METTL16在雞不同類型肌肉中的表達規律及其對肌肉功能的調控作用
2023-02-27 14:26:52龐立川單艷菊劉一帆甘達峰屠云潔姬改革巨曉軍束婧婷鄒劍敏
畜牧獸醫學報 2023年2期
龐立川,單艷菊,劉一帆,章 明,甘達峰,屠云潔,姬改革,巨曉軍,束婧婷*,鄒劍敏*
(1.江蘇省家禽科學研究所 江蘇省家禽遺傳育種重點實驗室,揚州 225125;2.崇仁縣畜牧良種場,撫州 344200)
隨著生活水平的不斷提高,人們對雞肉品質的要求也越來越高,如何提高雞肉品質已成為肉雞育種學者需要面對的關鍵問題。骨骼肌作為動物軀體最主要的組成部分,約占體重的40%[1]。肌纖維是骨骼肌的基本構成單位。肌纖維的形成是一個復雜的過程,由單核成肌細胞增殖融合形成多核肌管,最后肌管進一步分化形成。通常,骨骼肌群由具有不同理化特性的肌纖維混合組成,骨骼肌群中肌纖維類型的組成因部位和功能不同存在較大區別,并可能隨著生長發育和環境應激發生相互轉化[2]。研究表明,肌纖維類型與雞肉品質密切相關,總體上,慢肌纖維含量多的肌肉其肉品質要優于快白肌纖維含量多的肌肉[3]。因此,揭示雞骨骼肌肌纖維類型形成及其轉化機制對改良雞肉品質具有重要意義。
N6-甲基腺苷(m6A)修飾于1974年首次在大鼠的信使RNA中被發現[4]。m6A修飾是真核生物中含量最豐富的內部修飾,影響RNA的剪接、轉錄、翻譯、定位、代謝和穩定性[5]。m6A修飾廣泛存在并涉及多個生理過程,如胚胎發育、神經發生、性別決定和腫瘤發生等[6-8]。m6A修飾需要甲基轉移酶、去甲基化酶以及識別蛋白來發揮作用。甲基轉移酶METTL3和METTL14共同構成甲基轉移酶復合體的異二聚體核心,介導RNA特定位點的甲基化[9]。METTL3發揮主要的催化活性[10],METTL14不具有催化活性但可以促進METTL3與底物的結合[11]。已有很多研究發現m6A甲基化與肌肉生長發育存在聯系,但大多來自哺乳動物的研究。例如m6A甲基化酶METTL3通過提高Wnt通路的LEF1 mRNA甲基化水平,正向調控骨骼肌分化[12]。METTL3/14還可通過甲基化ERK信號通路的關鍵調節因子MNK2 mRNA,調節MNK2蛋白水平,反向調控骨骼肌分化[13]。去甲基化酶FTO可通過去甲基化GADD45BmRNA提高p38 MAPK磷酸化水平,促進骨骼肌分化[14]。
本課題組前期發現甲基化轉移酶METTL3和METTL14在雞不同類型肌肉組織中差異表達[15]。METTL16是繼METTL3和METTL14之后,近幾年新發現的另一種m6A甲基轉移酶[16-18]。為深入探究METTL16甲基化轉移酶在雞肌肉功能中的作用,本研究檢測了METTL16在雞不同類型肌肉組織中的表達水平,同時在體外干擾雞原代成肌細胞中的表達,分析METTL16敲低表達后對雞成肌細胞增殖的影響以及對肌細胞分化相關基因和肌纖維類型標志基因表達的影響。本研究結果為進一步闡明m6A甲基化修飾在雞骨骼肌中的作用提供理論依據。
