蘇尼特羔羊褐色脂肪鑒定及特征研究
2023-02-27 14:26:54方勤圓付紹印何小龍何江峰王力偉王大清張立嶺劉永斌
畜牧獸醫學報 2023年2期
方勤圓,付紹印,王 標,何小龍,何江峰,王力偉,陳 欣,張 林,王大清,張立嶺,劉永斌*
(1.內蒙古大學生命科學學院,呼和浩特 010021; 2.內蒙古自治區農牧業科學院,呼和浩特 010031)
哺乳動物脂肪組織主要可以分為白色脂肪組織(white adipose tissue, WAT)和褐色脂肪組織(brown adipose tissue, BAT)[1]。其中,白色脂肪在哺乳動物體內分布最為廣泛,主要功能是儲存能量。不同于白色脂肪組織,褐色脂肪組織是幼小哺乳動物非顫栗性產熱的主要熱量來源,同時消耗機體攝取的過剩能量,維持能量平衡[2]。并且褐色脂肪細胞內具有豐富的線粒體與細胞色素,使得細胞呈現褐色[3],線粒體內膜上有解偶聯蛋白1(uncoupling protein 1, UCP1)。UCP1蛋白由305個氨基酸組成,形成6個由α-螺旋組成的跨膜結構域,分子量為32 ku。UCP1蛋白能夠通過解偶聯作用降低膜電位,形成質子滲漏由此介導細胞產熱。隨著人們生活水平的提高,由飲食引發的糖尿病越來越普遍,研究人員也越來越關注這種高發疾病的治療方法。其中褐色脂肪組織與糖尿病以及肥胖的治療密切相關,因此一直備受關注。鑒于褐色脂肪的特殊性,越來越多關于褐色脂肪的研究也開始轉向大型哺乳動物,尤其是家畜。研究人員期望通過提高家畜的褐色脂肪含量來減少白色脂肪沉積,提高瘦肉率,由此提高肉品風味和口感。
在世界各地眾多的綿羊品種中,蒙古羊是歷史悠久、分布廣泛、種群多樣、特征明顯、品種眾多、規模最大的亞洲種群之一。其主體分布于蒙古高原及其周邊高原,其中規模最大、最有特色、綜合性能最突出的種群分布于蒙古高原東部[4]。例如生活在內蒙古自治區的錫林郭勒半干旱草原上的蘇尼特羊。蘇尼特羊屬于蒙古系綿羊的一種,是肉脂兼用粗毛型地方綿羊品種[5],具有耐寒、抗旱、適應性強、肉鮮嫩多汁、無膻味、胴體豐滿,肉層厚實緊湊、高蛋白、低脂肪、瘦肉率高、板皮厚等優點[5]。蘇尼特羊的相關研究大多集中在肉品質與脂肪沉積等方面,但是很少與褐色脂肪相關。利用體外培養技術可以充分研究和利用褐色脂肪細胞,從胚胎或羔羊的BAT中分離出褐色脂肪前體細胞,在體外研究褐色脂肪的分化和功能以及與白色脂肪之間的關系。進而深入研究蘇尼特羊BAT在體內的生物學功能并進一步利用BAT選育和培育耐寒和瘦肉型的蘇尼特羊。但是對于蘇尼特羊褐色脂肪特點并沒有較為系統的研究,為彌補這一空白,本研究從細胞形態學水平、細胞超微結構、基因水平和蛋白水平系統探究蘇尼特羊褐色脂肪的分布特點,初步了解蘇尼特羊的BAT分布。對進一步研究褐色脂肪在家畜個體生長代謝方面以及優化育種方面具有重要意義。
