樹突細胞miR-17調節初始CD4+T淋巴細胞分化Treg/Th17失衡機制的研究
2023-02-10 06:00:34盛云峰邱美華陳園園孫麗芳甄利波
中國防癆雜志 2023年1期
關鍵詞:研究
盛云峰 邱美華 陳園園 孫麗芳 甄利波
結核分枝桿菌感染后會啟動依賴于細胞介導的適應性免疫應答,通過抗原提呈細胞如樹突細胞(dendritic cells,DC)將細菌抗原提呈給初始T細胞,進而分化為效應性T細胞。當再次接觸結核分枝桿菌抗原后,通過分泌多種炎癥介質,增強巨噬細胞功能從而殺滅結核分枝桿菌。但有研究發現結核分枝桿菌感染后機體的這種適應性免疫應答可發生失調,導致結核分枝桿菌不能被完全清除[1]。微小RNA(miRNA)是一類內源性非編碼RNA分子,為表觀遺傳調控機制之一。研究表明,miRNA參與調節人體各種免疫過程,在人類的腫瘤及自身免疫性疾病中已經發現多種miRNA的表達改變[2-3]。基于Solexa測序和qPCR方法的研究發現,肺結核患者的血清miRNA-17(簡稱“miR-17”)的表達顯著增加[4]。筆者通過研究肺結核患者的DC內miR-17的表達及肺結核患者的DC對初始CD4+T淋巴細胞分化的作用,探討miR-17參與調控結核分枝桿菌感染宿主的免疫失衡的相關機制。
對象和方法
一、 研究對象
選擇2022年2月1日至4月30日在浙江大學醫學院附屬杭州市胸科醫院收治的肺結核患者20例(肺結核組),同時收集同期門診的健康體檢者20名(健康對照組)。研究經浙江大學醫學院附屬杭州市胸科醫院倫理委員會批準,批準文號:[2022]研審第(62)號。肺結核組:男性12例、女性8例,年齡為22~42歲;健康對照組:男性11例、女性9例,年齡為24~38歲。肺結核患者的診斷均符合《WS 288—2017肺結核診斷》[5]的標準,并排除結核性胸膜炎和肺外結核。健康對照組入組標準:無肺結核臨床表現,無胸部影像學檢查異常。兩組研究對象均為浙江省的漢族人群,無合并其他傳染病、慢性病、腫瘤和自身免疫性疾病。
二、研究方法
1.主要試劑和儀器:人淋巴細胞分離液 Ficoll-Paque(美國 GE公司);初始 CD4+T淋巴細胞分離試劑盒;人T細胞活化劑CD3/CD28購自美國Thermo Fisher公司;胎牛血清購自美國Gibco公司;GM-CSF 購自美國R&D System公司;抗人Foxp3抗體購自英國AbCAM公司;Trizol液購自美國Invitrogen公司;miRNA提取試劑盒、cDNA逆轉錄試劑盒及mRNA 實時熒光定量PCR試劑盒均購自江蘇康為世紀生物科技有限公司;miR 陰性對照物(miR NC)、miR-17模擬物(miR-17 mimic)和miR-17抑制劑(miR-17 inhibitor)均購自上海吉瑪制藥技術有限公司。CO2細胞培養箱為Thermo Scientific(3111);倒置顯微鏡為Olympus(CX23);超凈工作臺為蘇信(YJ-840/YJ-1340);低速離心機為Eppendorff(5810R);分光光度計為Beckman(DU730);流式細胞儀為BD(Accuri C6);實時熒光定量PCR儀為Agilent(Stratagene MX3005p)。
2.健康對照組外周血初始CD4+T淋巴細胞的分離: 人濃縮白細胞加入磷酸鹽緩沖液(PBS),稀釋 2~4倍; 稀釋過的濃縮白細胞加入到15 ml的Ficoll 上, 400×g離心35 min,取出外周血單個核細胞層(PBMC); 加入PBS 300×g離心10 min洗滌1~2次。取 1×108的細胞加入0.4 ml PBS+BSA重懸細胞,再加入100 μl初始CD4+T淋巴細胞生物素-抗體Ⅱ混勻,4 ℃放置10 min。加入10~20 ml緩沖液,300×g離心10 min后去上清。加入0.8 ml純化緩沖液和 200 μl抗生物素微珠混勻,4 ℃放置15 min;加入10~20 ml緩沖液,300×g離心10 min,去上清。加入0.5 ml緩沖液重懸細胞,以LS柱分選獲得初始CD4+T淋巴細胞。