1 材料與方法
1.1 試驗動物
本研究使用的種蛋來源于揚州翔龍禽業發展有限公司,置于實驗室孵化器(溫度設為37.5 ℃,濕度設為60%)孵化至11胚齡時用于雞成肌細胞提取。隨機選取體重變異系數在5%之內的120日齡(上市日齡)廣西麻雞母雞6只,采集胸大肌、胸小肌、縫匠肌、恥坐骨肌內側肌、恥坐骨肌外側肌、髂脛外側肌、腓腸肌內側頭肌和背闊肌組織樣,置于液氮速凍,然后轉入-80 ℃冰箱保存備用。
1.2 主要試劑
DMEM培養基、EDTA胰酶、胎牛血清、馬血清、Opti-MEMⅠReduced Serum Medium購自美國Gibco公司;CellCounting-Lite 2.0試劑、反轉錄試劑盒、熒光定量試劑盒購自南京諾唯贊生物科技股份有限公司;siRNA購自廣州銳博生物科技有限公司;LipofectamineTMRNAiMAX轉染試劑購自美國Invitrogen公司;RNase-free ddH2O、TRIzol購自天根生化科技(北京)有限公司;DEPC、氯仿、無水乙醇、異丙醇購自生工生物工程(上海)股份有限公司。
1.3 引物設計及合成
根據GenBank公布的雞METTL16及雞成肌細胞分化標志基因MyoD、MyoG和肌纖維分型標志基因MyHC以及內參基因β-actinmRNA序列,利用在線工具Primer 3設計引物,送由生工生物工程(上海)股份有限公司合成。引物信息見表1。

表1 實時熒光定量PCR引物信息
1.4 雞原代成肌細胞的分離、培養與誘導分化
取孵化11胚齡的雞胚,PBS清洗后,分離腿肌,將肌肉剪碎后,用含有EDTA的胰酶,置于37 ℃消化20 min后,用生長培養基(DMEM+20%胎牛血清和1%青/鏈霉素)終止消化,1 000 r·min-1,5 min離心后,細胞沉淀用生長培養基重懸,分別用200目、400目濾篩過濾,置于37 ℃、5% CO2培養箱中培養。利用差速貼壁法純化提取的雞成肌細胞,即培養40 min后,將細胞液轉入新的培養板中,并重復1次操作。待細胞匯合至80%時,換用分化培養基(DMEM+5%馬血清和1%青/鏈霉素)誘導分化。分別在培養0 h、培養12 h(增殖50%密度)、培養28 h(增殖100%密度)及誘導分化后1、3、5、7 d用TRIzol總RNA提取試劑收集雞成肌細胞。每組設4個重復。
1.5 siRNA轉染
siRNA(包括METTL16-RNA和NC-siRNA)由廣州銳博生物科技有限公司設計并合成。參照LipofectamineTM 3000轉染試劑說明書進行操作,將100 nmol·L-1METTL16-siRNAs和NC-siRNAs分別用Lip3000包裹成轉染復合物后,轉染雞成肌細胞,在Opti-MEM減血清培養基培養8 h后更換含有DMEM的增殖或分化培養液。
1.6 細胞增殖檢測