1 材料與方法
1.1 材料
1.1.1 試驗動物 蘇尼特羊來源于巴彥淖爾市烏拉特中旗蘇尼特羊種羊場,相同飼養條件下(母乳喂養),選擇出生1、7和30 d的蘇尼特羊各3只,包括2只公羊1只母羊。不同日齡蘇尼特羊平均體重分別為3.83、6.75、11.17 kg。
1.1.2 主要試劑及儀器 無水乙醇、二甲苯、鹽酸、氨水、中性樹膠均購自國藥集團化學試劑有限公司;蘇木素-伊紅染液購自武漢谷歌生物科技;脂肪專用固定液、電鏡固定液、BSA均購自武漢賽維爾生物科技有限公司;RNAiso Plus、PrimeScriptTMRT Master Mix、TB Green Premix Ex TaqTM Ⅱ均購自TaKaRa公司。Anti-UCP1 Antibody購自Abcam;Anti β-Actin Mouse Monoclonal Antibody、Goat Anti-Mouse IgG, HRP Conjugated、Goat Anti-Rabbit IgG,HRP Conjugated均購自康為世紀。脂肪組織蛋白提取試劑盒、BCA法蛋白定量試劑盒均購自百奧萊博。
脫水機、包埋機、凍臺購自武漢俊杰電子有限公司、病理切片機購自上海徠卡儀器有限公司、組織攤片機購自浙江省金華市科迪儀器設備有限公司、NIKON ECLIPSE CI正置光學顯微鏡廠家為日本尼康、Leica UC7切片機購自Leica公司、TECNAI G2 20 TWIN透射電子顯微鏡廠家為FEI、熒光定量PCR儀、垂直電泳槽、半干轉印儀和高壓電泳儀購自美國BIO-RAD公司、全波長掃描式多功能讀數儀、低溫高速離心機購自美國THERMO FISHER公司。
1.2 方法
1.2.1 樣品采集與處理 采用放血法處死羔羊后,采集皮下脂肪,包括頸部、背部、尾部、胸部和腹股溝脂肪,以及內臟脂肪,包括腎周脂肪和心包脂肪。采集的脂肪一部分立即投入脂肪專用固定液中,一部分投入電鏡固定液中,用于切片的制作;一部分投入液氮中,后轉入-80 ℃冰箱保存,用于提取RNA及蛋白。
1.2.2 石蠟切片與HE染色 固定至少24 h后,將脂肪組織從固定液中取出,進行修塊。經過脫水浸蠟、包埋后,切成厚度為4 μm的薄片,再進行脫蠟、染色、脫水、封片等步驟。在光學顯微鏡下鏡檢,進行圖像采集。在200倍視野下,選取脂肪組織切片的5個視野,利用軟件Image-Pro Plus 6.0,對脂滴面積進行統計。
