二月蘭主要在0~60 cm土層,小黑麥主要在0~100 cm土層。2021年4月1日和4月11日多年生牧草白三葉、鴨茅0~20 cm、20~40 cm、40~60 cm、60~80 cm土層土壤含水量均高于CK,越年生牧草小黑麥、二月蘭均低于CK,毛葉苕子與CK差異較小,說明4月11日之前白三葉和鴨茅尚未完全返青對土壤水分消耗少,枯草層覆蓋地表能抑制土壤蒸發,具有保墑作用,而小黑麥處于拔節期、二月蘭處于盛花期,二者均處于快速生長階段,對土壤水分消耗較大。4月21日之后,各種牧草均進入快速生長階段,越年生牧草不同土層含水量均低于多年生牧草,二者之間的差距呈增加的趨勢,5月5日二者差距最大,毛葉苕子、小黑麥、二月蘭0~100 cm土層平均含水量分別比白三葉低3.70%、7.94%、5.57%,比鴨茅平均分別低1.69%、5.92%、3.56%。越年生牧草中小黑麥0~100 cm土層平均含水量最低,分別比毛葉苕子、二月蘭低4.23%、2.37%。
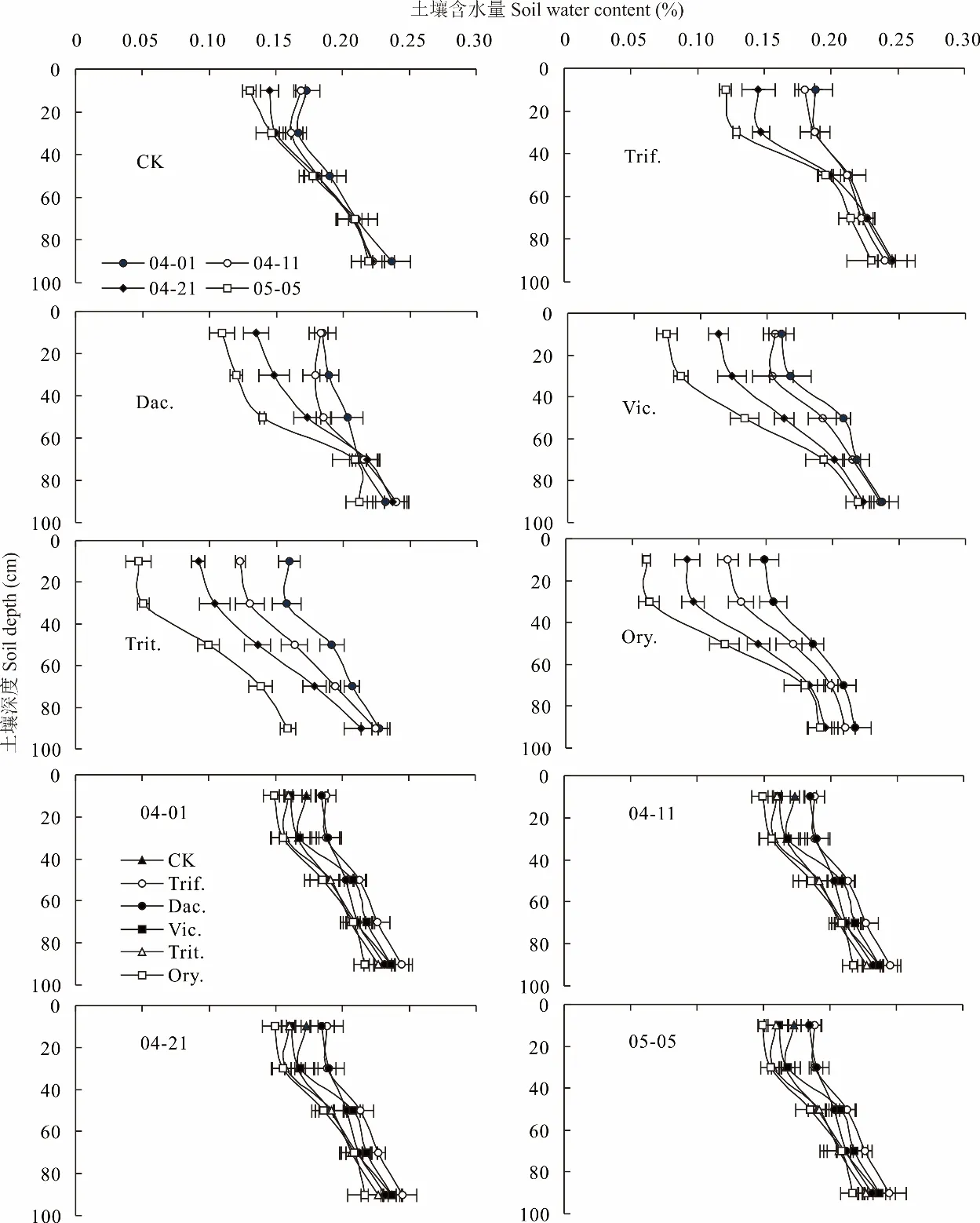
圖7 行間生草種類對果園0~100 cm土層土壤含水量的影響Fig.7 Effects of different grass species on the soil water contents of 0-100 cm soil profile in apple orchard
行間生草均消耗土壤水分,降低土壤貯水量。不同生草種類對土壤貯水量的影響不同(圖8)。與CK相比,多年生牧草白三葉、鴨茅1 m土體內土壤貯水量分別降低20.93、36.97 mm,其中白三葉降低不明顯,鴨茅顯著降低,越年生牧草毛葉苕子、小黑麥、二月蘭分別降低77.99、121.79、82.61 mm,差異均顯著(P<0.05)。越年生牧草1 m土體內土壤貯水減少量均高于多年生牧草,其中,與白三葉相比,小黑麥、二月蘭土壤貯水量顯著降低;與鴨茅相比,小黑麥土壤貯水量顯著降低,這可能與春季越年生牧草生長迅速、蒸騰耗水量大相關。2021年4月1日至5月5日蘋果樹1 m冠層空氣相對濕度與1 m土體內土壤貯水減少量呈線性顯著正相關關系,說明行間生草改善果園空氣濕度的水分主要來自牧草對土壤貯水的消耗,且二者具有正向協同關系。