將原代成肌細胞以1×105個·孔-1的密度接種96孔板,放入CO2培養箱過夜培養后,借助Lip3000將METTL16-siRNA和NC-siRNA分別轉染細胞,每組6個重復。在轉染24 和48 h用CellCounting-Lite 2.0試劑檢測雞成肌細胞增殖情況。具體操作步驟按CellCounting-Lite 2.0試劑說明書進行。用酶標儀檢測發光值,按照說明書計算細胞增殖情況。
1.7 RNA提取及cDNA合成
使用TRIzol總RNA提取試劑分別提取胸大肌、胸小肌、縫匠肌、恥坐骨肌內側肌、恥坐骨肌外側肌、髂脛外側肌、腓腸肌內側頭肌和背闊肌等肌肉組織及雞成肌細胞的總RNA。NanoDrop1000微量分光光度計檢測總RNA的純度和濃度,-80 ℃保存備用。按照HiScript ⅢRTSuperMix反轉錄試劑盒操作程序進行cDNA合成,體系20 μL:4×gDNA wiper Mix 4 μL,5×HiscriptⅢ 4 μL,RNA模板1 000 ng,RNase-free ddH2O補足20 μL。反應程序:42 ℃ 2 min,50 ℃ 15 min,85 ℃ 2 min。cDNA于-20 ℃保存備用。
1.8 m6A甲基化水平檢測
采用比色法檢測成肌細胞m6A甲基化水平,具體步驟參考 m6A RNA Methylation Quantification Kit(Abcam,ab185912)說明書進行,反應結束后使用多功能酶標儀測量 450 nm 處的吸光度值,計算各樣品的RNA m6A含量,每組3個重復。計算公式如下:m6A(%)=((Sample OD-NC OD)/S)/((PC OD-NC OD)/P)×100,式中:S為RNA含量,P為陽性對照RNA含量。
1.9 實時熒光定量PCR反應
對反轉錄獲得的cDNA,使用ChamQ Universal SYBR qPCR Master Mix 熒光定量試劑盒進行定量分析。每個樣本設3個生物學重復。PCR反應體系20 μL:2X ChamQ Universal qPCR Master Mix 10 μL,上、下游引物(10 μmol·L-1)各0.4 μL,cDNA模板2 μL,RNase-free ddH2O補足體系。PCR反應程序:95 ℃預變性30 s;95 ℃變性10 s,56 ℃退火30 s,共40個循環。以β-actin為內參。
1.10 統計分析
實時熒光定量PCR結果采用2-ΔΔCt方法進行計算。將胸大肌設為不同部位肌肉組織中的表達對照組,ΔΔCt=ΔCt(其他部位肌肉)- ΔCt(胸大肌);細胞分化前后表達分析將培養0 h設為對照組,ΔΔCt=ΔCt(其他時間點)-ΔCt(0 h)。每個樣本至少4個重復;用SPSS26.0軟件進行統計學單因素方差分析表達量,組間差異采用獨立樣本t檢驗進行比較,用GraphPadPrism8.0作圖。P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結 果