3.肺結核組外周血DC的分離與分化:取肺結核患者外周抗凝血10 ml轉入離心管中,吸取10 ml PBS稀釋并混勻,置于離心管中,1500×g離心10 min,去上清,加入1 ml紅細胞裂解液,靜置3 min,再加9 ml PBS溶液,1500×g離心10 min,棄上清,重懸細胞,加入1640培養基;第3天,1500×g離心5 min,棄上清,加入新鮮1640培養基(含10 ng/ml GM-CSF);第6天,1000×g離心5 min,棄上清,將細胞重新種植于150 mm直徑的培養皿中,加入新鮮1640培養基;第9天,1000×g離心5 min,棄上清,培養皿底部的松散貼壁細胞即為未成熟樹突細胞(imDC)。取部分上述細胞,加入PBS洗1次,1000×g離心3 min;調整細胞濃度為1×105細胞懸液1 ml,加入10 μl 抗人Foxp3抗體,室溫下避光孵育45 min,1 h內流式細胞儀檢測。將上述imDC按照5×106/ml密度,接種于100 mm 直徑的培養皿中,加入10 ml新鮮1640完全培養基,即可得到成熟DC。
4.成熟DC與初始CD4+T淋巴細胞共同培養:實驗分3組,miR陰性對照組、miR-17模擬物組和miR-17抑制劑組。將上述來源于肺結核組的成熟DC與來源于健康對照組的初始CD4+T淋巴細胞共培養于96孔板中,加入1640完全培養基(含人T淋巴細胞活化劑CD3/CD28);同時分別加入miR 陰性對照物、miR-17 模擬物、miR-17 抑制劑,將5 μl miRNA稀釋到500 μl的血清培養基中。將10 μl脂質體2000稀釋到500 μl的血清培養基中,輕輕混勻,靜置5 min。將miRNA溶液加入到脂質體2000溶液中,輕輕混勻,靜置20 min;將細胞培養板中的培養液吸出,重新加入5 ml新鮮的培養基(含10%胎牛血清的1640)。將兩種混合溶液加入到細胞培養皿中,37 ℃,5% CO2培養箱中培養48 h后收集細胞。將上述細胞調整到密度為1×105/ml,各取1 ml。固定和破膜后再分別進行抗Foxp3-488和抗IL-17A-PE標記。流式細胞儀檢測含有此兩種特異性標記的Treg細胞和Th17細胞的比例。
5.蛋白免疫印跡法檢測CD4+T淋巴細胞內蛋白Eos:收集上述細胞,于冰上裂解30 min。裂解完后,于4 ℃下12 000×g離心5 min。上清液轉移至新的潔凈EP管內進行定量。制膠完成后取適量樣品上清加入樣品孔中,旁孔加入預染的蛋白標記物。經過電泳-轉膜封閉后,進行一抗、二抗孵育。以三磷酸甘油醛脫氫酶(GAPDH)為內部對照。配制ECL發光液,加入化學反應底物,利用成像系統進行成像分析。
6. 檢測DC中miR-17及CD4+T淋巴細胞中Eos mRNA的基因表達水平:以江蘇康為世紀生物科技有限公司試劑盒對目標基因的mRNA表達量進行實時熒光定量PCR檢測,操作按照說明書。對miR-17的表達量進行檢測,以U6作為內部對照;對Eos mRNA表達量進行RT-PCR檢測,以GAPDH作為內部對照。相應引物序列見表1。反應條件:50 ℃ 2 min,95 ℃ 10 min,95 ℃ 15 s,60 ℃ 60 s,40個循環。繪制熔解曲線。采用2-△△Ct方法進行數據計算。

表1 實時熒光定量PCR引物
三、統計學處理
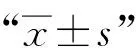
結 果
一、肺結核患者的DC中miR-17基因表達水平
利用miR-17及U6的特異性引物進行qPCR檢測miR-17的表達水平。結果發現,肺結核組DC的miR-17基因表達水平(12.546±1.572)明顯高于健康對照組DC的表達水平(2.409±1.097),差異有統計學意義(t=28.356,P<0.05)。