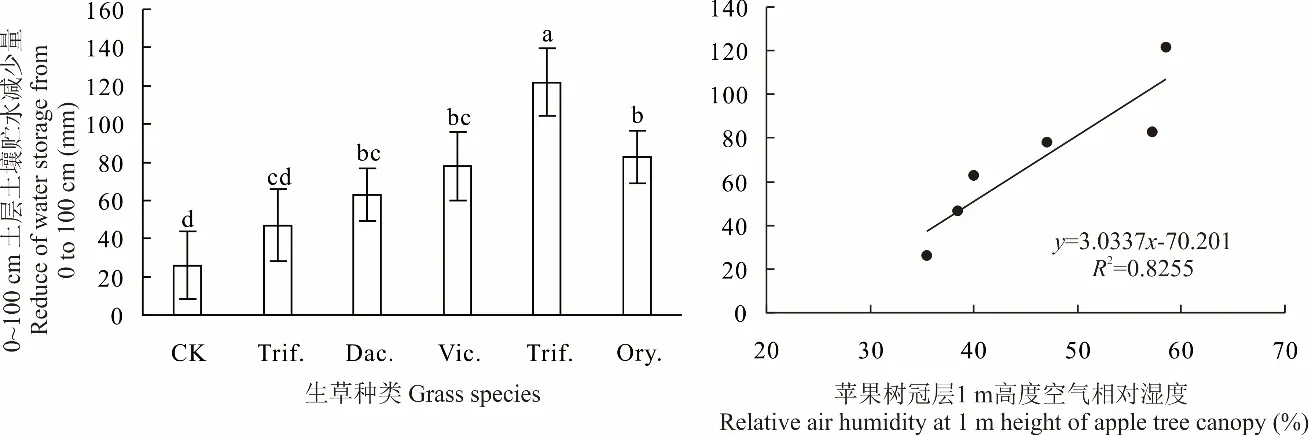
圖8 2021年4月1日-2021年5月5日果園0~100cm土層土壤貯水量的變化及與空氣相對濕度的關系Fig.8 Changes of the soil water storage of 0-100 cm soil profile in apple orchard and related with the air humidity from 2021-04-01 to 2021-05-05
2.9 行間生草種類對蘋果產量與果實品質的影響
由表3可見,行間生草均降低了蘋果產量,平均每株降低4.07 kg。與CK相比,行間種植多年生牧草白三葉、鴨茅蘋果產量下降不顯著(P>0.05),種植越年生牧草二月蘭、毛葉苕子、小黑麥下降顯著(P<0.05),其中,小黑麥下降幅度最大,每株降低7.06 kg。行間生草提高了蘋果果實可溶性固形物含量、可溶性糖含量、糖酸比,顯著降低了含酸量,說明行間生草能改善蘋果果實品質。
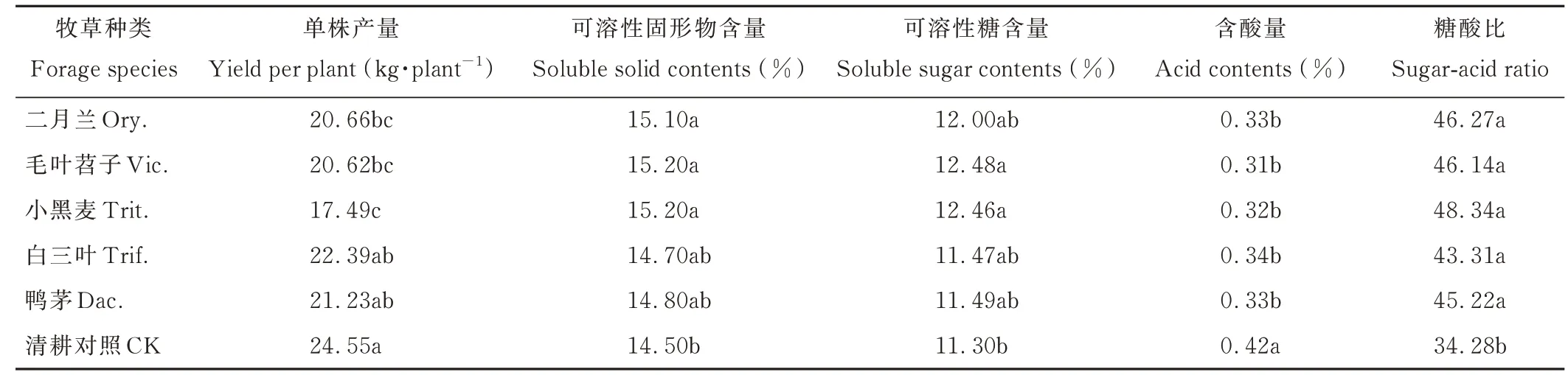
表3 行間生草種類對蘋果產量與果實品質的影響Table 3 Effects of different grass species on the yield and fruit quality of apple
3 討論
3.1 行間生草種類對果園土壤蒸發的影響
本研究結果表明,行間生草能抑制春季果園土壤蒸發,這與行間生草改變了果園下墊面性質,影響了地表與大氣能量、熱量的交換相關[4,14]。牧草冠層物理阻隔了太陽對地表的直接輻射,減弱了地表對太陽能量的吸收,降低了土壤溫度。眾多研究表明,植被LAI能抑制土壤蒸發[15],太陽輻射、土壤溫度能促進土壤蒸發[30-31]。本研究證實,土壤蒸發與牧草群體LAI呈顯著負相關關系,與PAR透射率、土壤溫度呈顯著正相關關系,這與孫宏勇等[15]、胡兵輝等[19]、李會科[32]的研究結果相一致。