1.2.3 免疫組織化學 石蠟切片脫蠟后將切片放在盛滿檸檬酸抗原修復緩沖液的修復盒中,于微波爐內進行抗原修復,脫色搖床上晃動洗滌。切片放入3%雙氧水中,避光、室溫條件下孵育25 min,阻斷內源性過氧化物酶。3% BSA室溫封閉。在切片上滴加配好的一抗,4 ℃孵育過夜。洗滌后加二抗孵育。洗滌后稍甩干后滴加DAB顯色液,在顯微鏡下觀察控制顯色時間,顯色棕黃色為陽性位點,自來水沖洗切片終止顯色。蘇木素復染細胞核,脫水封片。
1.2.4 透射電鏡 將脂肪組織在4 ℃條件下固定2~4 h后取出,進行漂洗、固定、脫水、滲透、包埋等步驟,切成60~80 nm的超薄切片。在進行鈾鉛雙染色,在透射電子顯微鏡下觀察,采集圖像分析。
1.2.5 RNA 提取、反轉錄與實時熒光定量PCR 用TRIzol法提取各部位脂肪組織的總RNA,對RNA的濃度與純度進行檢測。利用TaKaRa公司的PrimeScriptTMRT Master Mix試劑盒進行反轉錄。
在GenBank數據庫中查找目的基因Ucp1和內參基因GAPDH的序列。利用Primer Premier 5.0設計引物(表1)。通過比對確保其特異性后交由生工生物工程(上海)股份有限公司合成。

表1 Ucp1和GAPDH基因引物序列
實時熒光定量PCR體系為:95 ℃ 3 min;95 ℃ 5 s;62 ℃ 20 s;72 ℃ 30 s;72 ℃ 7 min;40個循環,每個樣品進行3次技術重復。
1.2.6 蛋白提取以及Western blot 脂肪組織總蛋白提取:取200 mg脂肪組織樣本剪碎,加入500 μL蛋白提取液,震蕩使其充分勻漿。將組織勻漿轉移到一預冷的干凈離心管中,置于冰上1 h,每隔5~6 min用移液器吹打混勻。在4 ℃,12 000×g條件下離心15 min。將上清吸入另一預冷的干凈離心管,得到蛋白樣品。用BCA法測定蛋白濃度。進行SDS-PAGE電泳,半干轉進行轉膜。5%脫脂奶粉封閉,一抗1∶1 000稀釋后4 ℃孵育過夜,洗滌后二抗孵育,洗滌后ECL顯色。按照Western blot試驗條件重復3次。
1.2.7 數據統計與分析 根據2-ΔCt方法計算不同組織間基因相對表達水平,用Graphpad Prism6.0軟件進行作圖。對不同部位以及不同日齡脂肪組織的基因表達量進行獨立性t檢驗,以P<0.05表示差異顯著,以P<0.01表示差異極顯著。
2 結 果
2.1 蘇尼特羊褐色脂肪組織和白色脂肪組織特點