2.1 METTL16基因在雞不同類型肌肉中的表達
對廣西麻雞胸大肌(XD)、胸小肌(XX)、縫匠肌(FJ)、恥坐骨肌內側肌(CN)、恥坐骨肌外側肌(CW)、髂脛外側肌(QW)、腓腸肌內側頭肌(FN)和背闊肌(BK)8種類型肌肉中METTL16基因表達情況進行檢測(圖1)。由圖1可見,METTL16基因在8種類型肌肉組織中均有表達,總體上,METTL16在白肌纖維為主的肌肉(XD、XX、QW、CW)中的表達要高于紅肌纖維為主的肌肉(FJ、CN、FN、BK)。其中,METTL16基因在XD和XX中表達量較高,顯著高于其它類型肌肉(P<0.05);其次在QW中表達較高,顯著高于其它5種類型肌肉(P<0.05);其它5種類型肌肉間表達差異不顯著(P>0.05)
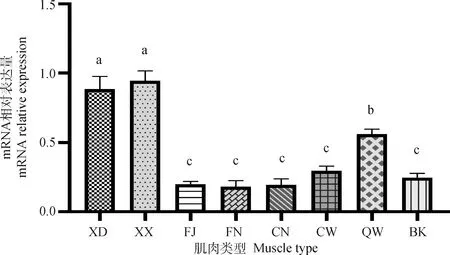
XD. 胸大肌;XX. 胸小肌;FJ. 縫匠肌;FN. 腓腸肌內側頭肌;CN. 恥坐骨肌內側肌;CW. 恥坐骨肌外側肌;QW. 髂脛外側肌;BK. 背闊肌。柱上不同字母表示差異顯著(P<0.05);柱上相同字母表示差異不顯著(P>0.05)。下同。n=6
2.2 METTL16基因在雞原代成肌細胞中的表達規律
METTL16基因在雞成肌細胞增殖和分化中的表達規律顯示(圖2),剛分離培養的雞原代成肌細胞中METTL16基因表達最高,隨著細胞的增殖,METTL16基因表達開始顯著下降(P<0.05);誘導分化1 d后,METTL16基因表達最低,隨后隨著細胞的分化METTL16基因表達開始顯著上升;分化5 d后,METTL16基因表達維持在相對穩定的水平。
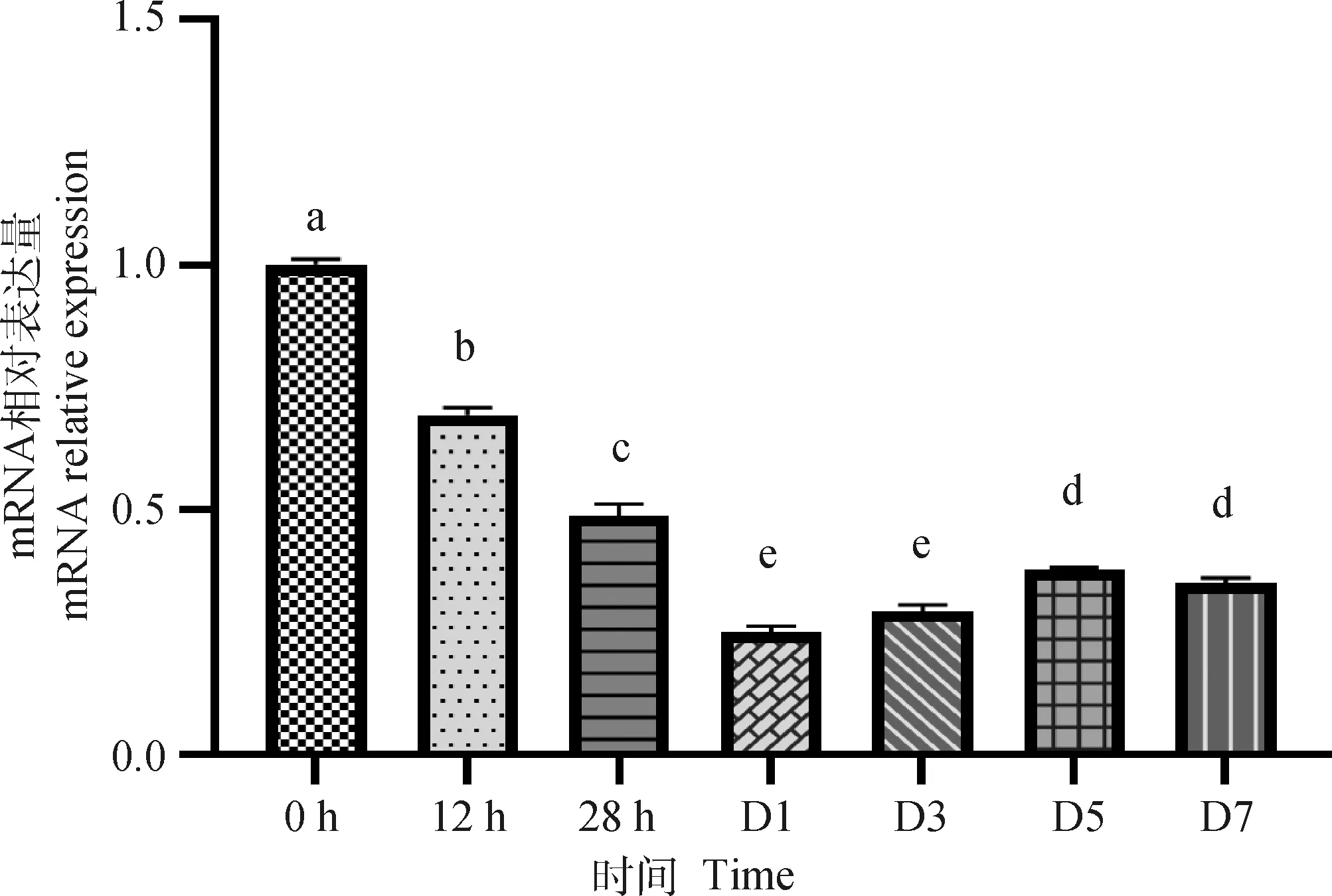
圖2 METTL16在雞原代成肌細胞增殖分化過程中的表達
2.3 干擾METTL16基因對雞成肌細胞增殖的影響
利用CellCounting-Lite 2.0增殖檢測試劑盒檢測干擾METTL16基因表達后雞成肌細胞的增殖情況。由圖3可見,干擾METTL16基因24 h時,雞成肌細胞干擾組的增殖活力開始低于NC對照組,干擾METTL16基因48 h后,干擾組的細胞增殖活力顯著低于NC組(P<0.05)。提示METTL16對雞成肌細胞的增殖有促進作用。

*. P<0.05,**. P<0.01.,***. P<0.001,下同
2.4 干擾METTL16基因對雞成肌細胞m6A甲基化水平的影響
在分化期干擾雞成肌細胞中METTL16基因表達后,采用比色法檢測雞成肌細胞RNA m6A甲基化水平。由圖4可見,與NC組相比,干擾組的甲基化水平呈現下降的趨勢。提示METTL16在雞成肌細胞分化期,正向調控RNA m6A甲基化水平。