行間生草種類不同,在華北平原春季的生育進程不同,處于不同物候期的牧草群體葉面積指數、PAR透射率、土壤溫度均表現不同,將對土壤蒸發產生不同的抑制效果。本試驗中越年生牧草二月蘭、小黑麥、毛葉苕子均比多年生牧草白三葉、鴨茅萌發早、生長快,導致越年生牧草群體LAI均顯著高于多年生牧草,PAR透射率、0~25 cm土層土壤溫度均顯著低于多年生牧草。同時,越年生牧草生長進程快會消耗大量土壤水分,引起土壤表層含水量下降,抑制土壤蒸發,劉悅等[17]的研究結果也證實了這一點。本研究結果表明,越年生牧草被切碎覆蓋還田之前,蘋果樹行間種植越年生牧草二月蘭、小黑麥、毛葉苕子土壤的日蒸發量和累積蒸發量均低于多年生牧草鴨茅、白三葉,即越年生牧草二月蘭、小黑麥、毛葉苕子對果園土壤蒸發的抑制效果好于多年生牧草鴨茅、白三葉;而還田之后,越年生牧草對果園土壤蒸發的抑制效果小于多年生牧草,這可能與還田后越年生牧草物理阻隔太陽輻射的效果降低有關。
已有研究表明,秸稈還田可以抑制土壤蒸發,秸稈還田量是影響土壤蒸發的一個重要因素[33-34]。本研究中越年生牧草被切碎覆蓋還田之后,越年生牧草二月蘭、小黑麥、毛葉苕子土壤的日蒸發量和累積蒸發量仍明顯低于清耕,說明牧草秸稈切碎覆蓋還田具有抑制土壤蒸發的效果。3種越年生牧草中,小黑麥覆蓋還田后土壤日蒸發量明顯低于二月蘭和毛葉苕子,這可能與小黑麥還田時生物量大有關。因為李艷等[35]研究表明,秸稈覆蓋對土壤蒸發的阻力隨覆蓋量增加呈指數增加。本試驗中,還田時小黑麥地上生物量分別比毛葉苕子、二月蘭多8.20、9.25 mg·hm-2,差異顯著。同時,秸稈含水量顯著低于毛葉苕子、二月蘭,C/N顯著高于毛葉苕子、二月蘭。秸稈含水量、C/N是影響植物秸稈腐解快慢的兩個重要因素,秸稈含水量低、C/N大不利于土壤微生物對植物殘體的分解[36-39],因此還田后小黑麥秸稈腐解速率慢于毛葉苕子、二月蘭,對土壤蒸發的抑制效果強于毛葉苕子、二月蘭。
3.2 行間生草種類對果園空氣濕度的影響