2.1.1 不同類型脂肪組織的HE染色 對蘇尼特羔羊的各部位脂肪進行HE染色,有兩種不同類型的脂肪細胞,褐色脂肪細胞和白色脂肪細胞。褐色脂肪細胞內為多室的小脂滴。白色脂肪細胞為空泡狀的單室大脂滴,細胞核被大脂滴擠壓到細胞邊緣(圖1A、1B)。BAT的脂滴面積小于WAT的脂滴面積,且差異顯著(P<0.05,圖1C)。

*. 差異顯著(P<0.05),下同
2.1.2 不同類型脂肪組織的透射電鏡 通過對不同類型脂肪組織進行透射電鏡觀察,褐色脂肪細胞內脂滴為多室的小脂滴,并且在脂滴之間分布著大量有嵴的線粒體,線粒體呈現圓形或橢圓形(圖2A、2B)。白色脂肪細胞內只有一個空泡狀的大脂滴,幾乎占滿了細胞的整個空間,細胞核被大脂滴擠壓到了細胞的邊緣,細胞內只有少量的線粒體(圖2C、2D)。
2.1.3 不同類型脂肪組織的免疫組化 對BAT和WAT切片進行免疫組化,觀察BAT特異性蛋白UCP1在細胞中的分布。BAT中有大量顯示棕黃色的陽性位點(圖3A),同時也能觀察到組織中含有小脂滴的細胞,在細胞中間或邊緣有藍紫色的細胞核。而WAT中沒有陽性位點(圖3B)。對免疫組化的陽性位點統計分析表明,BAT與WAT的陽性位點差異極顯著(P<0.01,圖3C)。

***. 差異極顯著(P<0.01),下同
2.2 不同日齡羊各部位脂肪形態特點
對不同日齡蘇尼特羊各部位脂肪進行HE染色。可以觀察到出生1 d時腎周、頸部、背部、腹股溝和心包脂肪都有褐色脂肪細胞的分布。出生7 d時腎周、頸部、腹股溝和心包脂肪有褐色脂肪細胞。出生30 d時,腎周、頸部、心包脂肪依舊有少量的褐色脂肪細胞分布在白色脂肪細胞之間。而尾部脂肪和胸部脂肪都是白色脂肪細胞(圖4)。

PrF. 腎周脂肪;CF. 頸部脂肪;DF. 背部脂肪;TF. 尾部脂肪;StF. 胸部脂肪;IngF. 腹股溝脂肪;PcF. 心包脂肪;下同
2.3 不同日齡羊各部位脂肪組織Ucp1的表達
通過實時熒光定量PCR檢測不同日齡蘇尼特羊各部位脂肪組織Ucp1基因的表達。出生1 d時,腎周脂肪的Ucp1基因表達量最高,頸部、背部、腹股溝和心包脂肪都有Ucp1基因表達。7 d時,腎周、頸部、心包脂肪基因表達量較高,與出生1 d時差異不顯著。30 d時,Ucp1基因表達量較低,下降幅度較大。尾部和胸部脂肪組織已檢測不到Ucp1基因的表達(圖5)。

圖5 不同日齡蘇尼特羊各部位脂肪組織的Ucp1基因表達
2.4 不同日齡羊各部位脂肪組織UCP1蛋白表達