二、 轉染miR-17模擬物及抑制劑的DC對初始CD4+T淋巴細胞分化的影響
流式細胞儀檢測標記抗Foxp3-488和抗IL-17A-PE的初始CD4+T淋巴細胞。計算Treg細胞和Th17細胞的比例。miR-17模擬物組的Treg細胞比例[(4.740±0.901) %]明顯低于miR陰性對照組[(59.235±4.652) %],差異有統計學意義(t=50.755,P<0.01);miR-17模擬物組的Th17細胞比例[(67.610±3.495) %]明顯高于miR陰性對照組[(27.645±2.075) %],差異有統計學意義(t=38.521,P<0.01)。miR-17抑制劑組的Treg細胞比例[(83.080±5.770) %]明顯高于miR陰性對照組[(59.235±4.652) %],差異有統計學意義(t=14.988,P<0.01);miR-17抑制劑組的Th17細胞比例[(11.405±1.777) %]明顯低于miR陰性對照組[(27.645±2.075) %],差異有統計學意義(t=27.044,P<0.01)。
三、轉染miR-17模擬物及抑制劑的肺結核患者的DC對初始CD4+T淋巴細胞的Eos mRNA基因轉錄水平的影響
miR-17模擬物組的初始CD4+T淋巴細胞內EosmRNA基因轉錄水平(0.181±0.123)下調至38.4%,明顯低于miR陰性對照組(0.471±0.217),差異有統計學意義(t=10.449,P<0.05);miR-17抑制劑組的初始CD4+T淋巴細胞內EosmRNA基因轉錄水平(0.889±0.295)上調至1.887倍,明顯高于miR陰性對照組(0.471±0.217),差異有統計學意義(t=16.635,P<0.05)。
四、蛋白免疫印跡實驗檢測初始CD4+T淋巴細胞內蛋白Eos的表達
miR-17模擬物組的初始CD4+T淋巴細胞內Eos相對表達量(3.626±1.319)減少至77.7%,明顯低于miR陰性對照組(4.664±1.456),差異有統計學意義(t=8.528,P<0.05);miR-17抑制劑組的初始CD4+T淋巴細胞內Eos相對表達量(6.148±1.701)增加至1.318倍,明顯高于miR陰性對照組(4.664±1.456),差異有統計學意義(t=8.035,P<0.05)(圖1)。

注 *P<0.05; **P<0.01;GAPDH:三磷酸甘油醛脫氫酶;miR NC:miR陰性對照組;miR-17 mimics:miR-17模擬物組;miR-17 inhibitor:miR-17 抑制劑組圖1 miR-17模擬物和miR-17抑制劑轉染樹突細胞對初始CD4+T淋巴細胞蛋白Eos表達水平的影響
討 論
Th17細胞通過分泌白細胞介素-17A(IL-17A)、IL-17F、IL-21等多種炎癥因子介導免疫反應,可誘導中性粒細胞募集活化,參與機體對胞內菌感染的保護性免疫反應。Treg細胞是高表達CD25、低表達CD127的一組CD4+T淋巴抑制細胞,具有阻斷T淋巴效應細胞(Teff)的功能。Treg細胞能特異性表達具有穩定細胞功能的蛋白Foxp3。此外,還有其他特征性編碼基因(如Il2ra、Ctla4、Tnfrsf18、Ikzf2、Ikzf4等)在發揮Treg細胞的功能上也起著重要的作用[6-9]。在機體健康狀態下,Th17細胞與Treg細胞保持動態平衡。初始CD4+T淋巴細胞在轉化生長因子-β(TGF-β)、IL-6等多種炎癥因子的作用下可以分化為Treg細胞和Th17細胞。DC作為重要的抗原遞呈細胞,在機體適應性免疫應答中發揮著重要的作用[10]。有研究顯示,活動性肺結核患者體內Treg細胞數量明顯增多[11],與本研究的結果相一致。本研究還發現與肺結核患者的DC共培養的初始CD4+T淋巴細胞在DC受到miR-17基因抑制時其分化發生了偏移。提示miR-17在Treg/Th17分化失衡中發揮了一定作用,也可能參與了結核分枝桿菌感染的過程。