許多研究發現,牧草通過蒸騰散水能改善果園空氣濕度[4,40-41]。本研究表明,與清耕相比,行間生草后華北平原春季蘋果樹冠層1 m高度空氣相對濕度改善明顯,平均提高15.5%。但不同種類牧草對果樹冠層空氣相對濕度影響差異顯著,這可能與牧草群體LAI不同相關。Qin等[42]研究表明植物葉片蒸騰能力與LAI呈正相關關系。本研究中,在越年生牧草被切碎覆蓋還田之前,不同種類牧草因生育進程不同導致LAI差異顯著,越年生牧草二月蘭、小黑麥、毛葉苕子萌發早、生長快,LAI顯著高于多年生牧草白三葉、鴨茅,試驗結果也表明越年生牧草對蘋果樹冠層1 m高度空氣相對濕度的調節效果顯著高于多年生牧草,平均提高15.1%。但越年生牧草中二月蘭LAI高于小黑麥,蘋果樹冠層1 m高度空氣相對濕度卻低于小黑麥,這可能因為牧草的冠層結構、植株高度、種植密度對蘋果樹冠層空氣的濕度有影響[43-45],具體的影響機制還需要進一步的研究。在越年生牧草被切碎覆蓋還田之后,二月蘭、小黑麥、毛葉苕子對蘋果樹冠層1 m高度空氣相對濕度的影響仍然高于多年生牧草鴨茅和白三葉,這可能與還田時植株含有大量水分相關。還田后小黑麥對蘋果樹冠層1 m高度空氣相對濕度的影響小于毛葉苕子、二月蘭,這可能是因為小黑麥植株含水量低、C/N高不易腐解,導致植株體內水分散失較慢。土壤水分蒸發會對果園空氣濕度產生影響[46],本研究中小黑麥還田時較高的生物量顯著抑制了土壤蒸發,這也可能是導致其對果樹冠層空氣濕度影響小的一個因素。
3.3 行間生草種類對果園土壤水分的影響
作物蒸騰、土壤蒸發構成了農田耗水結構,嚴重影響著土壤-植物-大氣連續體(soil-plant-atmosphere continuum,SPAC)界面的水分傳輸和農田水分平衡[47]。果園生草因改變地表特性、牧草生長耗水而降低其所接受的太陽輻射及土壤含水量,減少土壤-大氣界面水分傳輸,抑制土壤無效蒸發,提高農田水分利用效率。較高的LAI既能更好抑制土壤蒸發,也能促進牧草蒸騰散水而有利于改善果園空氣濕度。經牧草群體蒸騰改善果園空氣濕度的水分主要來自根系對土壤水分的吸收。