通過Western blot檢測不同日齡蘇尼特羊各部位脂肪組織UCP1蛋白的表達。出生1 d時,腎周脂肪的UCP1蛋白表達量最高,頸部、背部、腹股溝和心包脂肪都有UCP1蛋白表達。7 d時,腎周、頸部、心包脂肪蛋白表達量較高,與出生1 d時差異不顯著。30 d時,除了腎周脂肪,其他部位UCP1蛋白表達量較低,下降幅度較大。尾部和胸部脂肪組織已檢測不到UCP1蛋白表達(圖6)。
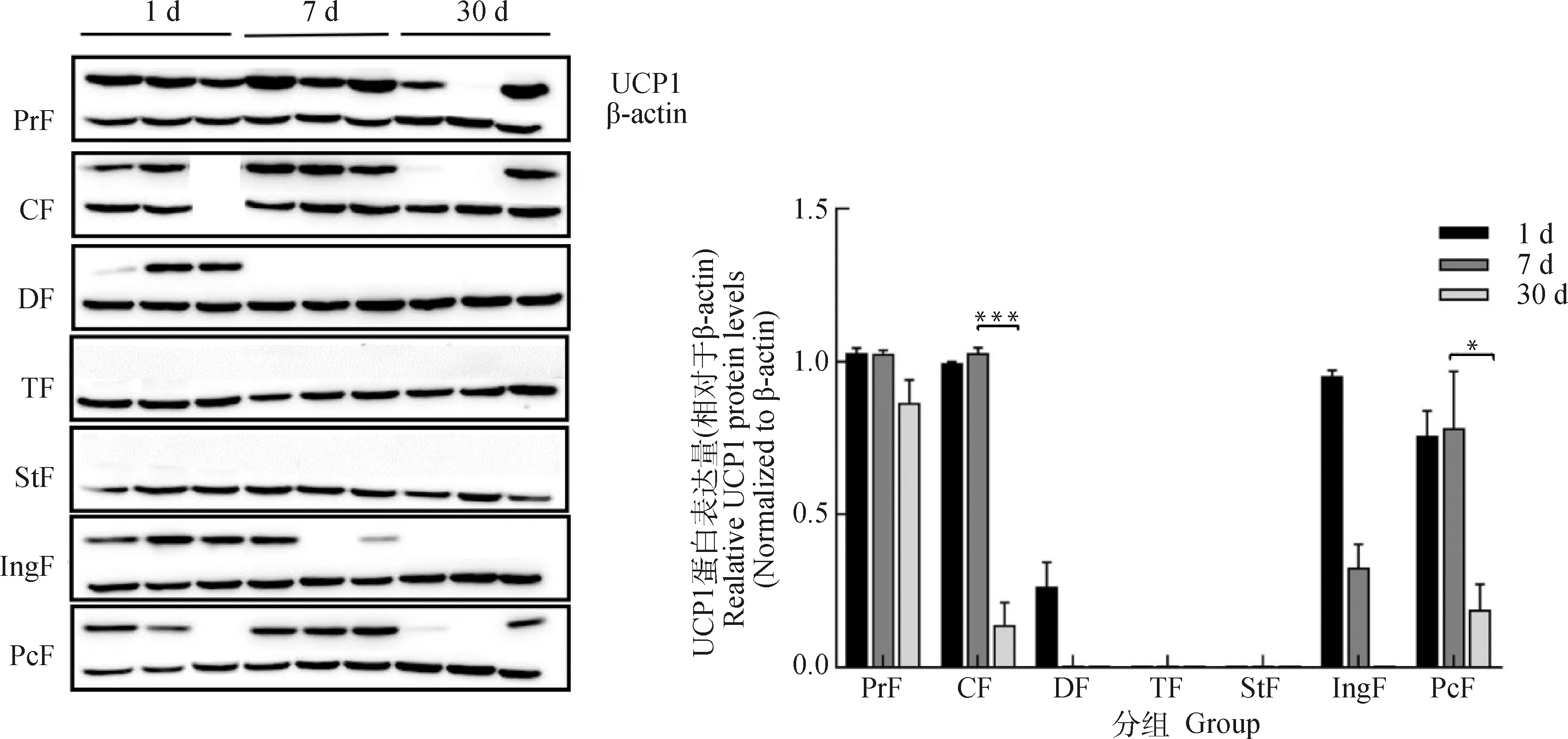
圖6 不同日齡蘇尼特羊各部位脂肪組織的UCP1蛋白表達
3 討 論
3.1 蘇尼特羔羊褐色脂肪的鑒定
事實上,BAT它既不能稱為脂肪,也不能稱為肉,介于兩者之間,類似于牛乳腺,這是最早對于褐色脂肪的形態描述。通過對蘇尼特羊脂肪的組織學觀察,本研究發現蘇尼特羊的褐色脂肪和白色脂肪有明顯的形態學差異。類似于小鼠褐色脂肪與白色脂肪的顏色[6],通過解剖蘇尼特羔羊,采集不同部位的脂肪,可以觀察到蘇尼特羊褐色脂肪顏色呈現類似肌肉的紅褐色,與白色脂肪相比顏色更深。蘇尼特羊脂肪經過HE染色之后,褐色脂肪由于細胞內脂滴為多室的小脂滴,而白色脂肪細胞內單室的大脂滴占滿細胞的整個空間。白色脂肪的脂滴面積顯著大于褐色脂肪的脂滴面積。通過脂肪細胞的形態學觀察和褐色脂肪特異性標記基因Ucp1的檢測,能夠鑒定出不同部位、日齡的脂肪類型。Basse等[7]對特克賽爾羊和哥特羊雜交的后代綿羊進行褐色脂肪的相關研究,不同類型脂肪組織的HE染色顯示出同樣的結果。Gemmell等[8]早在1972年就通過透射電鏡對綿羊褐色脂肪線粒體進行了系統地觀察,表明褐色脂肪具有較多的線粒體,并且線粒體的嵴較多且平行排列。隨著脂肪類型的變化,線粒體數量逐漸變少,線粒體嵴變得不規則。本試驗中,蘇尼特羊褐色脂肪的透射電鏡也顯示細胞內分布著大量的具有規則嵴的線粒體;而在白色脂肪細胞內,線粒體數量極少,且并不是所有線粒體嵴都規則排列。通過不同類型脂肪細胞的結構以及分子特點,確定了不同的脂肪組織類型。
3.2 蘇尼特羊褐色脂肪分布部位
蘇尼特羊7個部位中,腎周、頸部、背部、腹股溝和心包都有褐色脂肪細胞分布。褐色脂肪在人體內主要分布在腎周、頸部、肩胛部、腋下和脊柱旁等部位[9],在小鼠體內的分布與在人體內分布相似。2009年,Marken等[10]通過正電子發射斷層-X射線計算機斷層掃描(positron emission tomography, computer tomography,PET-CT)發現成人體內,主要為肩胛部有褐色脂肪的分布。關于嚙齒類動物褐色脂肪的研究,褐色脂肪來源部位一般為肩胛部[6,11]。綿羊褐色脂肪的研究也大多集中于腎周脂肪[12-13],在本試驗中也發現腎周脂肪中褐色脂肪細胞數量最多,UCP1表達量最高。而蘇尼特羊尾部和前胸部脂肪都是白色脂肪。