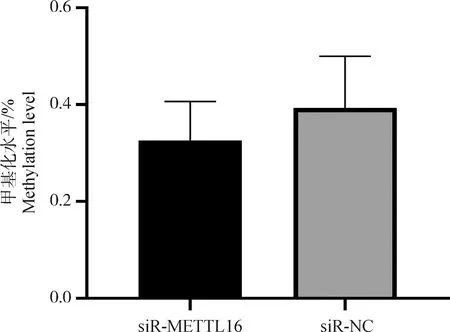
圖4 干擾METTL16對雞原代成肌細胞RNA m6A甲基化水平的影響
2.5 干擾METTL16基因對雞成肌細胞分化的影響
在分化期干擾雞成肌細胞中METTL16基因表達后的細胞形態如圖5。由圖5可見,各個時間點試驗組(METTL16)與對照組(NC)細胞和肌管形態沒有明顯區別。在分化期干擾雞成肌細胞中METTL16基因表達后,細胞分化及肌纖維類型相關基因表達結果見圖6,與NC組相比,干擾組中MyoD的表達顯著下降(P<0.05),MyoG的表達呈下降趨勢(P>0.05),fast-MyHC的表達極顯著上升(P<0.001),slow-MyHC的表達呈下降趨勢(P>0.05)。

A、B分別表示對照組分化期轉染si-NC 24與48 h后細胞形態;C、D表示試驗組分化期轉染si-METTL16 24與48 h后細胞形態

A.干擾METTL16后MyoD mRNA表達情況;B.干擾METTL16后MyoG mRNA表達情況;C.干擾METTL16后fast-MyHC mRNA表達情況;D.干擾METTL16后slow-MyHC mRNA表達情況
3 討 論
METTL16是RNA修飾領域的新成員,最初被認為是核糖體RNA甲基轉移酶。現有研究顯示METTL16通過m6A甲基化修飾前RNA(pre-RNA)和非編碼RNA(ncRNA)[19]以及長非編碼RNA(lncRNA)[20]調節小鼠細胞中MAT2A的mRNA水平,維持S-腺苷蛋氨酸(SAM)的穩態[21-22]。另外,METTL16不僅在細胞核內發揮m6A編寫器的作用,在胞漿中還以不依賴m6A的方式,通過其甲基轉移酶結構域(Mtase domain)直接與真核細胞起始因子3a(eIF3a)和3b(eIF3b)以及核糖體RNA相互作用,加快翻譯起始復合體的組裝,從而促進mRNA的翻譯[23]。有研究發現通過紫外線輻射造成的DNA損傷后,METTL16被招募到損傷區域并與附近的小RNA相關聯[24]。可見METTL16在m6A甲基化修飾過程中較重要。但METTL16在家禽中的研究報道還比較少。
課題組前期通過ATPase染色將8種類型肌肉分型,結果表明,胸大肌、胸小肌、恥坐骨肌外側肌和髂脛外側肌以白肌纖維為主;腓腸肌內側頭肌、背闊肌、縫匠肌和恥坐骨肌內側肌以紅肌纖維為主[15]。為研究METTL16在雞肌肉中的作用,本研究首先通過qRT-PCR對8種類型肌肉中METTL16基因表達情況進行檢測,發現METTL16基因在胸小肌等以白肌纖維為主的肌肉中的表達要高于縫匠肌等以紅肌纖維為主的肌肉。這與本課題組研究發現m6A甲基化轉移酶METTL3、METTL14、WTAP和VIRMA在紅肌纖維為主的肌肉中的表達量高于白肌纖維為主的肌肉的結果不一致[15]。同時對干擾METTL16基因后雞成肌細胞RNA m6A甲基化水平進行檢測表明,METTL16能夠促進雞成肌細胞RNA m6A甲基化水平的提高。提示METTL16表達水平與肌纖維類型分布相關,可能通過不同于METTL3等甲基化轉移酶的特殊的機制參與肌肉m6A調控。