miRNA為長度約20~24個核甘酸的非編碼小分子,可與mRNA分子中的互補序列結合而切斷mRNA鏈,影響基因表達。miRNA還能通過縮短 poly(A)尾增加mRNA的不穩定性,也可降低核糖體翻譯mRNA的效率,從而導致基因沉默[12]。miRNA廣泛參與細胞基因表達、信號轉導、炎癥反應、腫瘤發生等多種細胞反應。由于miRNA穩定性高,且能被快速定量檢測,因此,可作為一種非侵襲性的生物標志物監測疾病的發生和發展[13]。當機體與結核分枝桿菌相互作用后,細胞會出現凋亡、自噬、極化和主要組織相容性復合物Ⅱ(MHCⅡ)類表達等反應。而這些細胞反應主要是由miRNA通過固有免疫和適應性免疫反應進行調節[14]。體內外研究發現,miRNA除了通過多種信號通路參與調節結核分枝桿菌感染后的機體免疫過程,還可能通過調節DC的免疫反應參與固有免疫和適應性免疫的轉換[15]。Tu等[16]研究發現,肺結核患者血清中miR-17的表達明顯增加。本研究也發現,肺結核患者的DC內miR-17的表達水平明顯升高,可作為結核分枝桿菌感染的生物標志物。有研究顯示,miR-17通過直接靶向酪氨酸蛋白激酶(janus kinase 1,JAK1) 和信號轉導及轉錄激活蛋白3(signal transducer and activator of transcription 3,STAT3)抑制IL-6家族自分泌而發揮抗炎作用[17];miR-17可以負性調節Toll樣受體4(TLR4),從而抑制巨噬細胞中炎癥因子IL-1β和腫瘤壞死因子-α(TNF-α)的濃度,減輕膿毒癥的炎癥反應[18]; miR-17 可以提高IL-18表達水平,導致中性粒細胞浸潤[19]; miR-17通過降低ATP結合盒轉運蛋白 A1(ABCA1)水平促進炎癥細胞聚集等[20],提示miR-17可能參與結核分枝桿菌感染機體后的一系列適應性免疫應答。
Eos是由位于第12號染色體(12q13.2)上的Ikzf4基因編碼的長約585aa的一種蛋白質,屬于Ikaros家族鋅指蛋白2中的一種。同時也是一種主要在Treg細胞中表達的轉錄因子,在維持Treg細胞穩定性和抑制性上起著關鍵性作用。有研究證實了卡介苗處理后在Treg細胞Eos基因中發現8個CpG高度去甲基化,從而更有利于促進Treg細胞的發育成熟[21]。另有研究發現,Eos與Treg細胞Foxp3形成復合物有助于維持Treg細胞的表型,且Eos下調能夠誘導Treg細胞向Th輔助細胞的轉化[22]。本研究發現,CD4+T淋巴細胞與轉染了miR-17模擬物的DC共培養后,其Eos基因表達減少,而抑制DC的miR-17表達后,共培養的CD4+T淋巴細胞的Eos基因轉錄及表達均明顯增加,Treg細胞分化比例明顯升高,表明miR-17可能通過調節Eos的表達影響CD4+T淋巴細胞的分化,進而調節Treg/Th17的平衡。這一結果提示Eos可能是miR-17控制Th17分化的靶點,通過抑制Eos增強Th17的分化。結核分枝桿菌感染后宿主復雜的適應性免疫應答過程中,miR-17究竟通過哪些信號通路進行調節尚需進一步研究。
綜上所述,結核分枝桿菌感染后導致miR-17表達增加,Eos基因參與Th17/Treg的分化,進而影響結核分枝桿菌引起的機體免疫反應。而這一免疫反應過程復雜多樣,其詳細的分子機制還需進一步研究。
利益沖突所有作者均聲明不存在利益沖突
作者貢獻盛云峰:設計實驗、分析數據和論文撰寫;邱美華:指導、設計和修改;陳園園和孫麗芳:數據采集和統計分析;甄利波:審閱修改
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