綿羊尾部脂肪在胚胎時期就開始沉積,出生后脂尾已經基本形成,但是無論在哪個時期都沒有研究表明尾脂中有褐色脂肪細胞。本試驗得出相同的結論,說明尾部是白色脂肪的經典部位。Thompson等[14]的研究通過褐色脂肪的特異性標記蛋白UCP1的表達證明在綿羊體內背部脂肪有褐色脂肪細胞。也有研究表明綿羊心包脂肪中有UCP1的表達[15]。本研究也發現背部和心包脂肪有褐色脂肪細胞。2017年Henry等[16]研究了雌性綿羊胸骨脂肪的產熱作用,研究結果表明成年母羊胸部脂肪有褐色脂肪細胞存在,但是在本試驗中沒有檢測到胸部脂肪有褐色脂肪細胞。
3.3 不同日齡蘇尼特羊褐色脂肪變化
反芻動物常常作為研究褐色脂肪分化的動物模型,很早就有研究發現綿羊在出生時UCP1表達處于峰值,隨著個體生長,WAT不斷增加,UCP1表達逐漸下降[17-20]。在140 d的胎羊中可以檢測到Ucp1的表達,說明從妊娠后期開始,胎羊開始分化出褐色脂肪[20]。隨著時間的增長,UCP1表達逐漸下降,到1個月時已完全沒有褐色脂肪細胞[15]。有研究測定了出生1、7和30 d綿羊Ucp1基因的表達,結果顯示在出生1 d時其表達達到峰值,隨后下降,到30 d時沒有表達[18]。還有研究表明,Ucp1 mRNA在綿羊所有組織和2~12個月齡各階段均有表達,但在腎周脂肪中的表達水平明顯高于其他組織,皮下脂肪的表達水平低于深部脂肪[21]。本試驗表明,蘇尼特羊在出生1和7 d時都有褐色脂肪細胞,UCP1的表達量較高,到出生30 d時UCP1的表達降低。在出生30 d時腎周、頸部和心包等部位依舊有褐色脂肪細胞。推測蘇尼特羊褐色脂肪的這種分布特點可能是對環境產生的一種應激,以此來彌補本身發育的不完善。
褐色脂肪的分布本身就具有異質性,激活褐色脂肪活性的因素較多。包括環境以及營養條件等都會不同程度地影響綿羊褐色脂肪,尤其母羊妊娠后期的環境會極大地影響出生羔羊的褐色脂肪含量。例如高海拔缺氧環境[22]、寒冷環境[23]和脂肪酸攝入[24]等。因此在不同品種綿羊中褐色脂肪分布的差異性可能與外界環境有較大的關系。要研究綿羊的褐色脂肪特點以及這種差異產生的原因則需要設置更加密集的時間點、選擇更多品種的綿羊進行比較以及通過轉錄組測序等方法進行更深入的研究。
4 結 論
本研究通過形態學觀察和BAT標記基因、蛋白檢測區分并鑒定了蘇尼特羔羊的褐色脂肪和白色脂肪,證明了褐色脂肪的主要來源部位是腎周,而尾部是白色脂肪的主要來源部位。出生1和7 d時褐色脂肪細胞數量較多,出生30 d時,褐色脂肪細胞數量,標記基因、蛋白表達呈現下降趨勢。