通過對METTL16基因在雞成肌細胞中增殖和分化不同時期的表達水平檢測表明,METTL16基因表達水平隨著成肌細胞增殖逐漸降低,隨著成肌細胞分化逐漸升高,提示METTL16可能在成肌細胞增殖與分化過程中發揮作用。本課題組前期研究發現甲基轉移酶METTL3、METTL14和WTAP在雞成肌細胞增殖過程中表達上調,在雞成肌細胞分化的前4天表達下降,第5天表達回升[15]。丁浩等[25]研究發現,METTL3、METTL14和WTAP表達水平在細胞增殖時期逐漸上調。雞成肌細胞中METTL16基因表達的趨勢再次與METTL3等甲基化轉移酶基因表達趨勢相反,推測METTL16很可能通過不同于METTL3等甲基化轉移酶的特殊的機制參與肌肉m6A調控。
成肌細胞的增殖分化是個非常復雜的過程,受很多因子的調控。如骨骼肌的分化被證明受生肌調節因子家族MyoD、MyoG、Myf5和Mrf4等調控[26],MyoD與Myf5在成肌細胞早期分化發揮作用,而MyoG與Mrf4在成肌細胞后期分化為肌纖維的過程中發揮調節作用[27-28]。骨骼肌纖維類型的轉化歸根結底是構成肌纖維的蛋白多肽鏈結構發生了變化,是多核肌纖維內反映不同種類特征的基因得以表達的結果[29]。而不同亞型的MYHC基因是表征肌肉纖維類型多樣性的最佳標志[30]。最近的研究表明,m6A甲基化在骨骼肌的分化中發揮重要作用。甲基化轉移酶METTL3已被證實介導MyoD等肌源性轉錄因子mRMA的5′UTR的m6A修飾來調控骨骼肌分化[31],但還未見METTL16在骨骼肌中的作用報道。Wang[32]等報道,敲低METTL16可通過降低胃癌細胞m6A水平,降低cyclinD1 mRNA穩定性,進而降低cyclinD1的表達來抑制G1/S期細胞增殖。本研究利用CellCounting-Lite 2.0細胞增殖試劑檢測發現,干擾METTL16基因表達后,雞成肌細胞增殖受到抑制。本研究在肌細胞分化條件下干擾METTL16基因的表達,結果表明MyoD的表達顯著下降,MyoG的表達變化不顯著,fast-MyHC的表達極顯著上升,slow-MyHC的表達呈現下降趨勢。提示METTL16可能在雞成肌細胞的增殖、分化和肌纖維形成中發揮重要作用。但是METTL16是通過何種機制調控雞成肌細胞增殖分化和肌纖維形成的還需進一步研究。
4 結 論
METTL16在雞的不同類型肌肉中廣泛表達,其表達水平與肌纖維類型組成相關。干擾METTL16基因表達能夠明顯抑制雞原始成肌細胞的增殖和MyoD基因的表達,明顯促進fast-MyHC基因的表達,提示METTL16可能調控雞成肌細胞的增殖、分化和肌纖維類型形成,在雞肌肉功能調控中發揮重要功能。