本研究表明,行間生草后春季土壤蒸發量降低27.8 mm,土壤貯水量減少52.5 mm,土壤蒸發減少量不能彌補牧草蒸騰失水而導致土壤含水量下降,加重土壤干旱,引起果樹減產,這與惠竹梅等[8]、李同川等[24]、趙政陽等[48]、張先來等[49]、李會科等[50]的研究結果一致。但也有研究表明[12],果園生草能提高土壤含水量,提高果樹產量,本研究結果與其不一致,可能與研究時間短,土壤理化性狀改善不明顯,不能增強土壤水分的調蓄能力有關。本研究條件下,行間種植白三葉土壤貯水、果樹產量下降均不明顯,可作為華北平原蘋果園行間生草的主推草種。雖然越年生牧草二月蘭、毛葉苕子、小黑麥能顯著改善果園空氣濕度,但因此引起的土壤水分脅迫會顯著降低果樹產量,因此,關于華北平原蘋果園種植越年生牧草時如何優化灌溉制度降低果樹減產有待深入研究。
4 結論
行間生草能抑制華北平原蘋果園春季土壤蒸發。越年生牧草二月蘭、小黑麥、毛葉苕子切碎覆蓋還田之前對土壤蒸發的抑制效果大于多年生牧草白三葉、鴨茅,主要與越年生牧草萌發早、生長快、群體LAI大、PAR透射率低、土壤溫度低、含水量低相關。還田后的越年生牧草二月蘭、小黑麥、毛葉苕子仍能抑制土壤蒸發,但抑制效果小于多年生牧草白三葉、鴨茅。
行間生草能顯著調節華北平原春季蘋果樹冠層1 m高度空氣相對濕度。無論還田前后,越年生牧草二月蘭、小黑麥、毛葉苕子對果樹冠層空氣濕度的調節效果均顯著高于多年生牧草白三葉、鴨茅。還田前,越年生牧草對空氣濕度調控能力強與其群體LAI大、蒸騰散水能力強相關。還田后,可能與其植株殘體含水率高,腐解過程中釋放大量水分有關。
通過行間生草調節果園空氣濕度會降低華北平原蘋果園春季土壤的含水量、貯水量。蘋果樹冠層1 m高度空氣相對濕度與1 m土體內土壤貯水減少量呈線性顯著正相關關系。因越年生牧草小黑麥、二月蘭、毛葉苕子對果園空氣濕度的影響顯著高于多年生牧草白三葉、鴨茅,其土壤含水量、貯水量降低的幅度明顯大于多年生牧草白三葉、鴨茅。果園生草后降低的土壤蒸發量不能彌補其用來改善果園空氣濕度對土壤的水分消耗量,會加重果園土壤干旱、降低果樹產量。推廣果園生草制時應根據不同種類牧草對果園干旱季節土壤水分的消耗特征優化果樹灌溉制度。
